
Abstract
Liver tissue engineering with hepatic stem cells provides a promising alternative to liver transplantation in patients with acute and chronic hepatic failure. In this study, a three-dimensional (3D) bioscaffold was introduced for differentiation of rat bone marrow mesenchymal stem cells (BMSCs) into hepatocytes. For hepatocyte differentiation, third passage BMSCs isolated from normal adult F344 rats were seeded into collagen-coated poly(lactic-co-glycolic acid) (C-PLGA) 3D scaffolds with hepatocyte differentiation medium for 3 weeks. Hepatogenesis in scaffolds was characterized by reverse transcript PCR, western blot, confocal laser scanning microscopy (CLSM), periodic acid-Schiff staining, histochemistry, and biochemical assays with hepatic-specific genes and markers. A monolayer culture system was used as a control differentiation group. The results showed that isolated cells possessed the basic features of BMSCs. Differentiated hepatocyte-like cells in C-PLGA scaffolds expressed hepatocyte-specific markers [eg, albumin (ALB), α-fetoprotein, cytokeratin 18, hepatocyte nuclear factor 4α, and cytochrome P450] at mRNA and protein levels. Most markers were expressed in C-PLGA group 1 week earlier than in the control group. Results of biocompatibility indicated that the differentiated hepatocyte-like cells grew more stably in C-PLGA scaffolds than that in controls during a 3-week differentiation period. The significantly higher metabolic functions in hepatocyte-like cells in the C-PLGA scaffold group further demonstrated the important role of the scaffold. Conclusion: As the phenomenon of transdifferentiation is uncommon, our successful transdifferentiation rates of BMSCs to mature hepatocytes prove the superiority of the C-PLGA scaffold in providing a suitable environment for such a differentiation. This material can possibly be used as a bioscaffold for liver tissue engineering in future clinical therapeutic applications.
Introduction
E
Numerous studies using liver tissue engineering for treatment of end-stage liver disease demonstrated survival of hepatocytes transplanted heterotopically, improvement in survival of transplanted hepatocytes with hepatotrophic stimulation, and enhancement in proliferation [14 –17]. However, the main challenge has been the limited number of viable and functional cells after implantation because of mature hepatocytes almost losing their differentiation and proliferation capabilities, and poor oxygen and nutrient supply during the initial period after transplantation due to the poor microenvironment for implanted cells [18].
To address these issues, many researchers attempted to establish three-dimensional (3D) tissues with biomimetic materials and replace mature hepatocytes with stem cells [10,19 –21]. Recent evidence indicated that cells forming the tissue to be regenerated are necessary for efficient tissue regeneration and stem cells are promising in both proliferation activity and differentiation potential [22,23]. Bone marrow mesenchymal stem cells (BMSCs) possess great potential for liver tissue engineering because autologous BMSCs can be isolated easily, proliferated extensively in vitro, and transdifferentiated into hepatocyte-like cells for transplantation [22]. We have previously isolated, from adult rat's bone marrow, a subset of stem cells that can be differentiated into hepatocyte-like cells in vitro with expression of hepatocyte-specific genes and proteins [24]. More importantly, these cells were able to metabolize urea and synthesize ALB in vitro. Recently, it was shown that a population of cells in the bone marrow expressing mesenchymal stem cell antigens could differentiate in vitro into hepatocyte-like cells, which exhibit highly differentiated hepatocyte functions [22]. We also showed that AHF in rats can alter or “prime” the bone marrow to increase the number of hepatic progenitor cells.
Other recent studies also have demonstrated that bone marrow-derived hepatic stem cells may play a major role in liver regeneration following acute or chronic hepatic failure. Differentiation of BMSCs into hepatocytes in monolayer or two-dimensional (2D) culture systems is now well established [25 –27], but challenges still need to be overcome to develop functional hepatocytes suitable for implantation from BMSCs.
Our recent study demonstrated that collagen-modified PLGA (C-PLGA) scaffolds are superior to the 2D culture system in supporting growth and proliferation of primary human hepatocytes, and selected hepatic metabolism functions including ALB secretion, urea synthesis, and ammonia clearance [28]. Many other studies confirmed that 3D scaffolds can significantly improve differentiation of embryonic stem cells into hepatocytes and provide an environment for liver regeneration-induced hepatocyte-like cells [21,29,30].
To demonstrate our hypothesis that BMSC can be differentiated into hepatocyte-like cells within designed C-PLGA scaffolds, with improved viability and hepatocyte-specific functions, we used in this study a combination of methods. RT-PCR, western blot, confocal laser scanning microscopy (CLSM), periodic acid-Schiff (PAS) staining, histochemistry, and biochemical assays with hepatic-specific genes and markers were all performed to show the ability and importance of 3D bioscaffolds to support differentiation.
Materials and Methods
Chemicals and reagents
Chemicals for cell isolation and culture were purchased from Sigma Chemical Corporation (St. Louis, MO). Dulbecco's modified Eagle's medium (DMEM), Iscove's modified Dulbecco's medium (IMDM), rat tail collagen I, antibiotic–antimycotic, trypsin–ethylenediaminetetraacetic acid (EDTA), and fetal bovine serum (FBS) were purchased from Invitrogen Corporation (Carlsbad, CA). Bovine serum albumin (BSA), MTT assay kit, dimethylsulfoxide (DMSO), ITX mixture, and oncostatin (OSM) were from Sigma Chemical Corporation (St. Louis, MO). Hepatocyte growth factor (HGF) was from Cell Science (Canton, MA), regular culture dishes and flasks were purchased from BD Falcon (Franklin Lakes, NJ). For enzyme-linked immunosorbent assay (ELISA) and western blot, rat ALB ELISA kit and antibodies against ALB were purchased from Bethyl, Inc. (Montgomery, TX). Other reagents for western blot were purchased from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA) and Bio-Rad Laboratories (Hercules, CA).
