
Abstract
We previously identified multipotent stem cells within the lamina propria of the human olfactory mucosa, located in the nasal cavity. We also demonstrated that this cell type differentiates into neural cells and improves locomotor behavior after transplantation in a rat model of Parkinson's disease. Yet, next to nothing is known about their specific stemness characteristics. We therefore devised a study aiming to compare olfactory lamina propria stem cells from 4 individuals to bone marrow mesenchymal stem cells from 4 age- and gender-matched individuals. Using pangenomic microarrays and immunostaining with 34 cell surface marker antibodies, we show here that olfactory stem cells are closely related to bone marrow stem cells. However, olfactory stem cells also exhibit singular traits. By means of techniques such as proliferation assay, cDNA microarrays, RT-PCR, in vitro and in vivo differentiation, we report that when compared to bone marrow stem cells, olfactory stem cells display (1) a high proliferation rate; (2) a propensity to differentiate into osseous cells; and (3) a disinclination to give rise to chondrocytes and adipocytes. Since peripheral olfactory stem cells originate from a neural crest-derived tissue and, as shown here, exhibit an increased expression of neural cell-related genes, we propose to name them olfactory ectomesenchymal stem cells (OE-MSC). Further studies are now required to corroborate the therapeutic potential of OE-MSCs in animal models of bone and brain diseases.
Introduction
W
Using immunomarkers and flow cytometry, we observed that human olfactory lamina propria-derived stem cells had little in common with neural stem cells and hematopoietic stem cells (unpublished data). We also demonstrated that these cells can give rise to mesodermal cell types [30], a feature never reported with neural stem cells. We therefore reasoned that, as resident of a connective tissue originating from a mesenchyme, this cell type could be part of the family of mesenchymal stem cells (MSC). In order to (in)validate our hypothesis, we combined the expertise of several teams with a recognized expertise in MSCs [33 –37] and comprehensively compared human olfactory lamina propria stem cells to human bone marrow MSC at the cellular and molecular levels. For that purpose, we devised a wide-ranging protocol, based on tools such as immunostaining, flow cytometry, microarray expression profiling, RT-PCR, in vitro and in vivo differentiation studies.
Materials and Methods
Patients
Human nasal mucosa and human bone marrow cells were obtained as described before [13,34]. Nasal biopsies were collected from 4 individuals, 2 males and 2 females aged 23 to 52 years, at North University Hospital (Marseille, France); bone marrow cells were collected from 4 age- and gender-matched individuals, 2 males and 2 females aged 25 to 56 years, at Tours University Hospital (Tours, France). Samples were obtained under a protocol that was approved by the local ethical committee (Comité de Protection des Personnes) of Marseille and Tours, respectively. Informed consent was given by each individual participating in the study, in accordance with the Helsinki convention (1964) and French law relating to biomedical research. For extensive transcriptome analyses, 32 samples of stem/progenitors cells from 32 donors were assessed. Data on olfactory stem cells from 3 individuals were compared to a databank of bone marrow stem cells, CD11b+ cells, CD45+ cells, CD235a+ cells, synovial fibroblasts, periosteal cells, and neural progenitor cells from 9, 3, 3, 3, 4, 3, 4 individuals, respectively [35].
Cell culture of olfactory lamina propria stem cells
Human olfactory lamina propria stem cells were cultivated as previously described [30]. However, because human olfactory laminae propriae are highly compacted tissues, their dissociation remains incomplete, whatever the cocktail of enzymes used. To overcome this hurdle, we devised a new culture technique. After collection in the operating theatre, olfactory mucosae were immediately placed on ice in DMEM/HAM'S F12 (Invitrogen, Carlsbad, CA) supplemented with fetal bovine serum (FBS, 10%), penicillin (1%), and streptomycin (1%) and then incubated for 45 min at 37°C in a 2.4 U/mL Dispase II solution (Boehringer). Laminae propriae were carefully separated from the epithelium under a dissection microscope with a microspatula. Sheets of olfactory epithelium were discarded while laminae propriae were cut into pieces using microscissors. Each piece was then placed under a sterile glass coverslip in a well of 4-well plates (Nunc) and fed with 500 μL of serum-containing DMEM/HAM'S F12. Two to three weeks after the initial plating (when confluency was reached), the coverslip was removed, inserted upside down in a well of a 6-well plate, and cells were fed with the same serum-containing medium. When confluent, adherent cells, on both the glass slide and the plastic culture dish, were passaged and replated at a density of 1,000 cells/cm2. In order to generate spheres, cells were replated at a density of 16,000 cells/cm2 into flasks pretreated with poly-
Cell culture of bone marrow mesenchymal stem cells
Human bone marrow mesenchymal stem cell cultures were established from bone marrow aspirates, as previously described [34,38]. In brief, bone marrow nuclear cells were plated at a density of 5 × 104 cells/cm2 in α-minimum essential medium (α-MEM), supplemented with 10% FBS, 1 ng/mL fibroblast growth factor 2 (FGF2), 100 U/mL penicillin, and 100 μg/mL streptomycin. When cultures reached near confluence, cells were detached with 0.05% trypsin and 0.53 mM ethylene diamine tetracetic acid (EDTA) and subsequently replated at a density of 1,000 cells/cm2. For generating spheres, cells were replated at the density of 16,000 cells/cm2 into flasks pretreated with poly-
In vitro proliferation assay
The assay was performed by a single operator using the same protocol for the 2 cell populations. Passage 1 (= 2 weeks of culture) and 10 (>15 weeks of culture) heterogeneous olfactory (n = 3) and bone marrow (n = 3) stem cells were seeded at the density of 200 cells/cm2 in 24-well plates and grown without passage with a medium change [DMEM/HAM'S F12 (Invitrogen) supplemented with FBS (10%), penicillin (1%), and streptomycin (1%)] every 3 days. After being paraformaldehyde-fixed and stained with Hoechst blue, cells grown in the 11 tested conditions (9, 31, 55, 81, 102, 127, 151, 175, 223, 295, 360 h) were counted in duplicate, each day, using an inverse microscope.
