
Abstract
Early death of grafted bone marrow mesenchymal stem cells (MSCs) represents a major limit to their use in cell therapy of solid organs. It is well known that oxidative stress plays a major role in cell death. We have recently shown that the serotonin-degrading enzyme monoamine oxidase A (MAO-A) generates large amount of hydrogen peroxide (H2O2) responsible for cell apoptosis. Hydrogen peroxide generation requires 5-HT internalization into the cell and its degradation by MAO-A. In the present study, we investigated whether MAO-A is expressed in MSCs and we defined its role in serotonin-dependent MSCs apoptosis. RT-PCR analysis and western blots showed that the serotonin transporter (SERT) and the 2 MAO isoenzymes, A and B, are expressed in MSCs. As shown by enzyme assays using [14C]serotonin or [14C]β-phenylethylamine as selective MAO-A or MAO-B substrates, MAO-A is largely predominant in MSCs. Incubation of MSCs with the MAO substrate tyramine led to a time-dependent generation of H2O2 that was prevented by the MAO inhibitor pargyline. Finally, exposure of the cells to serotonin promoted an increase in MSCs apoptosis prevented by pargyline and the SERT inhibitor imipramine. The pro-apoptotic effect of serotonin was associated to a decrease in the expression of the anti-apoptotic factor Bcl-2. In conclusion, these results show for the first time that the 5-HT-degrading enzyme MAO-A is an important source of H2O2 in MSCs and plays a major role in 5-HT-dependent MSCs apoptosis.
Introduction
M
Recent studies demonstrated that MSCs are a target of 5-HT. Indeed, it has been shown that 5-HT, through activation of 5-HT2 receptors, regulate neuroplasticity [24] and proliferation of MSCs [25].
In the present study, we investigated whether MAO-A is expressed in MSCs and we defined its role in serotonin-dependent MSCs apoptosis.
Materials and Methods
Cell culture
Marrow aspirates were obtained from femurs cavity of Lewis rats (Harlan, France) weighing 180–200 g. Bone marrow from femurs cavity was flushed with minimum essential medium (MEM) (ABCYs, France) containing 10% fetal calf serum (FCS) and 1% penicillin/streptomycin (Invitrogen, Carlsbad, CA) and the cell suspension was centrifuged (400g, 5 min). Then, cells were plated in culture flasks (200,000 cell/cm2). Nonadherent cells were removed after 72 h and MSCs were recovered by their capacity to adhere highly to plastic culture dishes. MSCs were then routinely cultured and were used for the experiments after passage 3. Most adherent cells expressed CD90, CD29, CD106 and were negative for CD34, CD45, and CD31.
RT-PCR
Total ribonucleic acid (RNA) was isolated from MSCs, or H9C2 using nucleospin kit (Macherey Nagel, Germany) and cDNA was synthesized from 1 μg total RNA using SuperScript II reverse transcriptase (Invitrogen, Carlsbad, CA). Polymerase chain reaction (PCR) was performed in 20 μL reaction buffer containing 17 μL PCR Platinium Mix (Invitrogen), 1 μg cDNA, 1 μg of specific primers. PCR conditions were as follows: 4 min at 94°C, followed by cycles of denaturation (30 s, 94°C), 30 s annealing at 60°C, and 30 s extension at 72°C. PCR products were confirmed on a 1% agarose gel and visualized under UV light after ethidium bromide staining. Followed primers (sense/antisense) were used (Eurogentec, Belgium): SERT-F AGTGCTGTCAGAGTGTAAGGA, SERT-R GCGCCCAGGCTATGATGGTGTT; MAO-A-R: ATGACGGATCTGGAGAAGCC; MAO-A-F: TGCCTCACATACCACAGGAAC; MAO-B-R: TTAGATAATTTGTGTGGGTTGAGAGAA; MAO-B-F: AAGAAAACAAAAGAACCCAGAAATTATT.
For mRNA expression of SERT, H9C2 cells have been used as a positive control. The H9C2 cardiomyocyte-like cell line (American Type Culture Collection, Molsheim, France) is a rat embryonic myoblast-derived cell line commonly used as an in vitro model of cardiomyocyte biology that shows similar hypertrophic and apoptotic responses as those seen in primary adult and neonatal cardiomyocytes [26].