Collagen-coated PLGA scaffold preparation
Collagen-coated porous scaffolds were prepared from poly(lactic-co-glycolic acid) (PLGA molar ratio of 85:15; Sigma Chemical Corporation., St. Louis, MO) and type I collagen from rat tail (Gibco BRL, Grand Island, NY) by our previously described method [28]. In brief, PLGA (3 wt%) was dissolved in DMSO and frozen at −20°C. The frozen polymer solution was immersed in a precooled 30 wt% ethanol aqueous solution, and the solvent DMSO was extracted and replaced with the ethanol solution, a nonsolvent for PLGA. Subsequently, the ethanol was removed by drying at room temperature and PLGA porous scaffolds were obtained. A foam-like collagen solution was introduced into the PLGA scaffolds and freeze-dried at −20°C to form a porous sponge. To control the pore size of PLGA scaffolds within a resultant range of 150–350 μm diameter, 2-step coarsening was used. In brief, after coarsening at 25°C for 5 h, the system was cooled down to 20°C and coarsened again for another 2.5 h. The solvents were then solidified at −20°C for 2 h, and were removed by freeze-drying. For cell culture and transplantation experiments, the scaffolds were cut into discs of 10 mm diameter and 3 mm thickness. C-PLGA scaffolds were sterilized overnight in 70% ethanol and washed 3 times in phosphate-buffered saline (PBS) before cell culture. For characterization of their inner microstructure with scanning electron microscopy (SEM, JEOL JEM-200CX, Japan), the 3D C-PLGA scaffolds were fractured in liquid nitrogen. The diameter of the pores was measured and averaged from the SEM images by a graph calculator software, SMILE view [31].
Rat bone marrow mesenchymal stem cells isolation
Bone marrow cells were harvested by flushing the diaphysis of femurs and tibias from Fischer 344 rats with PBS containing 1% BSA, 100 U/mL penicillin and 100 μg/mL streptomycin. Following washing and resuspending in 5 mL PBS, the cells were carefully laid on 5 mL Ficoll-Paque (GE Healthcare, Uppsala, Sweden). After centrifugation at 400g at room temperature for 30 min, the cells were harvested from the plasma–Ficoll interface and washed 3 times with PBS and maintained at 4°C for further use [24]. BMSCs were allowed to attach in DMEM medium supplemented with 10% FBS overnight at 37°C. Floating cells were washed out on the second day. All remaining cells were maintained in fresh DMEM supplemented with 10% FBS culture medium. The animals received humane care according to the guidelines prepared by the National Institute of Health, USA.
Surface marker characterized by flow cytometry
For characterization of BMSCs by flow cytometry, the first, fourth, and seventh passage BMSCs with 1 × 106 cells were incubated with direct PE- or FITC-conjugated mouse anti-rat monoclonal antibodies recognizing CD29 (ABR Affinity BioReagents, CO), CD45 (Invitrogen, Carlsbad, CA), CD90, and β-2 microglobulin (β2m, Santa Cruz Biotechnology, Santa Cruz, CA) for 20 min in the dark at room temperature, followed by washing and resuspending in PBS. The immunoglobulin isotype incubation was used as a negative control. FACS analysis was performed with a FACSCalibur system (Beckman Coulter FC500).
Hepatocyte differentiation with C-PLGA scaffolds
The BMSCs seeding in scaffolds was performed as previously described [28]. C-PLGA scaffolds were placed and immobilized by injection needles in 24-well culture plates. The third passage BMSCs with 1 × 106 cells (BMSCs suspension 5 × 106 cells/mL) were directly dropped onto the surface of the each scaffold. While agitating the plate, vacuum was applied for 5 min and then the pressure returned to ambient. Additional vacuum/aeration cycles were performed until the scaffold absorbed no further cells suspension. The scaffold was then removed to a new well with 2 mL serum-free hepatocyte differentiation medium (IMDM supplemented with 20 ng/mL hHGF, 100 μM dexamethasone, 50 mg/mL ITX+ premix, 100 U/mL penicillin, and 100 μg/mL streptomycin) and maintained in the same culture medium for an initial 2 weeks. Thereafter, the cells were changed into a maturation medium (IMDM supplemented with 20 ng/mL OSM, 100 μM dexamethasone, 50 mg/mL ITX+ premix, 100 U/mL penicillin, and 100 μg/mL streptomycin) and maintained for an additional week. As a control monolayer culture system, BMSCs with 5 × 105 cells (BMSCs suspension 2 × 104 cells/cm2) were differentiated in T-25 culture flasks and cover slides in 24-well culture plates (cell concentration was same with in T-25 culture flasks). The culture medium was replaced with fresh medium twice a week. Before and after differentiation at day 7, 14, and 21, the differentiated BMSCs were harvested and characterized for hepatic function with different approaches including RT-PCR, western blot, CLSM, PAS staining, histochemistry, and biochemistry. The experiment in hepatocyte differentiation with C-PLGA scaffolds was repeated >3 times.
RT-PCR
Total RNA was extracted from undifferentiated rat BMSCs, various stages of the differentiated hepatocytes cultured in scaffolds and culture flasks as well as primary hepatocytes using TRIZOL (Invitrogen, Carlsbad, CA) according to the manufacturer's protocol. Total RNA was reverse-transcribed and cDNA was synthesized using Quantitect reverse transcription reagent (Qiagen, Valencia, CA). In brief, the purified 1.0 μg RNA was incubated in gDNA wipeout buffer at 42°C for 2 min to remove contaminating genomic DNA and the RNA sample was then transcribed at 42°C for 15 min. The reverse-transcribed cDNA was subsequently inactivated at 95°C for 3 min and amplified by PCR using rat liver-specific gene primers (Invitrogen, Carlsbad, CA) as listed in Table 1. RT-PCR was performed as described previously [28].
