
Abstract
Human bone marrow stromal cells (BM-SCs) possess the potential to differentiate, self-renew, and produce diverse trophic/growth factors and are an excellent cell therapy tool for degenerative diseases. However, they exhibit different therapeutic efficacies, depending on the health status and age of the cell donor. Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disorder characterized by progressive motor neuron death in the central nervous system. In this study, we isolated BM-SCs from 11 ALS patients and characterized their potential secretory capacity of neurotrophic factors. We identified significant reductions in the expression of Oct-4 and Nanog, and in the trophic factors ANG, FGF-2, HGF, IGF-1, PIGF, SDF-1α, TGF-β, and VEGF, but not in BDNF or ECGF. Migration of ALS-SCs was reduced, although the cells expressed the same markers for human mesenchymal phenotypes. These data suggest that ALS-SCs have diminished capacity as trophic mediators and may have reduced beneficial effects in cell therapy.
Introduction
B
Amyotrophic lateral sclerosis (ALS) is a fatal neurodegenerative disorder characterized by progressive motor neuron death in the spinal cord, brainstem, and motor cortex [15]. The cause and pathogenesis of ALS are largely unknown. Recently, stem cell therapy using BM-MSCs was found to be a feasible and safe potential therapeutic strategy for ALS [16 –19]. However, despite these results, stem cell therapy in general is still disputed for either an ALS transgenic mouse model [20,21] or human trials [22]. Therefore, several issues, including the transplanted cell types, route of administration, optimal dosage, and optimal cell passage before transplant, must be studied to determine the maximal beneficial effects of cell therapy for ALS and other neurological disorders [23].
The use of autologous SCs is a promising avenue for clinical trials, but different efficacies have been reported, depending on the health status and age of the donor [24 –26]. This suggests that the potential of SCs as therapeutic stem cells needs to be evaluated. In this study, we assess stem cell features as trophic mediators, including production of trophic factors, and migration, for BM-SCs derived from ALS patients and normal controls.
Materials and Methods
Isolation of bone marrow stromal cells
BM-SCs that remained after a human trial were studied from 11 ALS patients. Two normal bone marrows were obtained from the femur of 2 persons who had the traffic accidents. All patients provided written informed consent and understood all issues involving the use of SCs for basic research. This study was approved by the Institutional Review Board for Human Investigation of Hanyang University Hospital. To obtain BM-SCs from ALS patients, mononuclear cells were isolated by bone marrow aspiration at the iliac crest, followed by density gradient (Histopaque, density 1.077 g/mL; Sigma-Aldrich, St Louis, MO) and washing twice with Dulbecco's modified Eagle's medium containing low-glucose (DMEM-LG; GIBCO BRL, Grand Island, NY). Cells at a density of 2 × 105 cells/cm2 were cultured in DMEM-LG with 10% fetal bovine serum (FBS; Hyclone, Waltham, MA) at 37°C in 5% CO2 for 3 days. After removing non-adherent cells, culture medium was changed twice per week. For passage, cells were detached with 0.25% trypsin/ethylene diamine tetraacetic acid for 3 min at 37°C, seeded at a density of 4 × 103 cells/cm2, and expanded up to 80%–90% confluence. Cell immunophenotype was confirmed by flow cytometry. Two normal BM-SCs were isolated by the same procedure.
Cell culture
Three normal human BM-SCs were purchased from Lonza (Walkersville, MD) and cultured according to the supplier's recommendations. Normal and ALS hBM-SCs were cultivated in hBM-SC growth medium (Lonza), containing mesenchymal cell growth supplement,
Grouping of bone marrow stromal cells
To account for age-related decline, BM-SCs were separated by donor age into midlife, 35 to 55 years, or late-life, 60–90 years [27]. Five ALS patients were midlife, and 6 were late-life. Five normal hBM-SCs were from midlife donors.
Fluorescence-activated cell sorting analysis
To determine the mesenchymal phenotype [28] of normal-and ALS-SCs, monoclonal antibodies detected HLA-DR, CD49c, CD73, and CD105 antigens (Abcam, Cambridge, MA), CD34 antigen (Santa Cruz Biotech, Santa Cruz, CA), and CD45, CD29, and CD44 antigens (Sigma-Aldrich, St. Louis, MO). In brief, SCs at fourth passage were stained in phosphate-buffered saline (PBS) (Ca2+- and Mg2+-free) supplemented with 5% FBS (Hyclone). After the final wash, cells were fixed in 1% paraformaldehyde prior to fluorescence-activated cell sorting (FACS) analysis by FACscan (Becton, Dickinson and Company, Franklin Lakes, NJ) using fluorescein isothiocyanate (FITC)- or phycoerythrin (PE)-goat-antimouse immunoglobulin as the isotype control. To eliminate non-specific binding, the same fluorochrome/protein ratio was used for isotype control and antibody specificity.