Apoptosis evaluation
Apoptosis was assessed by double staining cells with Syto 13 (1 μmol/L; Molecular Probes, Eugene, OR) and propidium iodide (6 μg/mL; Molecular Probes) for 15 min at 37°C. Cells were then examined with a fluorescent microscope, and apoptotic cells were characterized by condensed, fragmented nuclear regions. A total of 300 cells were counted for each condition.
Western blot
For western blot (WB), proteins were extracted from MSCs and prepared from rat brain and liver lysates. WB analyses were performed with samples normalized for protein concentration. Membranes were probed with anti-rabbit Bcl-2 (1:500; Santa Cruz Biotechnology, Santa Cruz, CA), anti-mouse Bax (1:500; Santa Cruz Biotechnology, Santa Cruz, CA), anti-rabbit MAO-A (1:400; Santa Cruz Biotechnology), anti-goat MAO-B (1:400; Santa Cruz Biotechnology), or anti-goat actin (1:1,000; Santa Cruz Biotechnology) antibodies. Following several washes in Tris-buffered saline-Tween (0.2%), membranes were incubated with anti-rabbit or anti-mouse or anti-goat secondary antibody (1:10,000; Santa Cruz Biotechnology). Expression of Bcl-2 and Bax is compared with expression of actin.
H2O2 production assay
As previously reported, the luminol-amplified chemiluminescence assay is a sensitive procedure to measure the specific contributions of MAO-A and MAO-B to H2O2 production [27].
H2O2 production was measured by chemiluminescence assay on MSCs scrapped in Hank's balanced salt sodium (HBSS; 5 μg) in the presence of luminol (30 μM) and horse radish peroxidase (0.1 U/μL) by using a thermostatically (37°C) controlled luminometer (Mithras, Berthold, France). The generation of chemiluminescence triggered with tyramine (30 μM), a common MAO substrate, was monitored every 20 s during 60 min.
Assays of MAO activity
Crude extracts of proteins (50 μg) were incubated at 37°C for 20 min, in the presence of 3.125 to 400 μM of [14C]serotonin or 200 μM of [14C]β-phenylethylamine to measure MAO-A and MAO-B activities, respectively [28]. To define the nonspecific activities, the MAO-A inhibitor clorgyline and the MAO-B inhibitor deprenyl were used (0.1 μM). The reaction was ended by the addition of 0.1 mL of HCl 4 N at 4°C. The reaction product was extracted (efficiency 92%) with 1 mL of ethyl acetate/toluene (v/v), and the radioactivity contained in the organic phase was counted in a liquid scintillation spectrometer.
Statistical analysis
Results are expressed as mean ± standard error of the mean (SEM). Statistical comparison of the data was performed using the t-test for comparison between 2 groups or one-way analysis of variance (ANOVA) and post hoc Tukey's test for comparison of >2 groups. A value of P < 0.05 was considered significant.
Results
Expression of 5-HT transporter and monoamine oxidases in MSCs
Serotonin transport into the cell and its degradation by MAO-A is necessary for H2O2 generation. RT-PCR performed on mRNA obtained from rat MSCs showed the expression of mRNA encoding for the 5-HT transporter SERT (serotonin transporter), and for the 2 isoforms of MAOs, MAO-A and MAO-B (Fig. 1, panels A and B, respectively). As shown in Figure 1C, western blot analysis performed on MSCs lysates revealed 2 proteins with the apparent molecular weight expected for MAO-A and MAO-B (∼61 and 60 kDa, respectively). The expression of both MAO-A and MAO-B was confirmed by radioenzyme assay using [14C]5-HT and [14C]β-PEA as substrates for MAO-A and MAO-B, respectively (Fig. 1D, upper panel) and by dose response using [14C]5-HT (Fig. 1D, lower panel). Results indicated that the 2 isoforms were found in MSCs with a higher expression of the MAO-A.