Abbreviations: ALB, Albumin; AFP, α–fetoprotein; G6PD, glucose-6-phosphate dehydrogenase; CK18, cytokeratin 18; CK19, cytokeratin 19; HNF-4, hepatocyte nuclear factor 4; GAPDH, glyceraldehyde phosphate dehydrogenase.
Western blot
The cultured cells were lysed by a lysis buffer (Pierce, Rockford, IL) containing protease inhibitor and EDTA. Protein concentration in cell lysates was measured using the Coomassie Plus—The Better Bradford™ Assay Kit (Pierce, Rockford, IL). About 20 μg of total protein was subjected to 4%–12% Bis–Tris gel electrophoresis and transferred to nitrocellulose membranes. Blots were blocked with 5% nonfat milk in Tris-buffered saline containing Tween-20. Blot membranes were incubated with anti-rat primary antibodies of liver functional markers including ALB, cytochrome P450 (P450), cytokeratin 18 (CK18), and cytokeratin (CK19) at 4°C overnight (final concentration of different antibodies according to the manufacturer's recommendations). Membranes were then washed, and specifically bound immunoglobulins were detected by incubation with horseradish peroxidase–conjugated secondary antibody for 1 h at room temperature. Protein bands were visualized by enhanced chemiluminescence systems (Pierce) and the images were captured on medical X-ray film (Phoenix Research, Hayward, CA).
Evaluation of cell survival and scaffold biocompatibility
To evaluate biocompatibility and efficiency of C-PLGA scaffolds in BMSC differentiation into hepatocyte-like cells, we observed the survival of differentiated hepatocyte-like cells in scaffolds and in a monolayer culture system. The cells' survival was determined by Vybrant CFDA SE cell tracer kit (Invitrogen, Carlsbad, CA) during the period of differentiation. Each sample was tested in triplicate at the same time. Ten millimolar CFDA SE stock solution was prepared by dissolving 500 μg CFDA SE in 90 μL of DMSO. A 10 μM CFDA SE working solution was obtained by diluting in PBS. For labeling the differentiated BMSCs in PLGA scaffolds, the medium was removed and pre-warmed (37°C). CFDA SE working solution was added into the cultures, and then the cultures were incubated with 5% CO2 at 37°C for 15 min. We replaced the loading solution with fresh, pre-warmed (37°C) DMEM containing 10% FBS and incubated the cultures with 5% CO2 at 37°C for 30 min. Subsequently, we washed with PBS several times and fixed with 4% paraformaldehyde for 15 min at room temperature and rinsed with PBS. Sections were cut and mounted on glass slides with Xylene-Soluble Mounting Medium and covered with coverslips. Cells labeled with CFDA SE were visualized by CLSM (Olympus IX 70 + Fluoview 300), using standard fluorescein wavelengths (fluorescence excitation/emission maxima: 488/517 nm).
The density of cells attached in scaffolds was evaluated by using an MTT colorimetric assay. In brief, the third passage BMSCs were cultured in C-PLGA scaffolds (6 × 105 cells/scaffold) and monolayer culture system (5 × 105 cells/T-25 flask, 2 × 104 cells/cm2). The scaffolds with differentiated cells were washed with PBS 3 times, transferred into a new 24-well plate containing 5:1 ratio of media and MTT solution (0.5%, Sigma, Germany 5 mg/mL in PBS), respectively, and incubated for 2 h at 37°C. After removing the culture media, 0.5 mL of extraction solution (DMSO) was added. The scaffolds were washed extensively by pipetting up and down repeatedly to allow total color release. The absorbance of the supernatant was read with a microplate reader with a 630-nm reference wavelength and 570 nm test wavelength. Cell number was determined through a standard curve that was established by using a known number of cells counted by a hemocytometer. The cells' density was analyzed once a week. Each sample was tested in triplicate at the same time. The cell growth curve was made with the time as abscissa (x-coordinate) and the cells number as ordinate (y-coordinate) [28].
Biochemical assay for hepatocyte-enriched metabolic functions
For evaluation of hepatocyte-enriched metabolic functions, ALB secretion, urea synthesis, ammonia elimination, and diazepam clearance from BMSC-derived hepatocyte-like cells in both of our differentiation systems were assayed. ALB secreted from BMSC-derived hepatocyte-like cells on C-PLGA scaffolds was tested by ELISA to evaluate hepatic function at various stages of differentiation (in weeks 0, 1, 2, and 3). The culture supernatant harvested from differentiated cells was tested using a commercially available kit. The assay was performed as described previously [28]. To evaluate the hepatic metabolic function of differentiated hepatocyte-like cells from BMSCs in bioscaffolds, the culture supernatants were measured for ammonia concentration determined by an enzymatic
Histochemical and PAS staining
The morphology of differentiated hepatocyte-like cells in C-PLGA scaffolds was characterized by hematoxylin–eosin (H&E) staining. C-PLGA scaffolds containing differentiated cells were fixed with 4% formaldehyde in PBS for 3 h at 4°C and then embedded in paraffin, sectioned into longitudinal and horizontal cross sections (5 μm thickness). The specimens were stained with H&E and examined under a light microscope (Nikon E800). For glycogen detection, PAS staining was performed to determine glycogen expression in differentiated hepatocyte-like cells in C-PLGA scaffolds. Specimen slides fixed with 4% paraformaldehyde were rinsed in slowly running tap water, and then immersed in periodic acid solution for 5 min at room temperature. After that, the specimen slides were rinsed in distilled water and immersed in Schiff's reagent for 15 min at room temperature. Followed by washing in running tap water for 5 min, slides were counterstained in hematoxylin solution for 90 s and rinsed in running tap water for 15–30 s, dried, and examined microscopically.
Statistical analysis
Data were evaluated by Student's t-test for each pair using software SPSS version 16.0. The level of significance for all statistical analyses was P < 0.05.