Real-time reverse transcription-polymerase chain reaction
To quantify gene expression, fourth-passage normal-and ALS-SCs were cultivated under identical conditions and harvested at 80% confluence (usually after 2 days). Total RNA was extracted using Trizol reagent following the manufacturer's instructions (Invitrogen, Carlsbad, CA), and 3–5 μg reverse-transcribed using RevertAid™ M-MuLV reverse transcriptase (MBI Fermentas, Burlington, Ontario, Canada), with 0.2 μg random primer (Invitrogen), 1 mM dNTPs (Invitrogen), and the supplied buffer. First-strand cDNA was amplified using Power SYBR Green PCR master mix (Abioscience, Hatfield, AL10 9NA, UK) with primers for human Nanog, Oct-4, Nestin-1, ANG, BDNF, ECGF1, bFGF-2, HGF, IGF-1, PIGF, SDF-1α, TGF-1β, VEGF, and β-actin (Table 1).Real-time reverse transcription-polymerase chain reaction (RT-PCR) was 95°C for 10 min; and 40 cycles of 15 s at 95°C, 1 min at 60°C. After amplification, a dissociation curve was constructed from 60°C to 90°C. Ct values were converted to absolute amounts of cDNA (E-Ct). To correct for differences in cDNA amounts between samples, the target PCR was normalized to the geometric mean value for the reference gene β-actin. Primers were from GenoTech (GenoTech, Daejeon, Korea; Table 1), except for SDF-1α, VEGF, and β-actin from SuperArray (SuperArray Bioscience Corporation, Frederick, MD). Data shown are from 3 experiments. Statistical comparisons between groups were performed using the Mann–Whitney U-test.
Acc. No. indicates gene access number.
Enzyme-linked immunosorbent assay
Fourth-passage BM-SCs were plated at 2 × 105 per well in 12-well plates in serum-free medium. After 48 h, each culture supernatant was divided into 200-μL triplicate samples, and trophic factors were measured via ANG, bFGF-2, IGF-1, VEGF ELISA kits (R&D Systems, MN) following the manufacturer's instructions. Data presented are from 3 experiments. Statistical comparisons between groups were conducted using the Mann–Whitney U-test.
Cell migration assay
Cell migration was examined by a QCM chemotaxis (8-μm pore size) 24-well migration assay (Chemicon, Temecula, CA) according to the manufacturer's instructions. In brief, 0.3 × 105 fourth-passage normal- or ALS-SCs in 300 μL serum-free medium were seeded in the migration chamber with 500 μL serum-free medium in the lower chamber. Plates were incubated at 37°C in 5% CO2 for 24 h, Suspended cells were gently removed by pouring, and the chambers were rinsed in water several times. Cells adhering to the top of the membrane were removed with a cotton applicator, and the clean chamber plate was placed onto a new 24-well feeder tray containing 400 μL prewarmed cell stain solution. After 20 min at room temperature, the chambers were rinsed in water several times, and 200 μL lysis buffer/dye solution was added to the feeder tray and incubated 15 min at room temperature. The mixture (100 μL) was transferred to a new 96-well plate and read with a fluorescence plate reader using a 480/520-nm filter set. The data presented are from 3 experiments. Statistical comparisons between groups were conducted using the Mann–Whitney U-test.
Results
Characteristics of primary human normal- and ALS-SCs
To analyze the mesenchymal phenotype of human BM-SCs from ALS patients, we used 5 normal human BM-SC samples (normal-SCs) as a control and 11 human BM-SC samples isolated from ALS patients (ALS-SCs). ALS-SCs had a flattened and spindle-shaped appearance, with morphology similar to normal-SCs. Flow cytometric analysis of both types of subcultured SCs revealed a CD29+ (Integrin β1), CD44+, CD49C+ (Integrin α3), CD73+ (5′-nucleotidase), CD105+ (endoglin), CD34− (hematopoietic marker), CD45− (leukocyte antigen), HLA-DR− (human leukocyte antigen DR) phenotype [28]. The characteristics of the normal- and ALS-SC groups are in Table 2.
M indicates male, F indicates female.
Abbreviations: ALS, amyotrophic lateral sclerosis; SC, stromal cell.