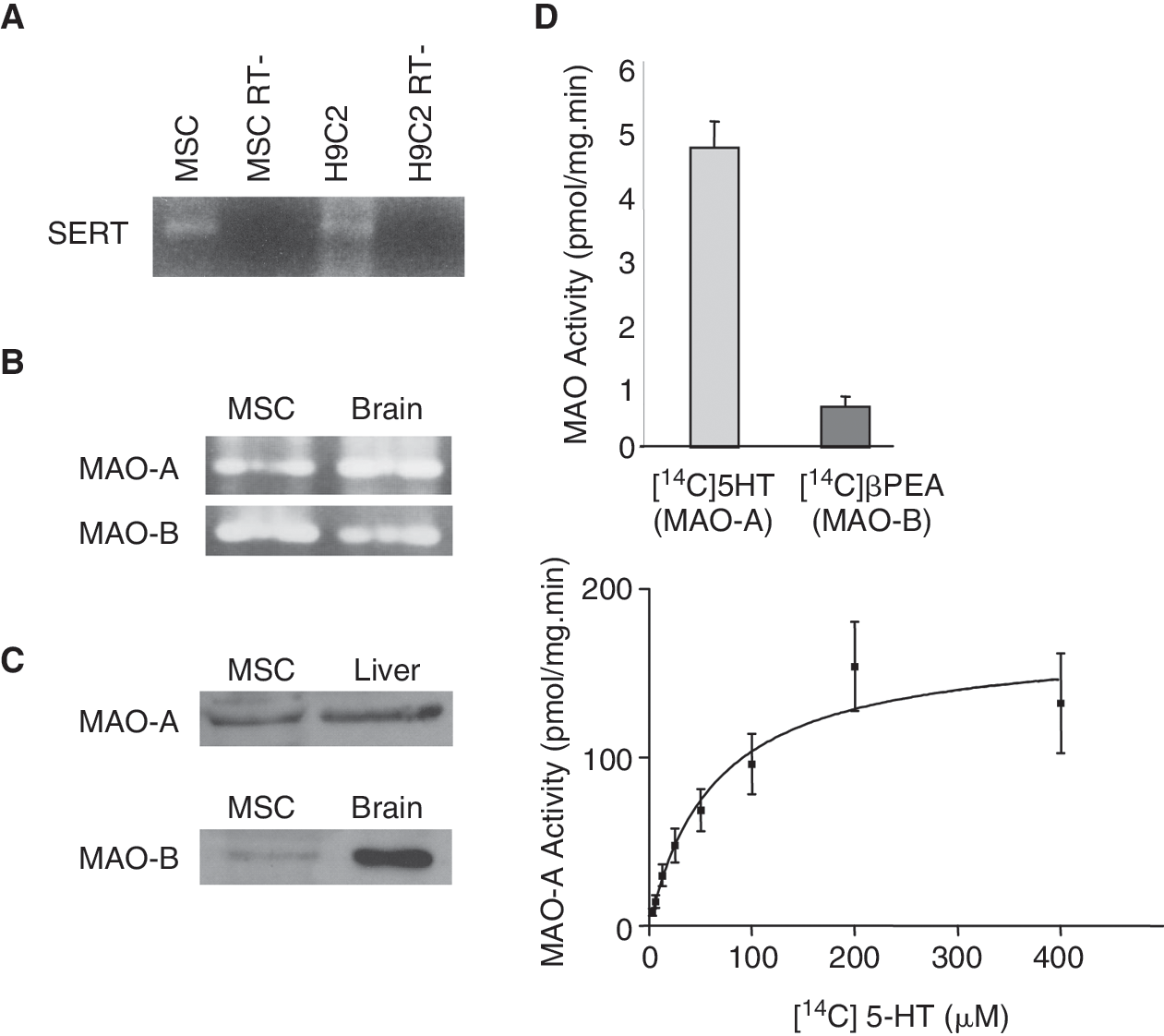
Serotonin transporter (SERT) expression and monoamine oxidases (MAOs) expression/activities in mesenchymal stem cells (MSCs). (
H2O2 production by MAOs in MSC
In order to determine whether MAOs are a source of ROS in MSCs, H2O2 production was assessed using a continuous luminol-amplified chemiluminescence assay. We have previously shown that this technique allows the measurement of H2O2 generation by MAOs in intact cells using the MAO-A/MAO-B substrate tyramine [27]. As shown in Figure 2, incubation of MSCs with tyramine (30 μM) led to a time-dependent increase in chemiluminescence. This effect was prevented by preincubation of MSCs with the irreversible MAO inhibitor pargyline (5 μM). We observed that ROS generation by tyramine was prevented by pargyline by 78% (94.8–21). Tyramine is a MAO substrate but it has also been shown that it can be metabolized by other amino oxidases, in particular, the semicarbazide-sensitive amine oxidase (SSAO). Consequently, it is possible that residual H2O2 generation may be related to the SSAO activity. H2O2 generation was also observed