Results
Scaffold properties
The morphology of the C-PLGA scaffold under SEM is shown in Figure 1. C-PLGA scaffolds with round-shaped pores and an average pore diameter of 150–350 μm were successfully prepared. Their porosities were 94%. The images showed that pores possessed a highly interconnected network, which is important for cell infiltration and nutrient transportation.
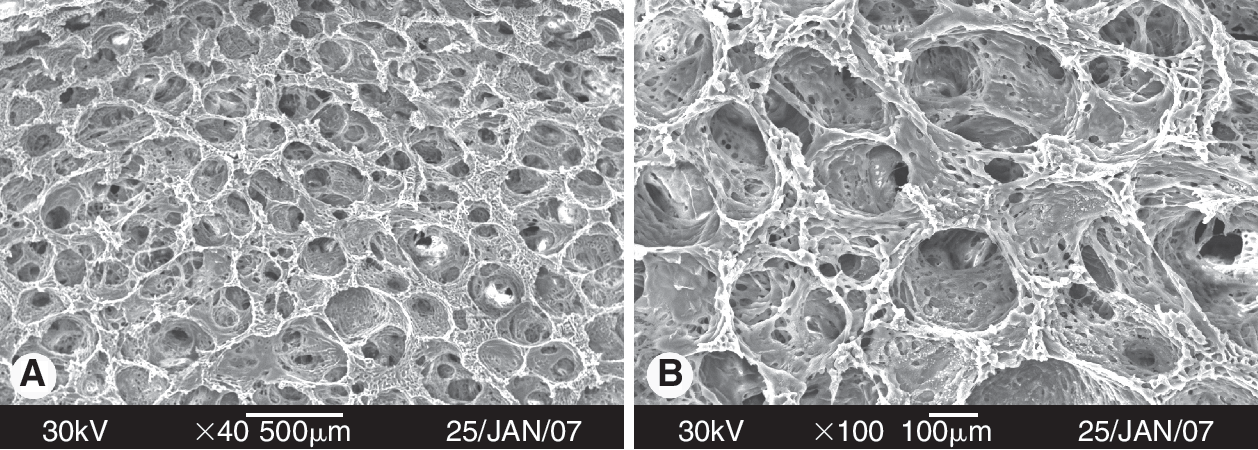
Scanning electron microscopy (SEM) images of collagen-coated poly(lactic-co-glycolic acid) (C-PLGA) scaffolds with pore diameters of 150–350 μm. The estimated porosity reached about 94% (original magnification:
Phenotype characteristic of BMSCs
The results of phenotypic properties of BMSCs (Fig. 2) showed that BMSCs were positive for CD29 (97.5%–98.1% within passage 1 to passage 7) and CD90 (93.8%–98.3% within passage 1 to passage 7), but negative for CD45 (0.4%–0.6% within passage 1 to passage 7) and β2m (0.3%–0.8% within passage 1 to passage 7). These data suggested that the cells used for differentiation in this study exhibited the characteristics of BMSCs.

Immunophenotyping of rat bone marrow mesenchymal stem cells (BMSCs) using flow cytometry. Bone marrow mesenchymal stem cells (BMSCs) were positive for CD29 (97.5%–98.1%) and CD90 (93.8%–98.3%) within passage 1 to passage 7. These cells were negative for CD45 (0.4%–0.6%) and β2m (0.3%–0.8%) within passage 1 to passage 7. The results shown are representative of 3 independent experiments (P-1, passage 1; P-4, passage 4; P-7, passage 7).
Results of RT-PCR
The results of RT-PCR shown in Figure 3A indicated that after 3 weeks of differentiation, differentiated hepatocyte-like cells were positive for several liver functional genes, including ALB, AFP, CK18, CK19, G6PD, and HNF-4, in both groups of C-PLGA scaffolds and control. All genes were expressed 1 week earlier in differentiated hepatocyte-like cells in C-PLGA scaffolds versus those in controls. ALB, AFP, CK18, and CK19, in particular, were expressed in the first week of differentiation in the scaffold group. In contrast, these genes were expressed in the second week in monolayer culture group.
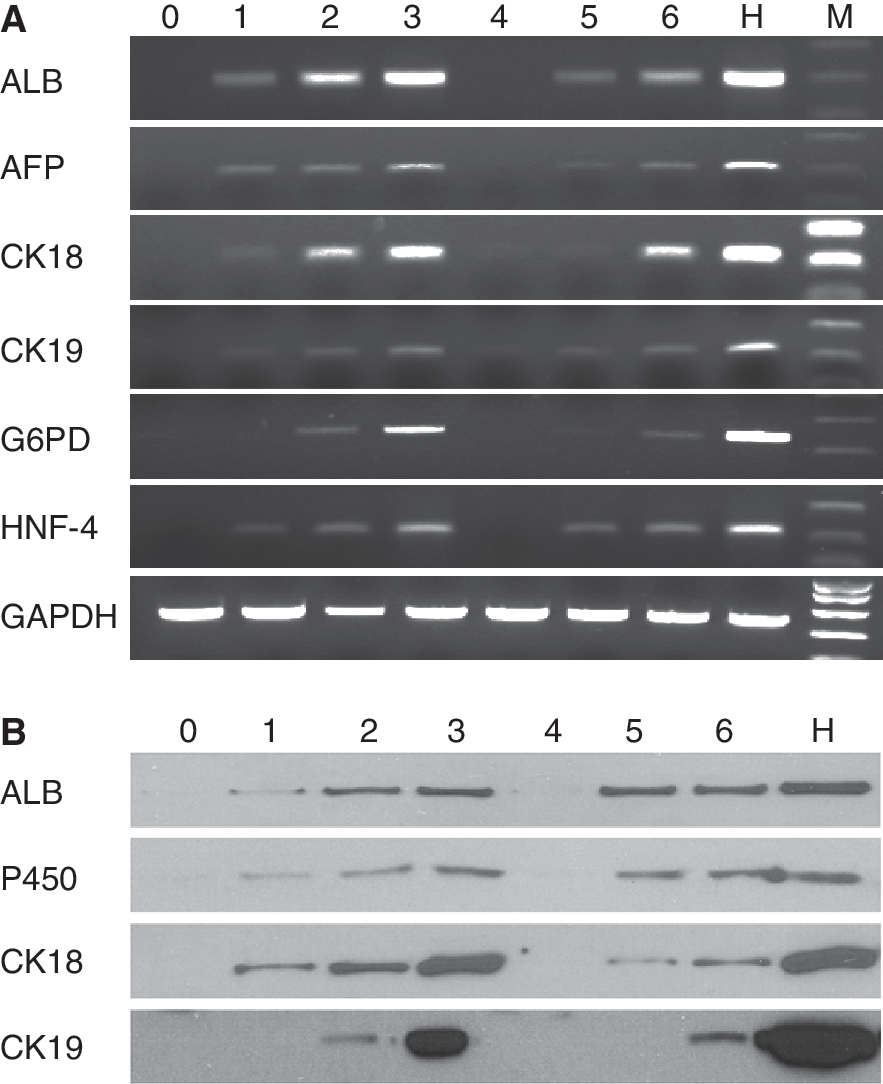
Results of RT-PCR (
Results of western blot