Reduced expression of Oct-4 and Nanog in ALS-SCs
To determine the plasticity of the isolated SCs, we performed quantitative real-time RT-PCR with primers (Table 1)for Oct-4 [1], Nanog [2], and Nestin [29]. Oct-4 and Nanog expression were significantly decreased in all ALS-SCs, while Nestin did not change, in compared to normal-SCs (Fig. 1, open bar). To examine age-related decline, ALS-SCs were analyzed in midlife and late-life groups (see Material and Methods) [27]. The expression of Oct-4 and Nanog significantly decreased in midlife ALS-SCs, while Nestin did not change in ALS-SCs relative to normal-SCs (Fig. 1, filled bar). No significant differences were observed between the midlife and late-life groups in ALS-SCs (Fig. 1, gray bar).
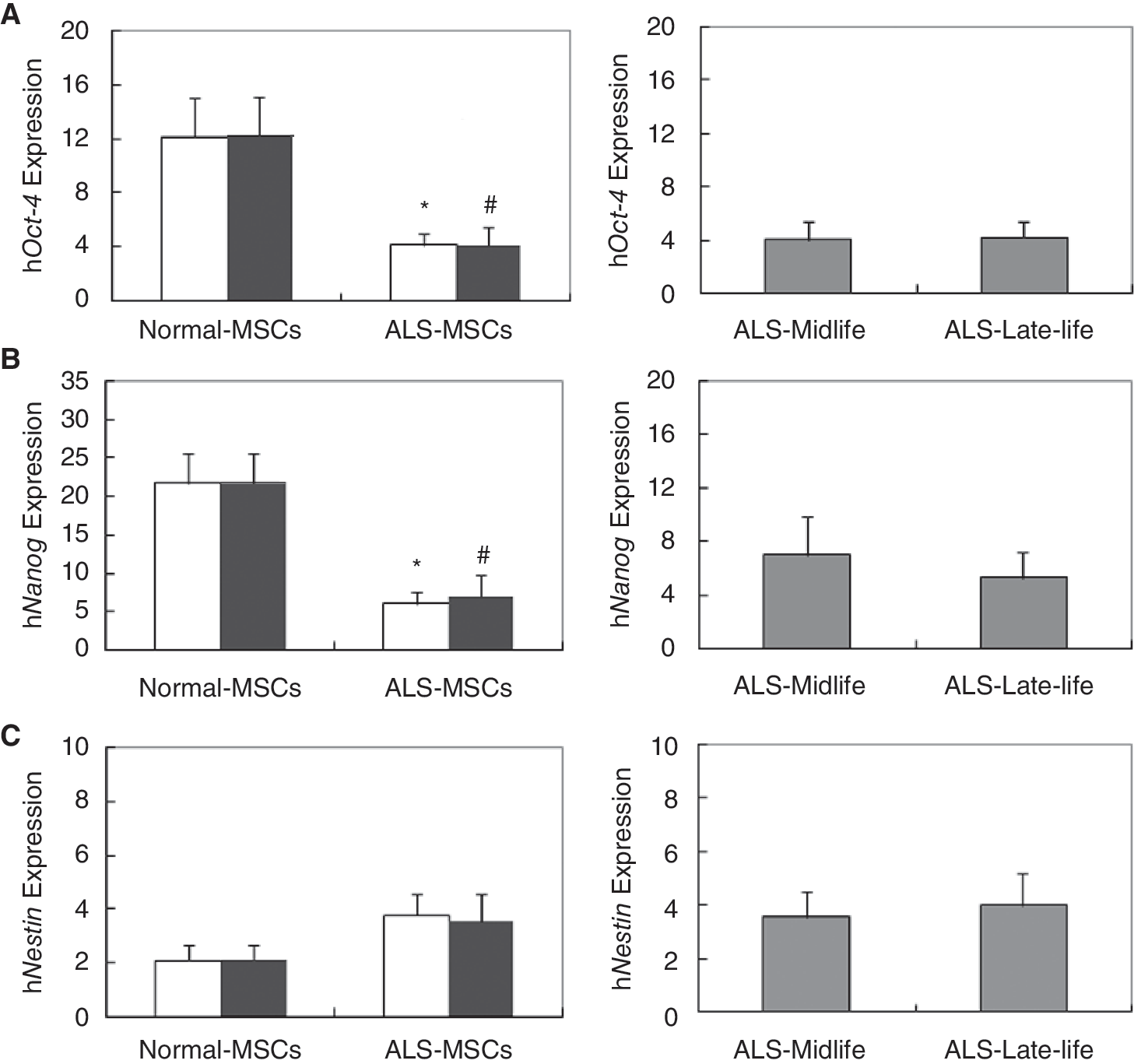
Reduced expression of Oct-4 and Nanog in amyotrophic lateral sclerosis-stromal cells (ALS-SCs). By real-time RT-PCR, levels of Oct-4 (
Differential expression of trophic factors between ALS- and normal-SCs
The ability to release trophic factors is an important characteristic of stem cells [6]. Quantitative real-time RT-PCRs with primers (Table 1)for various trophic factors identified a significant decrease in expression of ANG, bFGF-2, HGF, IGF-1, PIGF, SDF-1α, TGF-β, and VEGF in ALS-SCs, as compared to normal-SCs (Fig. 2, open bar). BDNF and ECGF did not change, according to the Mann–Whitney U-test (Fig. 2, open bar). The expression patterns were the same for the midlife groups of both MSC types (Fig. 2, filled bar), and no differences were observed between midlife and late-life groups in ALS-SCs (data not presented).