Clonal efficiency assay
The assay was carried out by plating early passage (P3) cells from both populations, olfactory (n = 3) and bone marrow (n = 3) stem cells, in 6-well plates at densities ranging from 320 to 10 cells/well in duplicate by using a 1:2 serial dilution. After plating, the dishes were placed in an incubator (37°C, 5% CO2) and left there for 21 days before being paraformaldehyde-fixed (4%). Crystal violet-stained colonies were counted. For each sample, clonal efficiency was calculated as follows: (mean number of colonies/total number of seeded cells) × 100. When too many colonies were overlapping, no counting was performed.
Flow cytometry analysis
Cells were trypsinized, centrifuged (250g, 5 min) and resuspended in cold PBS. One hundred thousand (105) cells were incubated 30 min at 4°C in cold PBS (200 μL) with phycoerythrin (PE)-labeled monoclonal antibodies, added at saturating concentration (see the Supplementary Table 1 (supplementary materials are available online at
Reverse transcription-polymerase chain reaction (RT-PCR) analysis
RT-PCRs were performed using SuperScript One-Step RT-PCR with Platinum Taq DNA polymerase (Invitrogen) according to the manufacturer's instructions. The sequence of PCR primers were for: housekeeping gene glyceraldehyde 3-phosphate dehydrogenase (GAPDH) (forward) 5′-GGTGAAGGTCGGTGTGAACG-3′, (reverse) 5′-CATC ATACTTGGCAGGTTTCTCC-3′; aggrecan (AGC) (forward) 5′-ACCCGCTACTCGCTGACCTTT-3′, (reverse) 5′-AAGGCGTGTGGCGAAGAACA-3′; PTHR1 (forward) 5′-ACATCTGCGTCCACATCAGGG-3′, (reverse) 5′-CCGTTCACGAGTCTCATTGGTG-3′; COL2A1 (forward) 5′-AACTGGCAAGCAAGGAGACA-3′, (reverse) 5′-AGTTTCAGGTCTCTGCAGGT-3′. All total cellular RNA samples were extracted with the RNA easy kit (Qiagen SA., Courtaboeuf, France) from cells cultured in proliferating medium or in differentiating medium. For specific mRNAs, 50 ng of total RNA were used and 5 ng for the GAPDH mRNA. RT-PCR products were analyzed with agarose gel electrophoresis.
Microarray experiments
Total RNA from stem cells was isolated as described above. Three samples of human olfactory lamina propria stem cells were compared to 9 bone marrow stem cell samples prepared by the same operator under the same conditions: identical culture medium, similar level of confluency (80%–90%), equal passaging (P3). In addition, in order to avoid any circadian rhythm-related variations in gene expression, cells were collected at the same time, the same day. Hybridization on HG-U133 Plus 2.0 microarrays was performed according to the standards supplied by the manufacturer (Affymetrix, Santa Clara, CA, USA). In brief, cDNA was synthesized from 3 μg of total RNA and submitted to in vitro transcription (ENZO Biochem, New York, NY) to generate biotin-labeled complementary RNA. Fragmented complementary RNA (15 μg/300 μL) was hybridized to the GeneChips for 16 h at 45°C. Arrays were washed and stained under standardized conditions and scanned on a Hewlett Packard Genearray Scanner (Affymetrix). Affymetrix GCOS 1.2 software was used to control washing and scanning, to generate DAT, CEL, and EXP files, and to process the raw data for signal calculation and pairwise chip comparison. Group comparison and gene retrieval were performed using the BioRetis database (
In vitro AOC differentiation assays
For adipogenic differentiation, olfactory and bone marrow stem cells were plated and grown for 2 days at 37°C, 5% CO2 in DMEM containing 10% FBS, 0.5 mM 3-isobutyl-1-methyl-xanthine (Sigma), 1 μM dexamethasone (Sigma), and 60 μM indomethacin (Sigma). Medium was then replaced by DMEM + 10% FBS, with 10 μg/mL insulin and cells were grown for 6 additional days with medium changes every 2 days. Cells were then collected for quantitative RT-PCR or fixed in 4% paraformaldehyde (Sigma) prior to staining with Nile Red O (0.3% w/v Oil Red O (Sigma)), in cold PBS (1/1,000) during 30 min. For osteogenic differentiation, olfactory and bone marrow stem cells were grown in DMEM + 10% FBS, 0.1 μM dexamethasone, 0.15 mM
In vitro neuronal differentiation assay
For neuronal differentiation, clone-derived olfactory stem cells (obtained using the limiting dilution method) were plated and grown, for 21 days at 37°C, 5% CO2 in neurobasal medium containing B27 (1×), 20 U/mL penicillin and streptomycin, 2 mM glutamine (Invitrogen), 0.025 mM glutamate (Sigma) with medium changes every 3 days. Cells were then fixed in 4% paraformaldehyde (Sigma) prior to immunostaining with anti-MAP2 antibody.