Western blot results shown in Figure 3B indicated that differentiated hepatocyte-like cells were positive for 4 liver-enriched functional markers including ALB, P450, CK18, and CK19 after 3 weeks of differentiation from both 2 different hepatocyte-like cells. Three proteins (ALB, P450, and CK18) were expressed in the first week of differentiation in the scaffold group but absent in control. All 4 markers detected by western blot were expressed earlier by 1 week in the scaffold group than those in the control group.
Cell survival and scaffold biocompatibility
Evaluation of biocompatibility results (showed in Figs. 4A–4D and 5) indicated that differentiated hepatocyte-like cells grew more stably (sustain proliferation) and their number was not significantly decreased in C-PLGA scaffolds after 3 weeks of differentiation (C-PLGA group, from 6 × 105 cells to 4.2 (±0.51) × 105 cells), unlike the control group (decreasing from 5 × 105 cells to 1.2 (±0.28) × 105 cells). Although there is a drop in cell numbers in the scaffold group in the first week, this phenomenon may be induced by the temporary microenvironment change of BMSCs being removed from the monolayer culture system to the 3D culture system. In contrast, the percentage of differentiated cells in monolayer culture system decreased gradually (cell confluence before and after differentiation in weeks 1, 2, and 3 was 90%, 85%, 60%, and 45%, respectively; data not shown) after 1 week of differentiation under phase-contrast microscopy (Fig. 4E–4H). Cell growth curves (Fig. 5) revealed that the densities of differentiated hepatocyte-like cells were significantly higher in the scaffold group compared with the control after 3 weeks of differentiation. Thus, the lifespan of hepatocyte-like cells differentiated in 3D scaffolds was prolonged when compared with those in monolayer culture.
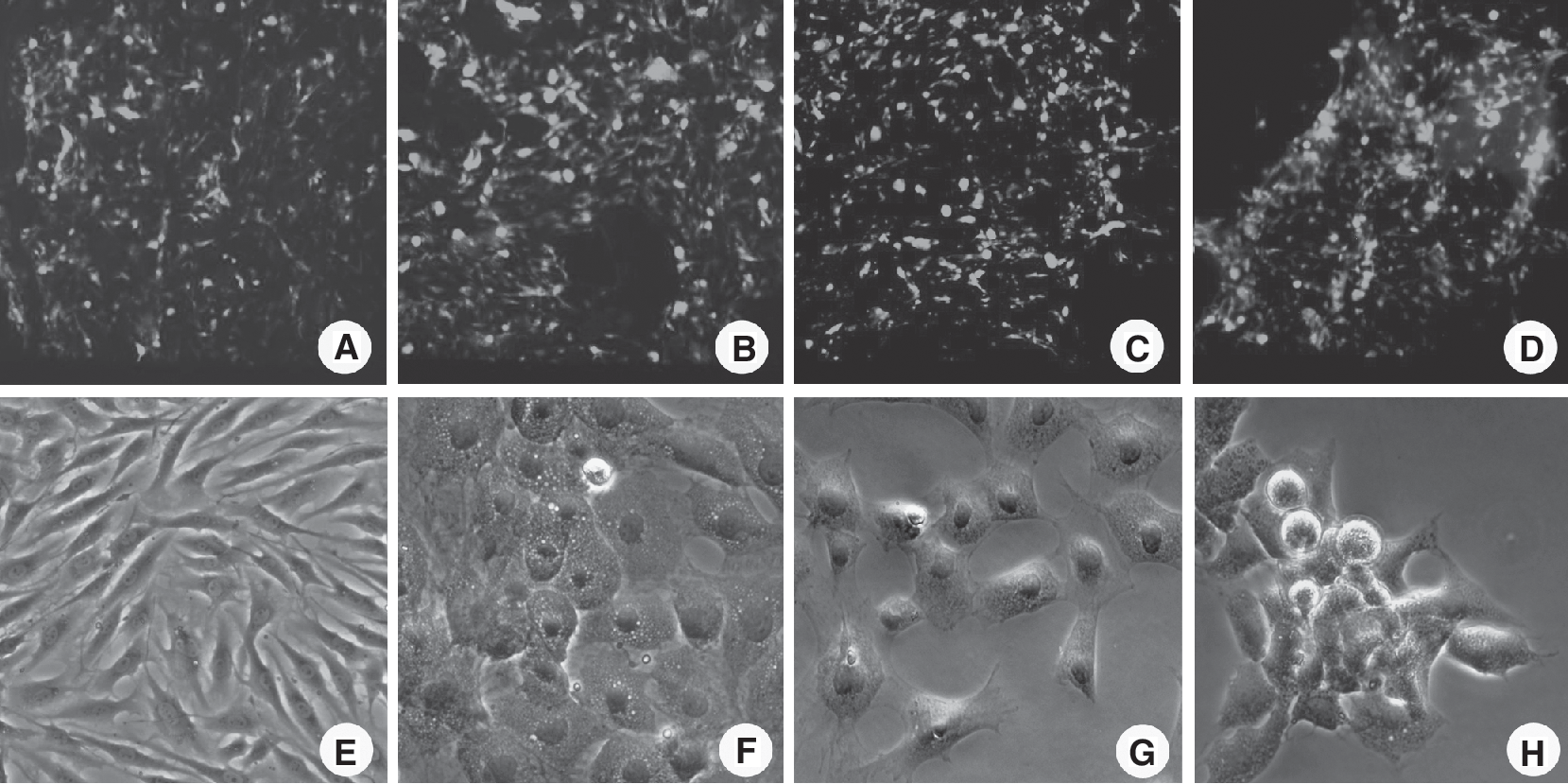
The morphology of bone marrow mesenchymal stem cell (BMSC)-derived hepatocyte-like cells in collagen-coated poly(lactic-co-glycolic acid) (C-PLGA) scaffolds under confocal laser scanning microscopy (CLSM) (

Cell growth curves showed that cell numbers underwent no significant decrease in the collagen-coated poly(lactic-co-glycolic acid) (C-PLGA) scaffold group but decreased quickly in the monolayer control group after 3 weeks of differentiation (the initial cell density was 6 × 105 cells per scaffold and 5 × 105 cells per T-25 culture flask).
Biochemical assay for hepatic metabolism functions