Differential gene expression of trophic factors in amyotrophic lateral sclerosis-stromal cells (ALS-SCs). Measured by real-time RT-PCR, expression levels of ANG (
To confirm differential expression at the protein level, ELISA analysis was performed on supernatants obtained for each cultured BM-SCs. As shown in Figure 3, ANG, bFGF-2, IGF-1, and VEGF were significantly decreased in ALS-SCs (Fig. 3, open bar). These patterns were the same for the mid-life groups for both SC types (Fig. 3, filled bar). Consistent with the data in Figure 2, midlife and late-life groups in ALS-MSCs were not significantly different (data not presented). This suggests that the capabilities as trophic mediators are severely reduced in ALS-SCs.

Reduced secretion of trophic factors by amyotrophic lateral sclerosis-stromal cells (ALS-SCs). The secretion of trophic growth factors from normal- and ALS-SCs was measured by enzyme-linked immunosorbent assay (ELISA). The levels of ANG (
ALS-SCs lacked migration capacity
Since ALS-SCs appear to have reduced capability and seem to release trophic factors, we hypothesized that their ability to migrate might be compromised. To test this hypothesis, we examined motility using a chemotaxis migration assay with 8-μm pore size, 24-well plates. With normal-SC migration set to 100%, ALS-SCs showed significantly decreased migration, at 63% for total group (Fig. 4A, open bar) and 58% for the midlife group (Fig. 4A, filled bar), suggesting that the decline in ALS-SCs correlated with cell migration. Comparison of midlife and late-life groups in ALS-SCs showed no differences (Fig. 4B, gray bar).
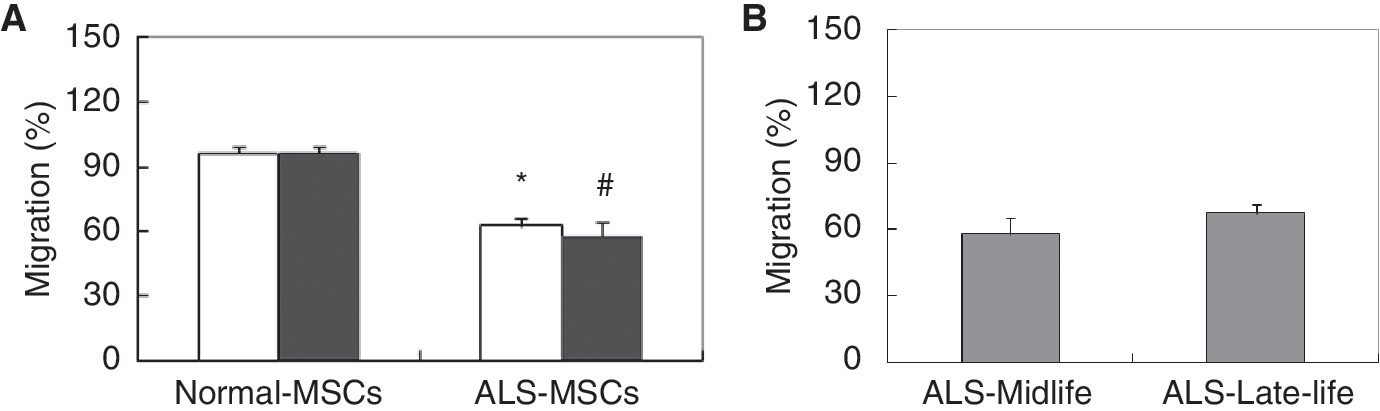
Amyotrophic lateral sclerosis-stromal cells (ALS-SCs) lacked migratory capacity. SCs were allowed to migrate for 24 h in Boyden chambers. After removal, migrating SCs were read on a plate reader using a 480/520 nM filter set. (
Discussion
We have isolated and characterized human SCs from the bone marrow of ALS patients. Eleven ALS-SC samples and 5 normal-SC samples were analyzed. Oct-4 and Nanog (Fig. 1), production of various trophic factors (Figs. 2 and 3), and migration ability (Fig. 4) were significantly reduced in ALS-SCs, although the cells expressed the same markers for mesenchymal phenotypes (Table 1), suggesting that ALS-SCs have reduced capacity as trophic mediators and may have reduced beneficial effects in autologous cell therapy.