In vivo osteoblastic differentiation
Macro- and microporous biphasic calcium phosphate ceramic discs (diameter: 8 mm; thickness: 3 mm (MBCP, Biomatlante, Vigneux de Bretagne, France)) were used as scaffolds for olfactory stem cells. Ceramic discs were composed of hydroxyapatite (HA) and β-tricalcium phosphate (β-TCP) in a 20%/80% ratio, as evidenced by X-ray diffraction analysis. Double packaged and gamma sterilized discs were first incubated for 48 h in 5 mL of DMEM/HAM'S F12 supplemented with 10% FBS, 100 U/mL penicillin, and 100 μg/mL streptomycin in 6-well plates. After discarding excess medium, twice passaged olfactory stem cells were then seeded on ceramics at a density of 105 cells/disc. After cell attachment on ceramic for 3 h, osteogenic differentiation media (3 mL) composed of DMEM high glucose supplemented with 10% serum, 1% penicillin/streptomycin, dexamethasone (0.1 μM),
Immunocytochemistry and quantification
Immunocytochemistry was carried out to identify cells expressing nestin marker or new born cells (proliferation assay). For BrdU immunostaining, the DNA was denatured using a pretreatment with HCl [2 N] for 30 min followed by an incubation in NaOH solution [0.02 N] for 10 s and in sodium borate solution [0.1 M] for 2 min. After 3 washes in PBS, cells were blocked in 5% goat serum, 3% bovine serum albumin (BSA), 0.1% Triton X-100 in PBS for 90 min at room temperature. We used the following primary antibodies: rabbit antinestin (1/500) (Abcys), rabbit anti-MAP2 (1/500) (Abcam, Cambridge, MA), mouse anti-NG2 (1/100; R&D systems, Minneapolis, MN). Cultures were incubated with primary antibodies, diluted in the blocking solution for 1 h at room temperature, washed 3 times, and incubated with the appropriate conjugated secondary antibody (goat anti-rabbit IgG conjugated with AlexaFluor 594 (1/500), goat anti-mouse IgG conjugated with AlexaFluor 488 (1/500)) for 1 h at room temperature. After several washes in PBS, slides were incubated with 0.5 μg/mL DNA intercalant Hoechst #33258 (Invitrogen, Cergy Pontoise, France) for 30 min and mounted with fluorescent anti-fading medium (ProLong® Gold; Invitrogen). For each coverslip, the total number of cells positive for BrdU was counted. Control conditions were carried out by omitting the primary antibody. The percentage of positive cells for each marker was represented as a mean percentage ± SEM.
Statistical analysis
Data were analyzed using SPSS/PC + statistics 11.0 software (SPSS Inc., Chicago, IL). Unpaired Student's t-test and Mann–Whitney test were used to detect any significant difference between olfactory and bone marrow stem cells.
Results
Olfactory stem cells are highly proliferative and give rise to spheres
Olfactory and bone marrow stem cells populations are composed of adherent cells with heterogeneous morphologies (Fig. 1A and 1B). Two key features of stemness—sphere formation and nestin expression—were evaluated. When grown on poly-
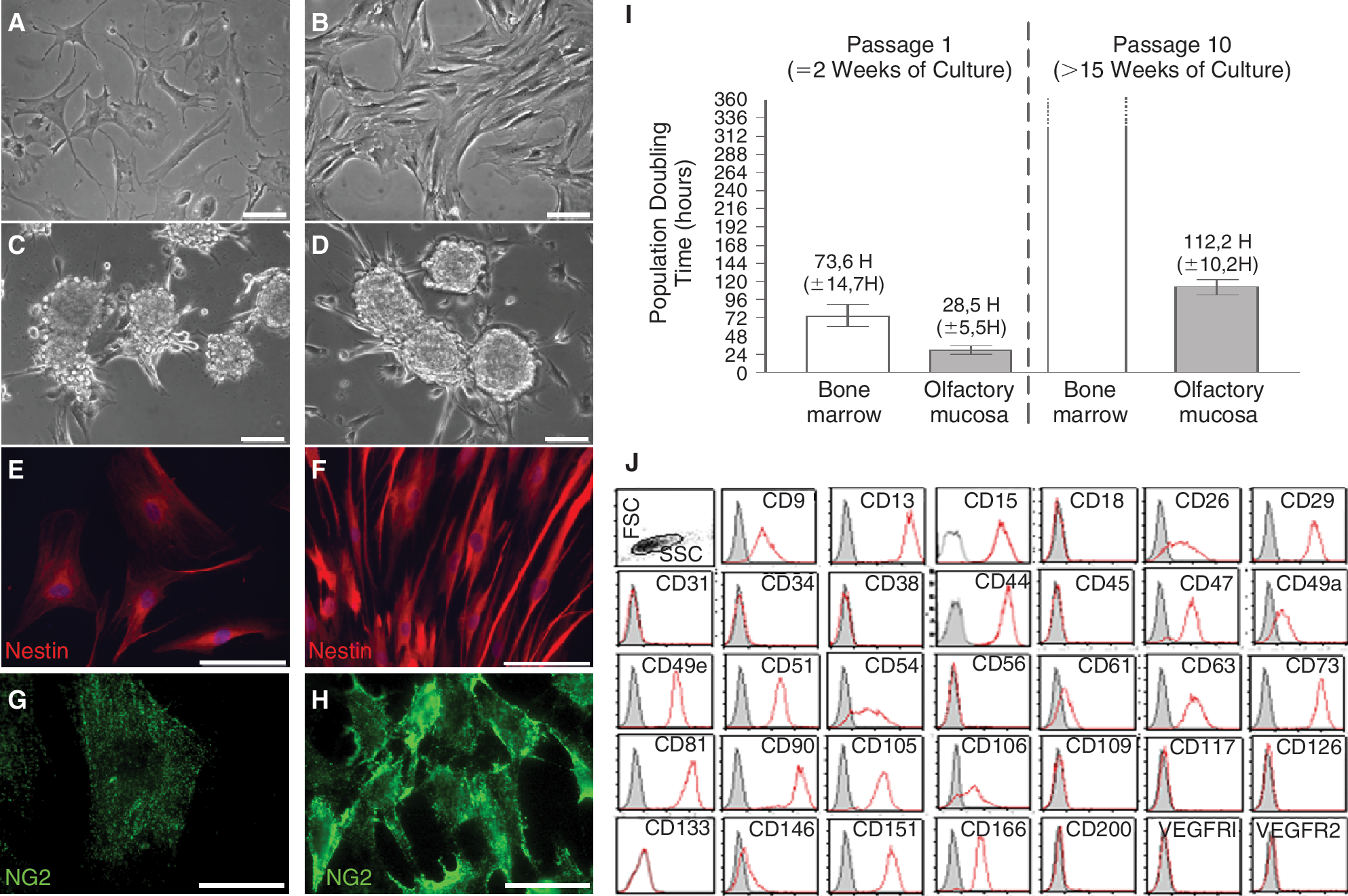
In vitro assessment of stemness characteristics and cell proliferation. Bone marrow stem cells (
Assessment of population doubling time in short- (2 weeks) and long-term (>15 weeks) culture revealed that olfactory stem cells are quickly dividing cells. As shown in Fig. 1I, doubling time at week 2 was 28.5 h (±5.5 h) and 73.6 h (±14.7 h) for olfactory and bone marrow stem cells, respectively. Long-term doubling time was 112.2 h (±10.2 h) for olfactory stem cells and remained unmet after 360 h for bone marrow stem cells. Quantifications were performed in duplicate on olfactory (n = 3) and bone marrow stem cells (n = 3).