The results of ALB detection (Fig. 6A) showed that ALB concentration was 0.04 ± 0.02 pg/cell/h in the first week, increased to 0.14 ± 0.02 pg/cell/h in the second week, and rose to 0.58 ± 0.15 pg/cell/h in the third week in scaffold. In contrast, ALB was not detected in the monolayer culture group in the first week. In the next 2 weeks, ALB was 0.07 ± 0.2 pg/cell/h and 0.36 ± 0.11 pg/cell/h, respectively. Compared with control group, ALB secreted from differentiated hepatocyte-like cells in the scaffold group was significantly higher than that in control (P < 0.05). Urea synthesis assay (Fig. 6B) revealed that urea synthesis from differentiated hepatocyte-like cells in scaffold was 1.4 ± 0.6 pg/cell/h in the first week of differentiation, but was not detected in the control group at the same time. In the next 2 weeks, the urea concentration increased to 10.1 ± 3.5 pg/cell/h and 24.6 ± 3.0 pg/cell/h in scaffold group. At the same observation period, urea concentration was only 4.9 ± 1.6 pg/cell/h and 11.7 ± 3.8 pg/cell/h, respectively, in the control group. The capability to synthesize urea in differentiated hepatocyte-like cells in C-PLGA scaffold was significantly higher than that in the monolayer differentiation system (P < 0.05). Metabolism analysis of ammonia and diazepam (Fig. 6C and 6D) showed similar results in C-PLGA scaffold group and monolayer differentiation group. In the first week of differentiation in scaffolds, the capability to eliminate ammonia and clear diazepam was 0.08 ± 0.02 pg/cell/h and 0.08 ± 0.05 pg/cell/h, respectively, but neither was not detected at all in the control group. Ammonia elimination increased >2-fold (0.32 ± 0.06 pg/cell/h vs. 0.11 ± 0.02 pg/cell/h at week 2 and 1.19 ± 0.21 pg/cell/h vs. 0.42 ± 0.1 pg/cell/h at week 3) in the scaffold differentiation system compared to the control during the next 2-week observation period. Diazepam clearance in the scaffold group indicated the same increased trend (0.52 ± 0.08 pg/cell/h vs. 0.34 ± 0.03 pg/cell/h in the second week and 0.86 ± 0.11 pg/cell/h vs. 0.48 ± 0.08 pg/cell/h in the third week) compared to the monolayer control group during the same observation period. The results suggest that the metabolic function—including ALB secretion, urea synthesis, ammonia elimination, and diazepam clearance—from differentiated hepatocyte-like cells in C-PLGA scaffolds was significantly higher than that in the control (P < 0.05).