MSCs have a substantial capacity for self-renewal and multipotency, as evidenced by their continuous expression of Oct-4 (Octamer-4), Nanog, and Sox-2 [1,2]. Here, we found that SCs from ALS patients were significantly reduced in their expression of Oct-4 and Nanog but not Nestin (Fig. 1). This may suggest that ALS-SCs lose plasticity, an important feature of stem cells.
MSCs are trophic mediators [6], and in vitro studies demonstrated that MSCs secrete multiple anti-inflammatory, angiogenic, neurotrophic, immunomodulatory, and antifibrotic factors [7 –10]. Preclinical studies with MSCs demonstrated functional recovery in a variety of injured tissues without significant differentiation of injected cells [30]. Thus, the release of pro-regenerative factors from injected MSCs, as well as the injected MSCs themselves, induces the protection and recovery of surrounding tissue and cells, which is an important beneficial effect in cell therapy [6]. In this study, we analyzed whether ALS-SCs are adequate trophic mediators. We quantified the secretion of trophic factors by quantitative PCR and ELISA, and found that BM-SCs isolated from ALS patients have diminished production of diverse trophic factors including ANG, FGF-2, HGF, IGF-1, PIGF, SDF-1α, TGF-β, and VEGF, but not BDNF and ECGF, as compared to normal controls (Figs. 2 and 3).
The trophic factors tested in this study are also angiogenic factors. Their transcription increases under hypoxic conditions [31,32], and they stimulate several biological functions, including protection and migration [33]. We found that migratory potential is decreased in all ALS-SCs, as compared to normal controls (Fig. 4). Previous studies showed that endogenous progenitor cells obtained from patients with diabetes or heart failure had profoundly reduced functional capacity, such as for neovascularization, migration, and adhesion [26,34,35]. The migratory activity of circulating endothelial progenitor cells inversely correlates with risk factors for coronary artery disease [36] and chronic ischemic heart disease [26]. These data support our findings that suggest reduced efficacy of ALS-SCs in stem cell therapy.
The number and potential for differentiation of human BM-MSCs significantly decline with age [37 –39]. Caplan used estimates of the marrow titers of human MSCs, measured as colony-forming units-fibroblastic (CFU-f), to identify a decline in the number and potential of MSCs [3]. He divided MSCs into 6 groups based on capacity: embryology, neonatology, teens, peak skeletal performance, midlife, and late-life [3,27]. In this study, we divided MSCs into mid-life and late-life groups to eliminate the parameter of age-related decline and found no statistical difference between midlife and late-life ALS-SCs for all characteristics tested. However, this result may be the effect of the small sample size of this study.
Cells used in this study were isolated from bone marrow. Recent studies have provided the evidences that there are non-mesenchymal pluripotent stem cells located within the bone marrow. These pluripotent stem cells are blastomere-like stem cells (BLSCs) [40], very small embryonic-like (VSEL) stem cells [41] and epiblastic-like stem cells (ELSCs) [42]. Therefore, the cells isolated from bone marrow are regarded as BM-SCs.
Autologous stem cell therapy may increase safety and alleviate ethical issues. However, if MSCs obtained from patients are not healthy, autologous cells offer reduced benefits [3, 6]. This may be the case for/in ALS patients, which means adequate healthy or alternative sources of MSCs for autologous and allogeneic use must be investigated.
In conclusion, we isolated BM-SCs from ALS patients and determined their trophic capacity as trophic mediators. We identified a decline in expression of Oct-4 and Nanog, and several trophic growth factors in ALS-SCs. Migratory effects were also reduced in ALS-SC versus normal controls. Our data that BM-SCs isolated from ALS patients have reduced potential as trophic mediators suggest that they may have reduced beneficial effects in compared to healthy BM-SCs applied to same conditions in cell therapy. Therefore, the natural deficit of ALS-SCs may be overcome by increasing the number of ALS-SCs in clinical trial of autologous stem cell therapy.
Footnotes
Acknowledgment
This work was supported by the cluster research fund of Hanyang University (HY-2008-C).
Author Disclosure Statement
No competing financial interests exist.