Noticeably, our new method of culture (without enzymatic dissociation of the lamina propria) was efficient in providing numerous stem cells. When compared to olfactory stem cells obtained after enzymatic dissociation, no difference in cell proliferation, sphere formation, and nestin expression was observed.
Olfactory stem cells share membrane markers with bone marrow stem cells
As shown on Fig. 1H, we analyzed by flow cytometry a panel of 34 membrane markers. Out of 21 markers known to be expressed by bone marrow stem cells, 21 are also expressed by olfactory stem cells (Table 1).However, the protein CD200, a recently identified marker of bone marrow stem cells, is not found on olfactory stem cells. In addition, 2 other membrane proteins are differentially regulated in the 2 populations: CD9 (TSPAN29) is overexpressed while CD146 (MCAM) is clearly underexpressed in olfactory stem cells, when compared to bone marrow stem cells. Remarkably, among the 12 CD markers not expressed by bone marrow stem cells (including hematological and/or endothelial such as CD31, CD34, VEGFR2, or CD45), none was detected on olfactory stem cells. The CD analysis was performed in parallel on cells that, before being trypsinized, were either adherent or clustered in spheres. No difference in CD expression was observed.
Mean expression of membrane markers on undifferentiated olfactory stem cells (n = 4) in comparison to undifferentiated bone marrow stem cells (n = 4). The number, the symbol, and the relative fluorescence mean intensity (rMFI) for each protein are indicated. Overexpressed proteins on olfactory stem cells are highlighted in gray; underexpressed proteins on olfactory stem cells are shown in boldface. * indicates that P < 0.05, according to the Mann–Whitney test.
Gene clustering indicates that olfactory stem cells are closely related to bone marrow stem cells
Transcript expression of undifferentiated olfactory stem cells was compared to the transcriptome of undifferentiated bone marrow stem cells, using Affymetrix pangenomic microarrays. In order to check whether the similarity observed at the protein level was confirmed at the transcript level, we performed a hierarchical clustering and principal component analysis (PCA) of our gene array data. For that purpose, we used the panel of 147 CDs that we previously described [35]. Figure 2A and 2B shows that the olfactory stem cell population is homogeneous and, within the mesenchymal lineage, closely related to bone marrow stem cells. Both populations were clearly discriminated from neural progenitor cells (NPC) and from 2 hematopoietic cell populations (CD11b+/CD45+ and GlyA+ (CD235a) cells). Among mesenchymal cells, olfactory and bone marrow stem cells were clustered together and can be clearly distinguished from POCs and SFbs.
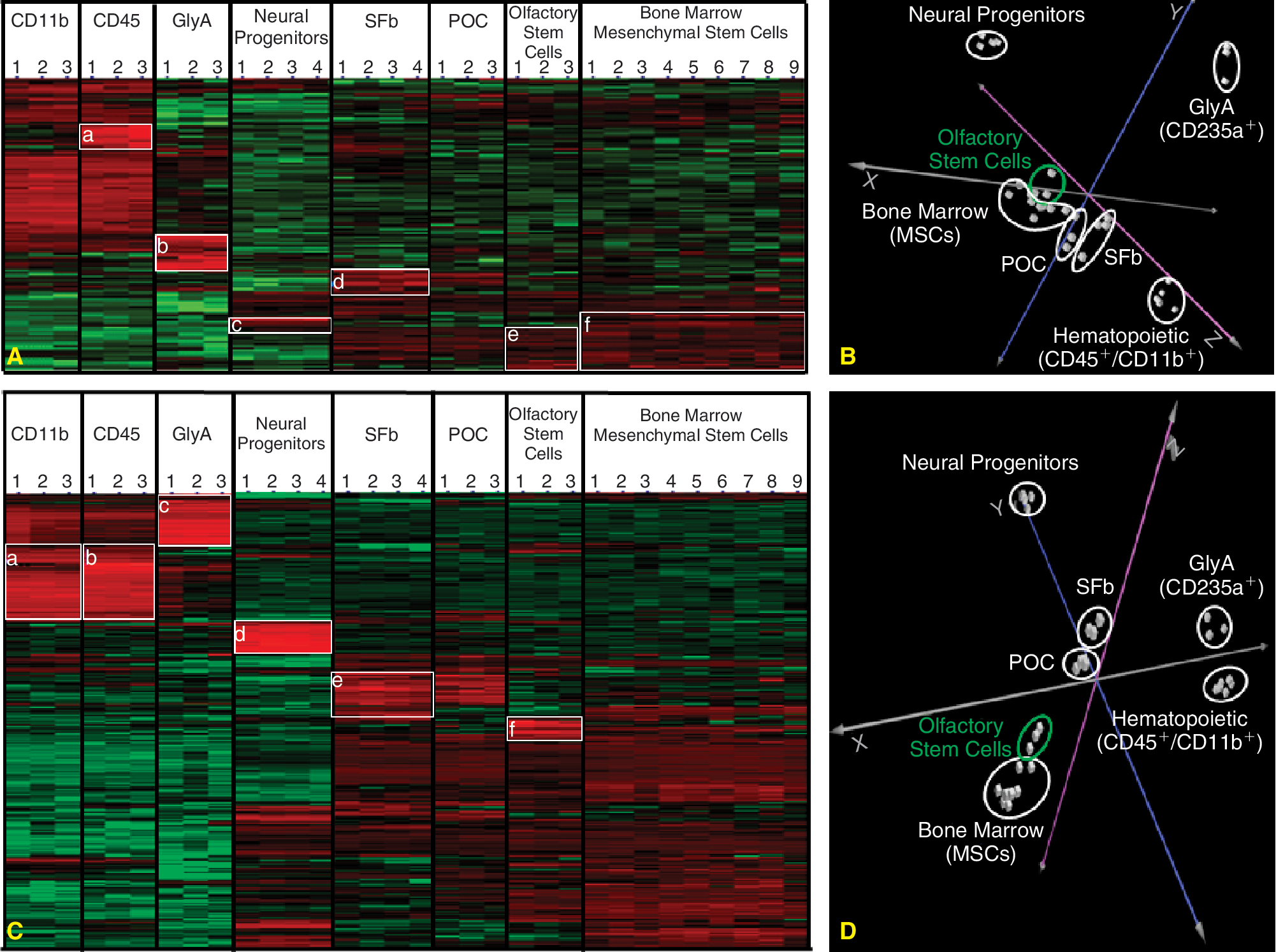
Transcriptome profile of olfactory stem cells. (
In order to refine our transcriptome profile, we performed an additional hierarchical clustering and principal component analysis using the top 50 up- and top 50 down-regulated transcripts in each cell type when compared to bone marrow stem cells. Figure 2 (C and D) shows that bone marrow and mesenchymal stem cells were discriminated from (1) hematopoietic stem cells according to the X-axis, (2) neural progenitor cells along the Y-axis, and (3) synovial fibroblasts and periosteal cells, according to the Z-axis. Supplementary Table 2 lists the top 50 up- and top 50 down-regulated transcripts in olfactory stem cells compared to mesenchymal stem cells.