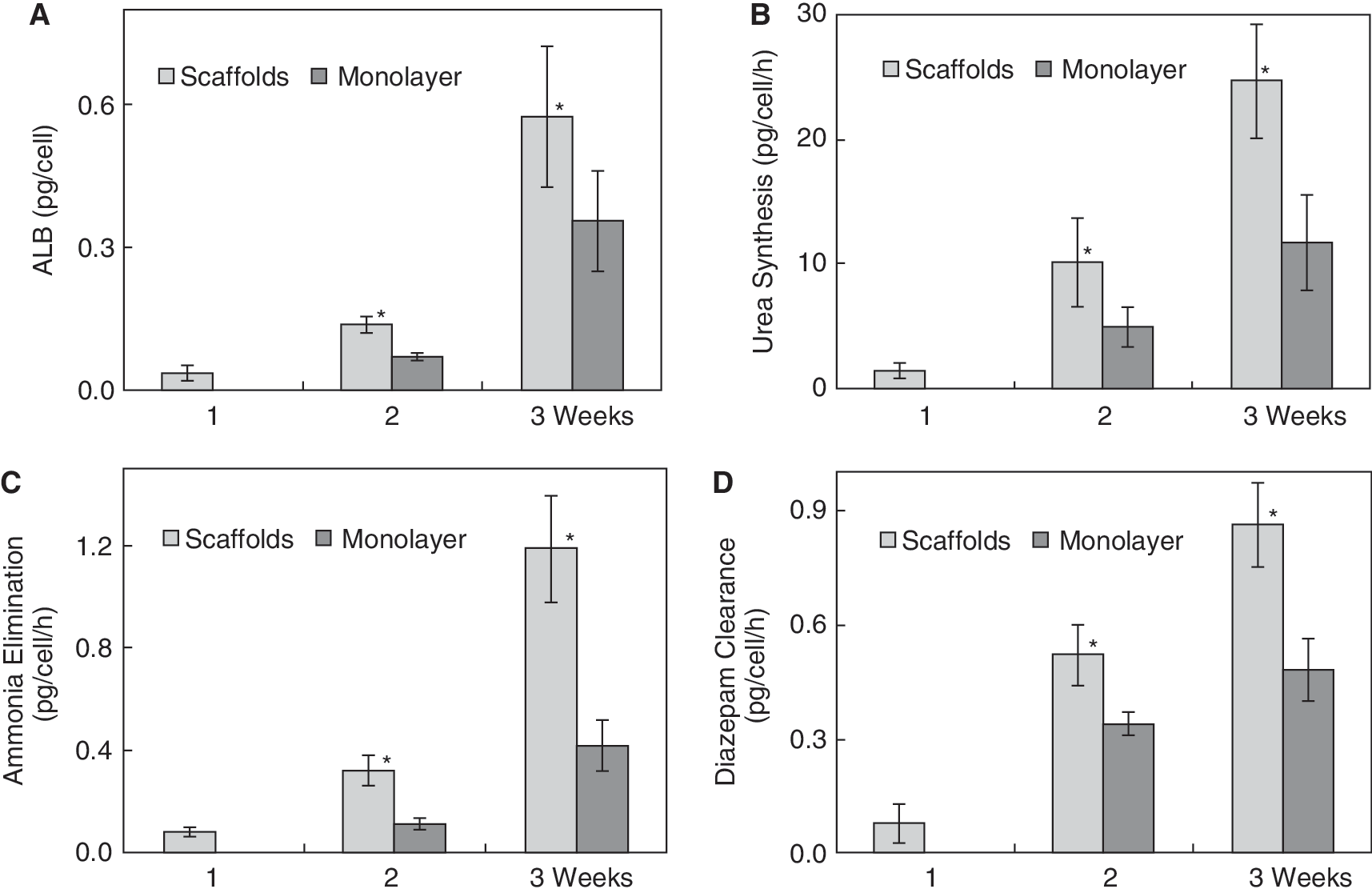
Results of biochemical assays. (
Histochemical and PAS staining
The result of H&E staining (Fig. 7) showed that differentiated BMSCs formed grape-shaped small cell clusters and the cells' morphology was changed from spindly to round shape in the second week of differentiation in C-PLGA scaffolds. A liver-like tissue was formed in the third week of differentiation. The result of PSA staining showed that glycogen was positive (cytoplasm stains pink) in the first week of differentiation. After 2 weeks of differentiation, the glycogen was expressed increasingly. The significantly increased expression of glycogen from differentiated BMSC-derived hepatocyte-like cells confirms the supporting roles of C-PLGA scaffolds.
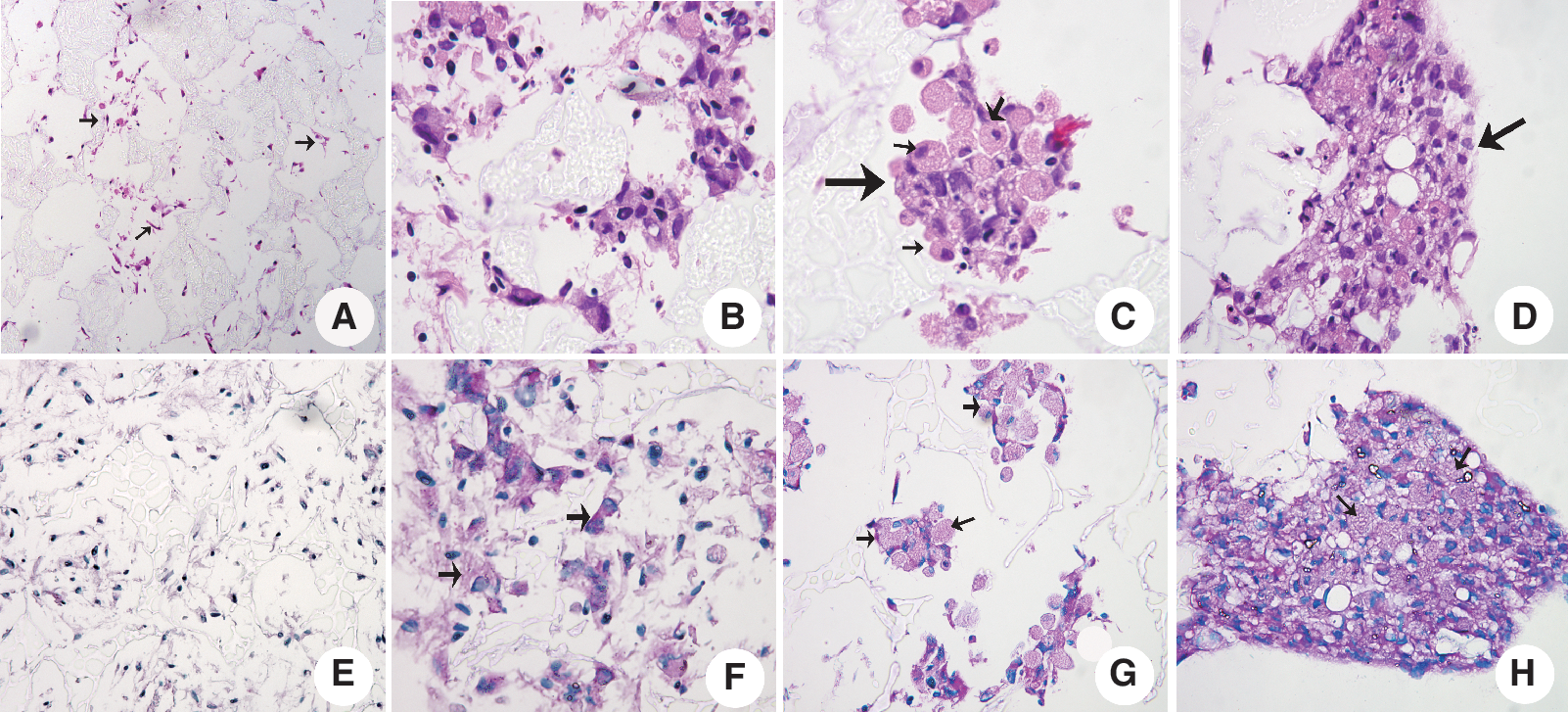
Results of H&E (
Discussion
Since the first demonstration of bone marrow stem cells differentiation into hepatocytes by Petersen et al. [32], the hepatogenic differentiation from BMSCs has been confirmed widely [32 –35]. This finding raised the possibility that these cells could be used for treatment of end-stage liver diseases due to various etiologies [36]. However, subsequent studies indicated that frequency of the transdifferentiation phenomenon and survival of implanted cells in vivo is very low [37,38]. Liver tissue engineering based on synthetic scaffolds provides a promising alternative to hepatocyte infusion into the portal vein or injection into the splenic pulp for treatment of end-stage hepatic failure [39,40]. As an important part of liver tissue engineering, scaffolds provide a place for cell attachment, increased surface area, and support for a large cell mass and specific structures [41 –43]. More importantly, the scaffold must be biocompatible, allowing the organ to grow inside and support itself over time. Our previous work [28] showed that a collagen-modified 3D PLGA scaffold was superior to a control without collagen modification and to a 2D culture system in supporting growth and metabolism of primary human hepatocytes. The cell number and lifespan of primary hepatocytes were significantly greater in collagen-coated PLGA scaffolds compared with those in a control PLGA scaffold without collagen modification. The capability of cultured hepatocytes to synthesize ALB and urea, and metabolize ammonia and diazepam in C-PLGA scaffold was significantly higher than that in control. Although different synthetic scaffolds have been fabricated and used for liver tissue engineering [44 –47], reports on the transdifferentiation of BMSCs into the hepatic lineage in a C-PLGA scaffold are infrequent. This work was motivated by the hypothesis that C-PLGA scaffolds can provide a good microenvironment for differentiation of BMSCs into hepatocyte-like cells in vitro.
As described in the methods section, following phenotype profiling of freshly isolated BMSCs derived from rat bone marrow, BMSCs were seeded into collagen-coated PLGA scaffolds. The phenotypic feature analysis demonstrated that BMSCs were positive for CD29 and CD90, but negative for CD45 and β2m. Previous reports showed that biomodified 3D scaffolds influenced cell behaviors. Our results on cells survival and scaffold biocompatibility revealed that the biomodified 3D structure could provide the hepatocyte-like cells with an excellent environment, supportive of cell viability and proliferation, as indicated by previous reports [13,18,21,48].