using higher tyramine concentrations (300 μM and 2 mM). However, this effect was observed only within 5 min after tyramine addition and rapidly declined with time probably because of MSCs death. This hypothesis is supported by our previous results showing that tyramine (50 μM, 24 h) induced MSCs apoptosis measured by MTT assay (data not shown). According to our findings, Marcocci et al. demonstrated that tyramine at concentrations from 100 to 500 μM altered the mitochondrial membrane permeability transition [29], an event generally associated with cell apoptosis [30].
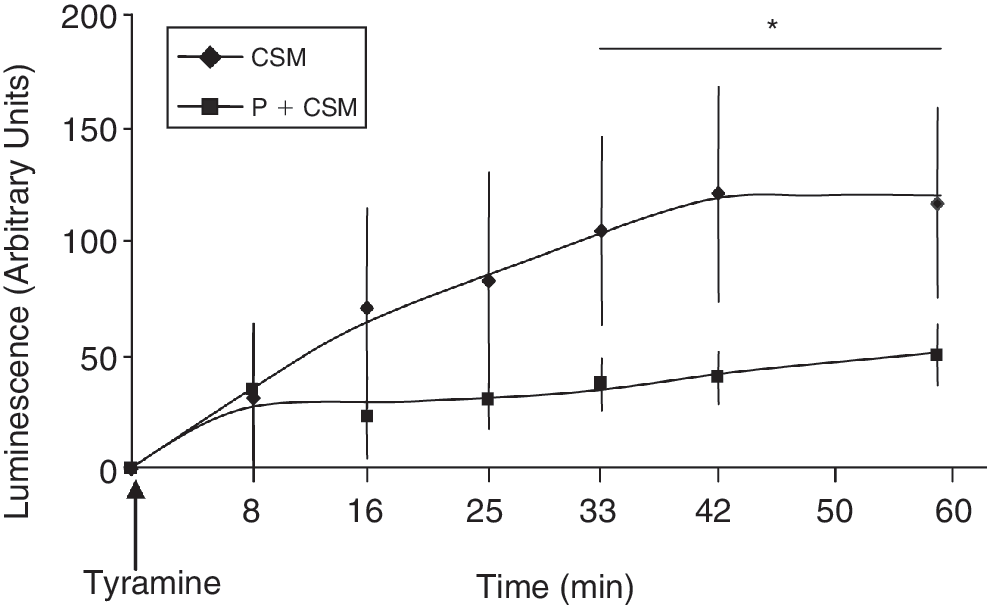
Monoamine oxidase (MAO)-dependent H2O2 production in mesenchymal stem cells (MSCs) lysates. Generation of chemiluminescence was monitored for 60 min after the addition of tyramine (30 μM) to MSCs as described in “Materials and Methods” section. MSCs were preincubated with pargyline (5 μM, 20 min) before tyramine addition. The plot is representative of 4 separate experiments. *P < 0.05 versus pargyline-treated MSCs.
These results indicate that MAOs are able to produce H2O2 during substrate degradation.