The transcriptomic profile of olfactory stem cells reveals a propensity to differentiate into osseous cells
Although related to mesenchymal stem cells, olfactory stem cells display singular characteristics. In total, 417 transcripts with an absolute fold change above 3 were found differentially regulated (248 overexpressed and 169 under-expressed). Selected results are summarized in Table 2. Confirmatory results at the transcript level are observed for CDS, CD200, and CD146 (MCAM). The predisposition of olfactory stem cells to divide actively is also substantiated by the up-regulation of many genes involved in mitosis.
Selected dysregulated transcripts in undifferentiated olfactory stem cells in comparison to undifferentiated bone marrow stem cells. Genes are clustered by main biological functions and ranked by decreasing fold change. The symbol, the full name, and the fold change of each gene are indicated. Duplicate or triplicate transcripts are reported only once with the appropriate mean value. Underexpressed transcripts are highlighted in the gray overlays. Alternative names are mentioned in parentheses.
Bone marrow MSC are known to be able to differentiate into adipocytes, chondrocytes, and osteocytes. The list of differentially regulated transcripts in the olfactory stem cell group (Table 2)indicates that inhibitors of chondrogenesis (eg, asporin) and adipogenesis (eg, GATA2) are up-regulated while activators of neurogenesis (eg, neurogenesin-1) and osteogenesis (eg, OSR2) were overexpressed. Therefore, we predicted that these stem cells will exhibit (1) a poor ability to produce chondrocytes and adipocytes and (2) an enhanced aptitude to give rise to neural cells and osteoblasts. This prediction is supported by an RT-PCR experiment showing that olfactory stem cells express PTHR1 when they are undifferentiated while AGC and COL2A1 are undetectable in these cells before differentiation into chondrocytes. In addition, COL2A1 is not found after attempted chondrogenesis induction (Fig. 3O and 3P).
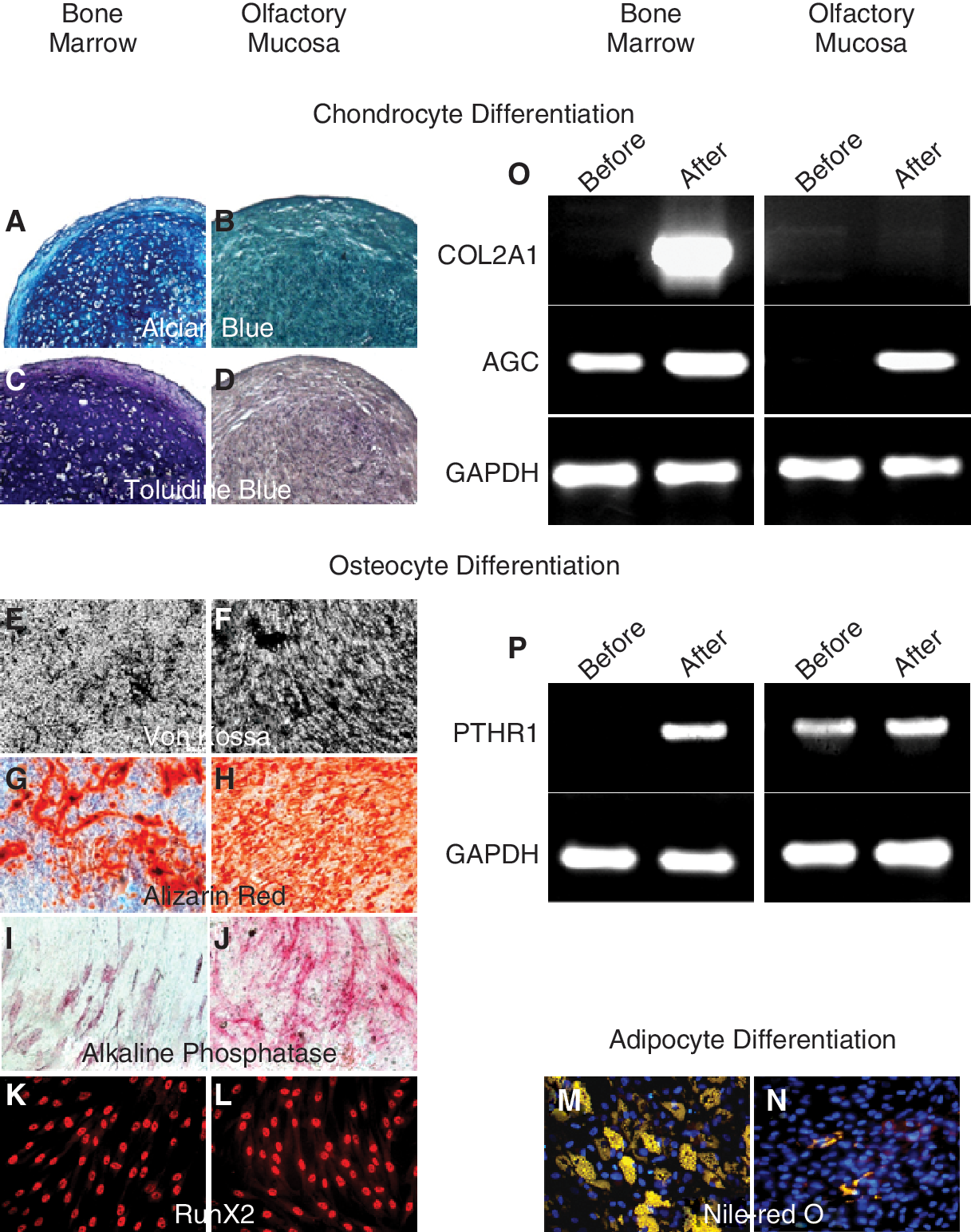
In vitro assessment of chondrogenesis/osteogenesis/adipogenesis and transcript expression of genes involved in chondrogenesis/osteogenesis. Bone marrow and olfactory mucosa stem cells were cultivated in a differentiating medium, for 6 to 21 days. (
Furthermore, Table 2 indicates an extensive overexpression of neuron- and glia-specific genes in the olfactory stem cell population.