In addition, many hepatocyte-specific genes commonly used to characterize the profiling of stem cell-derived hepatocyte-like cells (such as ALB, AFP, CK18, CK19, G6PD, and HNF-4) were expressed in our C-PLGA scaffold group 1 week earlier than that in our control group. Western blot analysis confirmed a similar result, namely that differentiated hepatocyte-like cells expressed liver-enriched functional markers including ALB, P450, CK18, and CK19 earlier by 1 week in scaffold group than that in control. These results demonstrated an effective differentiation of hepatocyte-like cells in scaffolds. Furthermore, biochemical analysis (Fig. 6) validated that BMSC-derived hepatocyte-like cells expressed hepatic metabolic functions (ALB secretion, urea synthesis, ammonia elimination, and diazepam clearance) higher (P < 0.05) and earlier by 1 week in C-PLGA scaffold compared with control. The results of histochemistry revealed that differentiated BMSCs in scaffolds exhibit hepatocyte morphology and form liver-like tissue in the third week after differentiation. Glycogen was expressed increasingly from differentiated BMSCs in scaffolds during the period of 3-week differentiation. These results confirmed our previous knowledge that 3D scaffold architecture provides a fine microenvironment to support differentiation of BMSCs.
In summary, BMSC-derived hepatocyte-like cells on C-PLGA scaffold expressed hepatocyte-specific genes and markers (ALB, AFP, CK18, CK19, G6PD, HNF-4α, P450, and collagen)—at mRNA and protein levels—1 week earlier by than those in a monolayer culture system. Successful transdifferentiation rates of BMSCs to mature hepatocytes with significantly higher metabolic functions on C-PLGA scaffold as compared to the monolayer culture system demonstrated the superiority of the scaffold. Although many additional issues need to be addressed in future studies, based on the above analysis and characterization, our experiments indicate that engineered 3D C-PLGA scaffolds can provide a suitable environment for such a differentiation. This material therefore appears as a valuable candidate for use as a bioscaffold for liver tissue engineering in future clinical therapeutic applications.
Footnotes
Acknowledgments
This work was supported by National S&T Major Project (2008ZX10002-011, 2008ZX10002-005), National Key Scientific Research Program (2007CB947802), National Natural Science Foundation of China (30970747), Zhejiang Provincial Natural Science Foundation (Y2090010), and by the US Navy Bureau of Medicine and Surgery. A part of this work was presented in the 44th Annual Meeting of EASL (The European Association for the Study of the Liver) as a poster in April, 2009. (Poster ID: 848)
Author Disclosure Statement
The authors declare that no competing financial interests exist.