5-HT treatment induced apoptosis of MSC
In these experiments, we investigated whether MAOs are involved in 5-HT-dependent MSCs apoptosis. Cell apoptosis was evaluated by analysis of DNA fragmentation and the measure of cell incorporation of 2 fluorescence dyes SYTO-13, a permeant DNA intercalating green-colored probe, and propidium iodide, a nonpermeant intercalating orange probe. Apoptotic and necrotic cells were discriminated by green (SYTO-13) and orange (propidium iodide) staining, respectively. Cell treatment with increasing concentrations of 5-HT for 24 h induced a significant increase (25% to 40% for 5-HT 10 μM) in the number of SYTO-13-stained cells as compared to untreated cells (Fig. 3A). In contrast, 5-HT treatment did not increase cell staining by propidium iodide, the marker of necrotic cells. Exposure of MSCs to 5-HT (10 μM) at different times (3, 6, 12, and 24 h) induced a time-dependent increase in apoptosis (Fig. 3B).
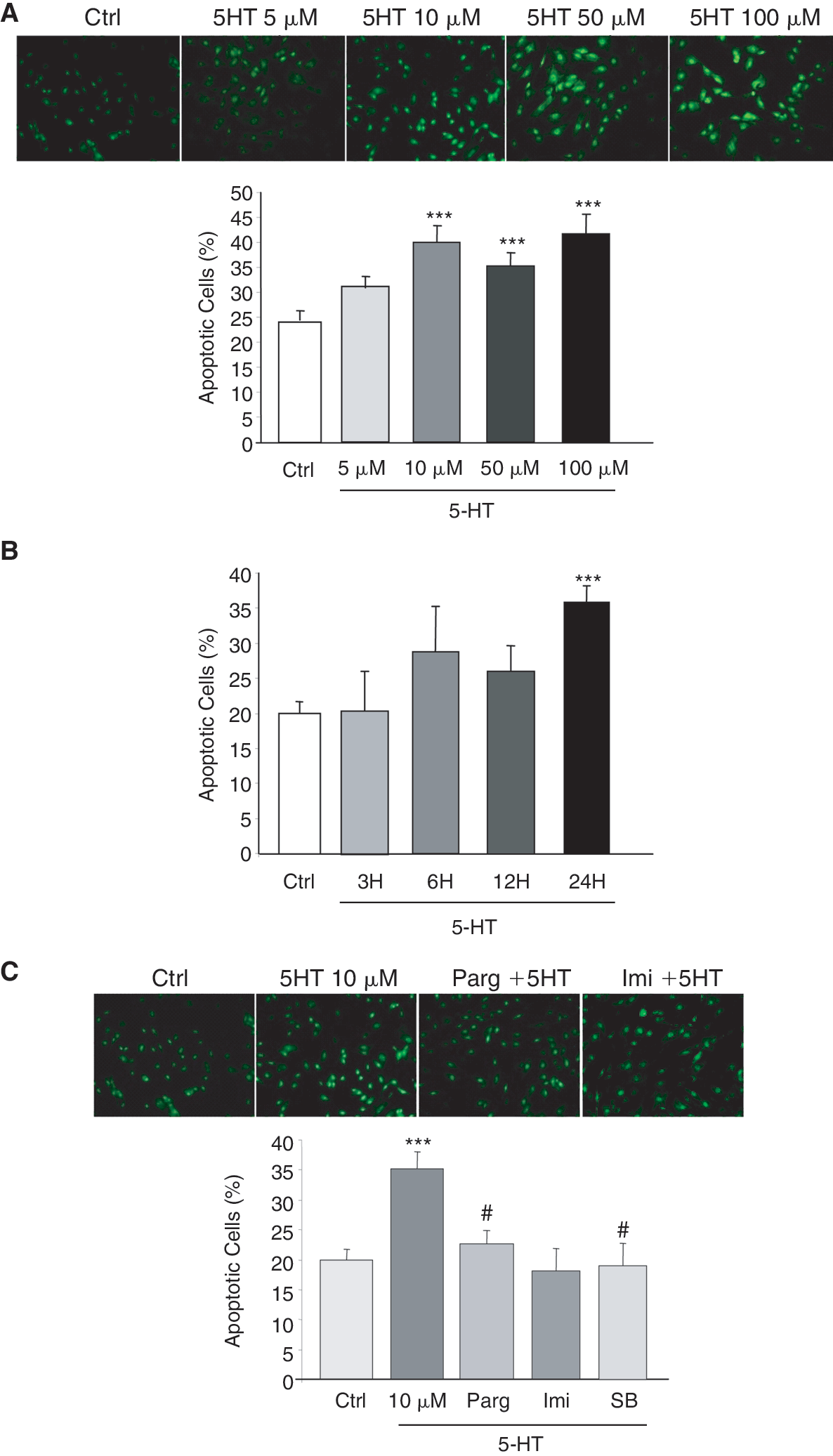
Time- and dose-dependent effects of 5-HT on apoptosis. Cell nuclei were double-stained with SYTO-13 and propidium iodide, allowing discriminating normal cells from cells undergoing apoptosis or necrosis by fluorescence microscopy. (
As we and others have demonstrated that 5-HT could induce apoptosis through a receptor-independent pathway, we have decided to clarify the involvement of MAOs in 5-HT-induced apoptosis. MSCs have been pretreated with the pargyline (5 μM, 24 h) before 5-HT treatment and results indicated that inhibition of MAO activity prevented apoptosis (Fig. 3C). Furthermore, use of the imipramine (10−7 M), a serotonin uptake inhibitor, and SB206553 (0.1 μM, 1 h), a 5-HT2B antagonist, also prevented the pro-apoptotic effect of 5-HT on MSCs.