Olfactory stem cells differentiate in vitro into osteoblasts and adipocytes but not chondrocytes
Figure 3 shows a striking difference between olfactory and bone marrow stem cells, in regard to adipogenic, osteogenic, and chondrogenic differentiation potential. Olfactory cells failed to produce chondrocytes (Fig. 3A–3D) and gave rise to a restricted number of adipocytes (Fig. 3M and 3N). In contrast, olfactory cells exhibited a robust inclination to differentiate into osteoblasts (Fig. 3E–3L).
Olfactory stem cells display a high clonogenicity and clone-derived cells give rise to bone cells and neurons
As shown in Fig. 4 (A and B), we compared the clonal efficiency of olfactory and bone marrow stem cells. A significant higher percentage (t-test, P < 0.0001) of olfactory stem cells (63.7% ± 3.1% (mean ± SEM)) gave rise to colonies, when compared to bone marrow stem cells (19.5% ± 1.9), after 21 days in culture. Quantifications were performed in duplicate on cultures of olfactory (n = 3) and bone marrow stem cells (n = 3). To assess whether the progeny of a unique cell may differentiate into both neurons and bone cells, we performed an in vitro differentiation assay using clone-derived olfactory stem cells. Figure 4 shows the capacity for a colony to give rise to either MAP2-positive neurons (C) or bone cells (D).
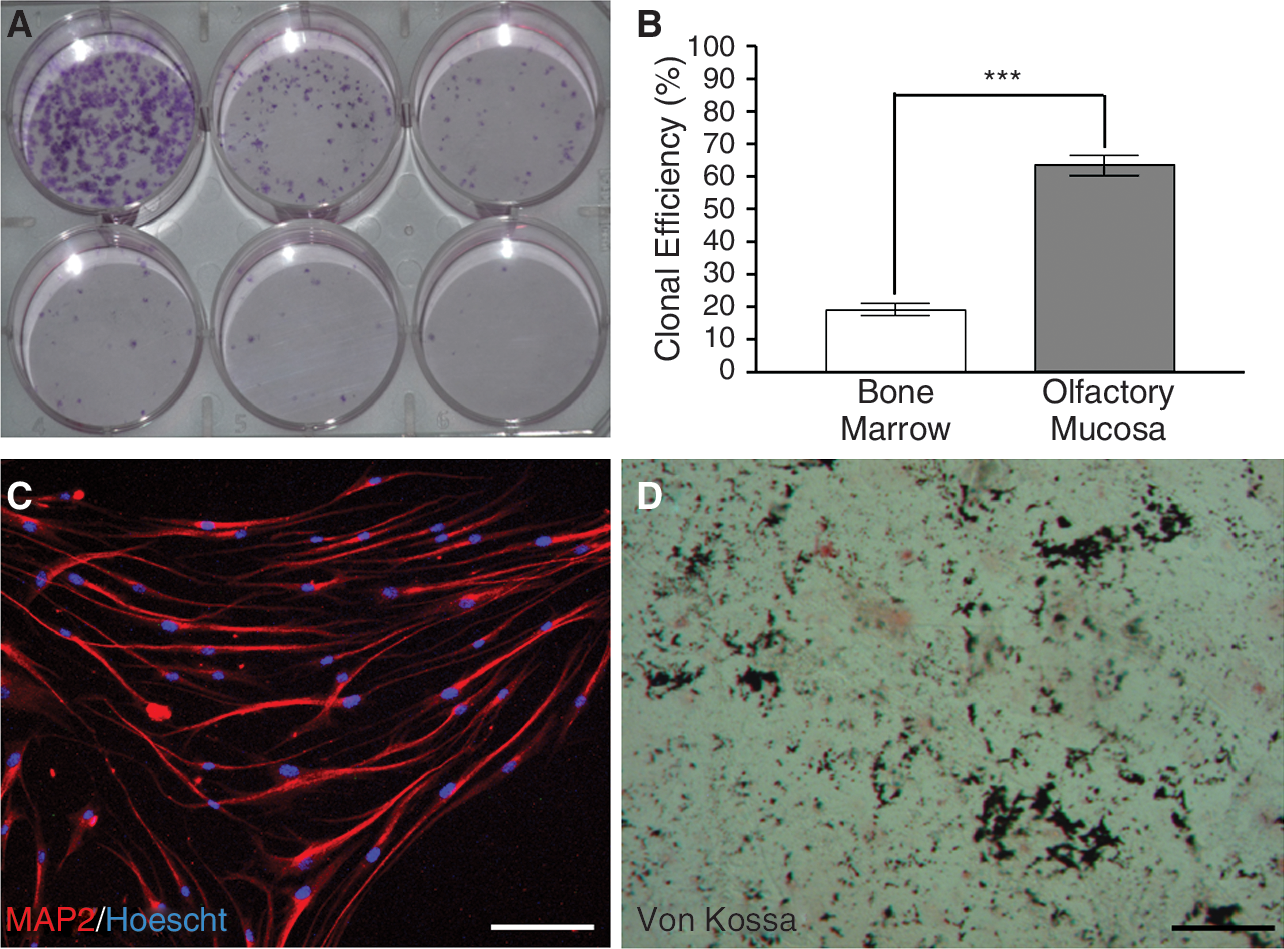
Olfactory stem cells display a high clonogenicity and clone-derived cells give rise to bone cells and neurons. Clonogenicity assay was performed by plating cells at densities ranging from 320 to 10 cells/well. At Day 21 post-plating, colonies were stained with crystal violet (
Olfactoy stem cells differentiate in vivo into osteoblasts
As shown on Fig. 5, the porous MBCP ceramics (Fig. 5A) harbors GFP-positive olfactory stem cells (Fig. 5B and 5C). Mineralized bone tissue with osteocyte lacunae was detected in the outer pores of the ceramic loaded with olfactory cells, 7 weeks after subcutis implantation of MBCP scaffolds (Fig. 5D and 5F). Osteoid tissue and adipocyte-rich bone marrow were also observed. The pores were filled for the most part by fibrous vascularized tissue. Green olfactory cells were co-localized with newly formed bone tissue (Fig. 5D–5G) that was found inside mature bone or at the margin of osteoid tissue. Exogenous olfactory cells were also observed in adipose and fibrovascular tissue. All constructs (10/10) exhibited ectopic bone formation while none of the ceramic discs implanted without cells did.

In vivo ectopic bone formation by olfactory stem cells in MBCP ceramic constructs. (
Discussion
The olfactory mucosa is a bi-stratum organ, including a neuroepithelium, facing the lumen, and a lamina propria, lining the nasal cartilage. Both tissues harbor stem cells. Neuroepithelial stem cells have been extensively studied by one team since 2001 [42]. Lamina propria stem cells were less investigated until recently. In a seminal paper, we showed that rodent and human lamina propria-derived stem cells can be purified, amplified, and differentiated into neural and non-neural cell types in vitro and in vivo [30]. We also demonstrated that human lamina propria-derived stem cells generate dopaminergic cells after transplantation in a rat model of Parkinson's disease [31]. Still, the characteristics of this new stem cell type remained largely unknown.
In order to depict the stemness profile of olfactory lamina propria-derived stem cells, we devised a comparative study. As we knew that olfactory stem cells had little in common with hematopoietic stem cells and, surprisingly, with neural stem cells (unpublished data), we searched for similarities/dissimilarities with another type of adult stem cells: the bone marrow MSC. The current study shows for the first time that olfactory stem cells can be considered as a mesenchymal cell subtype with specific traits, associated to their location in an ectodermal tissue. When compared to bone marrow MSC, olfactory lamina propria-derived stem cells display a high mitotic activity and a penchant for differentiating into cells with osseous phenotype. Conversely, these cells rarely give rise to chondrocytes and, to a lesser extent, to adipocytes. Moreover, we observed a higher clonal efficiency rate with olfactory stem cells. These data suggest that, when compared to bone marrow-derived cells, olfactory lamina propria-derived cell cultures include an increased percentage of stem cells.
A combinatorial approach with convergent data
In this study, we used a combination of cellular and molecular techniques. We first assessed the expression of 34 cell surface proteins, known for portraying bone marrow stem cells [35], using immunostaining and flow cytometry. Out of 34 markers studied, only 3—CD9, CD146, CD200—were found differentially regulated in olfactory stem cells when compared to mesenchymal stem cells. Interestingly, the pangenomic study revealed that the expression of CD9, CD146, and CD200 was also altered at the transcript level. Therefore, we can confidently state that olfactory stem cells differ from bone marrow stem cells by overexpressing CD9 and underexpressing CD146 and CD200. CD9 belongs to the tetraspanin family and is considered as a pluripotency marker [43] since it is involved in cell motility, metastasis, osteoclastogenesis, neurite outgrowth, oligodendrogenesis, myotube formation, angiogenesis and cell proliferation [44 –47]. CD200 is a membrane glycoprotein that co-stimulates T-cell proliferation and may regulate myeloid cell activity. It has been recently shown that CD200 is an immunosuppressive factor that allows the cancer stem cells to evade the immune system [48]. This very first transcriptomic analysis of olfactory lamina propria-derived stem cells was further refined by clustering the microarray data. The overexpression (with fold change up to +33) of 16 transcripts involved in cell division or apoptosis inhibition and the underexpression of one transcript playing a role as a proliferation inhibitor suggested that olfactory stem cells should have a high mitotic activity.
In regard to the canonical differentiation of MSC into adipocytes–chondrocytes–osteocytes, it was observed that olfactory cells overexpress transcripts coding for proteins known as (1) repressor (gata2) [49] or activator (lipin1) [50] of adipogenesis; (2) repressor (asporin) of chondrogenesis, and (3) activators (osr2, cyp27b1, pleiotrophin) [51 –53] of osteogenesis. Conversely, olfactory stem cells underexpress transcripts coding for proteins known as activators (chi3l1, xylt1, sox9) [54,55] of chondrogenesis.
It was therefore predicted that olfactory stem cells would exhibit (1) a high proliferation rate; (2) a predisposition for differentiating into osseous and neuronal cells, and (3) a reduced propensity to give rise to chondrocytes. Our conjecture was validated beyond expectation upon assessment of the differentiation potential of olfactory cells. When compared to mesenchymal stem cells, olfactory stem cells displayed (1) a high turnover rate in short- and long-term cultures, (2) a reduced ability to differentiate into adipocytes, (3) a poor chondrogenesis potential contrasting with (4) a potential for osteogenesis.