These results show that, in order to induce MSCs apoptosis, 5-HT needs to be internalized into the cells and metabolized by MAO-A and that 5-HT2B receptor could play a role in this mechanism.
Effect of serotonin on Bcl-2 and Bax expression and mitochondrial cytochrome c release in MSCs
Apoptotic processes could be associated to mitochondrial dysfunction involving members on the Bcl-2 protein family such as the anti-apoptotic protein Bcl-2 and the pro-apoptotic protein Bax. Exposure of MSCs to 5-HT (10 μM, 24 h) promoted an increase in cell apoptosis (Fig. 3B) that was associated to a decrease in Bcl-2 expression (Fig. 4A). This effect was partially prevented when MSCs were pretreated with pargyline (5 μM, 24 h). We next examined the effect of serotonin on Bax expression (Fig. 4B). Western blot analysis showed that incubation of MSCs with 5-HT did not modify the level of Bax protein.

Effect of 5-HT on Bcl-2/Bax expression and cytochrome c release in mesenchymal stem cells (MSCs). (
Release of cytochrome c from mitochondria to cytosol is a critical step of the mitochondrial-dependent cell apoptosis. As shown in Figure 4C, MSCs treatment with 5-HT (10 μM, 24 h) induced a slight increase in cytosolic cytochrome c as defined by western blotting.
Altogether, these results indicate that 5-HT induces MSCs apoptosis through a MAO-A-dependent mechanism. This effect could involve ROS generation concomitant to a decrease in the amount of the anti-apoptotic factor Bcl-2 that could contribute to cell apoptosis.
Discussion
In the present work, we showed the expression of the 5-HT-degrading enzyme MAO-A in bone marrow MSCs and its role in H2O2 generation. In addition, we demonstrated that MAO-A is involved in 5-HT-dependent MSC apoptosis.