A new member of the mesenchymal stem cell family
MSCs are multipotent stem cells that can differentiate in vitro or in vivo into a variety of cell types, including adipocytes (A), osteoblasts (O), and chondrocytes (C) [56]. MSCs have been detected in a number of adult (adipose tissues, trabecular bone, periosteum, synovial membrane, tendon, skeletal, muscle tissue, teeth, brain), and fetal tissues (placenta, amnion) (for a review, ref. 57). This open list of tissues that produce MSCs can now be updated with the inclusion of the olfactory lamina propria-derived stem cells that we propose to coin olfactory ectomesenchymal stem cells (OE-MSC). The olfactory lamina propria is a relatively thick layer of connective tissue that originates from the fronto-nasal mesenchyme [58,59], itself arising from the interaction of cranial neural crest and olfactory placodes [60]. It is therefore unsurprising to identify within this tissue MSCs with ectodermal characteristics. This finding is in line with previous reports describing neural crest-derived MSCs, found in the branchial arch during development and in the teeth during adulthood, and named ectomes-enchymal stem cells [61]. Interestingly, it has been shown that OE-MSC (also named “mesenchymal–neural” precursors) have the capacity to differentiate into ectoderm and mesoderm cell types [62]. As a new evidence, we demonstrate here the pluripotency of OE-MSCs. One colony can give rise to either neurons or bone cells. Additionally, a recent study demonstrated that the earliest wave of MSCs in the embryo originates from a neuroepithelium, during a neural crest stage, and not, as previously thought, from mesoderm [63]. Later on, they are replaced by MSCs from unknown sources.
When compared to bone marrow MSCs, OE-MSCs display singular properties. First, its AOC potential is atypical. Contrary to bone marrow stem cells, OE-MSCs fail to differentiate in vitro into chondrocytes. This observation was unforeseen since, during development, the nasal mesenchyme condenses to form cartilage and bone [64]. Moreover, as the lamina propria lies on the cartilaginous plates (septum and turbinates) of the nose, one would have assumed that OE-MSCs could participate in repair mechanisms of damaged cartilage. However, it should be mentioned that a recent study reported that lamina propria-derived stem cells can be induced to express chondrogenic phenotype in a rat inter-vertebral disc injury model [65]. With respect to adipocytes, the picture is slightly different. OE-MSCs exhibit in vitro a low potential for adipogenesis in vitro but, when seeded in ceramics and implanted under a mouse skin, they can give rise to adipocyte-rich bone marrow. However, OE-MSCs appear to be more committed to osteoblast differentiation: when cultured on ceramics in vivo, OE-MSCs produce in vivo a mature mineralized bone tissue and our in vitro comparison with bone marrow stem cells clearly demonstrates a proclivity toward bone formation. This finding has never been documented before and further investigations focused on the up-regulated genes coding for proteins involved in osteogenesis (OSR2, CYP27B1, PTN, AEBP1) and calcium homeostasis (S100A4, CLGN, CAMK4, PRSS3, STC1, CHP) are now required.
Second, the proliferating rate of OE-MSCs is unusually high. We found a nearly 3-fold increase in population doubling time when compared to bone marrow stem cells, at week 2 post-plating. Moreover, bone marrow stem cells failed to self-renew in long-term cultures (over 15 weeks) whereas OE-MSCs were still able to proliferate. This dramatic difference could be explained by dissimilarities in telomere regulation. It is well known that MSC telomeres are shortened and telomerase is not detected [66]. As a consequence, proliferation remains within the Hayflick's limit of 50 cell doublings [67,68]. In opposition, it has been demonstrated that olfactory stem cells retain telomerase activity and lack apoptotic activity [25].
Third, the association of OE-MSCs with a neuroepithelium is unique. Molecular signaling between neuroepithelium and lamina propria influences the olfactory pathway development [69]. During adulthood, cells migrating from one compartment to another can be observed, especially after an induced lesion (unpublished data). It can therefore be surmised that OE-MSCs are capable of crossing the basement membrane in order to differentiate into neurons and, possibly, replenish the olfactory epithelium after an extensive peripheral damage. A clue for this unusual behavior can be found in our list of differentially regulated genes (Table 2).The levels of transcripts encoding the protein S100A4 is 55 times higher in OE-MSCs, when compared to bone marrow stem cells. Interestingly, it has recently been shown that this calcium-binding protein is a key player in epithelial-mesenchymal transition and regeneration [70]. Otherwise, the increased expression of 18 transcripts encoding proteins expressed in neural cells clearly indicates that OE-MSCs are inclined to run along neural pathways. The intense NG2 immunostaining on every OE-MSC confirms this propensity. It has previously been shown that early postnatal NG2+ cells form neurospheres and are able to generate neurons in vitro [71] and in vivo [72]. Finally, this observation is substantiated by 2 of our previous reports [30,31] and by a parallel study that demonstrates that after transplantation in mouse lesioned hippocampus, human olfactory stem cells (1) differentiate into neurons, (2) restore long-term potentiation (LTP), and (3) induce memory recovery (unpublished data).
Conclusion
The olfactory mucosa is an easily accessible tissue that can be harvested in every individual under local anesthesia and without any side effect. Potentially, stem cells from the olfactory lamina propria may be used for autologous transplants, similarly to what has been done with nasal olfactory ensheathing cells in paraplegic patients [73]. The therapeutic potential of these stem cells has already been assessed using a rat model of Parkinson's disease [31]. The current study opens a new avenue for basic and clinical investigations in bone regeneration.
Footnotes
Acknowledgments
This work was financially supported by the Association Française contre les Myopathies (AFM), the Agence Nationale de la Recherche (ATOS), the Etablissement Francais du Sang (EFS 2005-11), an Inserm Osteoarticular research grant (PRO-A) and by the European Community (Key action 1.2.4-3 Integrated Project Genostem, contract No. 503161). Emmanuel Nivet is recipient of a doctoral fellowship from the Institut pour la Recherche sur la Moelle Epinière et l'Encéphale (IRME). The authors are grateful to Philippe Rosset (Orthopaedic Department of Tours University Hospital) for collecting bone marrow samples and Sophie Sourice for technical assistance in histology and scanning electron microscopy.
Author Disclosure Statement
Authors report no commercial association that might create a conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.