Monoamine oxidase A is widely distributed in the body. While the function of MAO-A has been extensively investigated in the central nervous system, less is known concerning its role in the periphery. As in neuronal cells, MAO-A in the liver [31] and in the lung [32] is involved in the clearance of 5-HT. In other cell types, like cardiomyocytes, we showed that MAO-A also participates in the cell effects of 5-HT [15]. The MAO-A-dependent effects of 5-HT requires 5-HT transport into the cell and its metabolism by MAO-A. Our results show for the first time that this mechanism of action of 5-HT exists in MSCs. Indeed, we showed that 2 partners, necessary for the receptor-independent effects of 5-HT, SERT, and MAO-A, are expressed in MSCs. As previously reported for cardiomyocytes, 5-HT induced MSC apoptosis and this effect was prevented by the SERT inhibitor imipramine, by the MAO inhibitor pargyline, and more surprisingly by SB206553. We may explain these very intriguing results by interactions between 5-HT receptors and SERT. Indeed, studies have showed that activation of 5-HT receptors could trigger the phosphorylation of SERT by PKC, PKG, or p38. These kinases modulate the SERT membrane expression and, consequently, promote a reduction or an acceleration of the kinetics of 5-HT uptake [33 –35]. Furthermore, in platelets, SERT-mediated accumulation of 5-HT results in “transamidation” to small GTPases such as Rho-A and Rho-4 [36]. In this case, 5-HT2A receptors synergizes with SERT. Consistent with these results, we also showed that 5-HT decreased the expression of the anti-apoptotic protein Bcl-2 through a MAO-A-dependent mechanism. These results show that 5-HT and MAO-A may behave as pro-apoptotic factors in MSCs. In our experiments, we observed only a slight increase of cytochrome c release after 5-HT treatment. This could be explained by the kinetics of the experiments. Indeed, cytochrome c release from mitochondria occurs in the early step of the apoptosis. After its release, cytochrome c is associated with Apaf-1 to form the apoptosome that will induce the caspase activation. Our experiments were done 24 h after 5-HT treatment. It is conceivable that, at this time, cytochrome c may be partially cleaved in apoptotic cells. Nevertheless, the slight increase in cytosolic cytochrome c supports the involvement of mitochondrial dysfunction in apoptosis of MSCs.
The relevance of 5-HT/MAO-A-mediated apoptosis of MSCs in vivo is supported by different observations. In the periphery, 5-HT is mainly stored in platelet and released by activated platelets. Platelet activation occurs in different pathological situations manageable by cell therapy and characterized by the increase in circulating or local 5-HT. In the heart, several studies showed that 5-HT accumulates during ischemia–reperfusion, and contributes to the progression of myocardial injury and dysfunction [37 –39]. In addition, circulating 5-HT concentrations also increase in chronic diseases such as diabetes [40,41] and atherosclerosis [42,43] and could be elevated because of blood platelets involvement in immune inflammatory processes [44 –46]. Based on our results, it is conceivable that the 5-HT/MAO-A pathway and the consequent H2O2 generation may contribute to the death of MSCs after intraparenchymal injection. Further studies will be necessary to confirm this possibility.
The demonstration of the expression of MAO-A and of the apoptotic effects of 5-HT in MSCs may exceed the field of cell therapy. Indeed, during the last years, several studies showed that 5-HT plays a critical role in proliferation of MSCs [25] and in the function of MSC-derived cells such as osteoblasts [47] and fibroblasts [48]. In addition, recent studies indicate that 5-HT also plays an important role in differentiation and survival of bone marrow hematopoietic cells [49]. Recent studies reported that ROS are implicated in the reduction of MSCs numbers and differentiation capacity associated with aging [50,51]. Interestingly, it has been also shown that the tissue MAO-A expression and activity increases with age [52,53]. Therefore, MAO-A appears to be a potential candidate to the increase in H2O2 generation during MSCs aging. Based on these and our results, it is conceivable that 5-HT and MAO may play an important role in the regulation of bone marrow cells in physiological and pathological situations.
In conclusion, we have shown for the first time the expression of the 5-HT-degrading pathway SERT/MAO-A in MSCs and its involvement in 5-HT-dependent MSCs apoptosis. These results open new perspective in the comprehension of the role of MAO-A in the regulation of 5-HT effects in MSCs and its involvement in physiological and pathological processes.
Footnotes
Acknowledgments
This work was supported in part by the INSERM and grants from the National Research Agency (ANR; Grant under program Physiopathologie des Maladies Humaines, project SYNMESCARI), the Région Midi-Pyrénées, and the Association Française contre les Myopathies. The research leading to these results has received funding from the European Community's Seventh Framework Programme [FP7/2007-2013] under grant agreement no. HEALTH-F5-2008-223007 STAR-T REK.
Author Disclosure Statement
No competing financial interests exist.