
Abstract
Mesenchymal stem/stromal cell-like populations derived from adult bone marrow (BMSC), dental pulp (DPSC), and periodontal ligament (PDLSC) have the ability to differentiate into cells of mesenchymal and non-mesenchymal tissues in vitro and in vivo. However, culture-expanded MSC-like populations are a heterogeneous mix of stem/committed progenitor cells that exhibit altered growth and developmental potentials. In the present study we isolated and characterized clonal populations of BMSCs, DPSCs, and PDLSCs to identify potential biomarkers associated with long-lived multipotential stem cells. Microarray analysis was used to compare the global gene expression profiles of high growth/multipotential clones with low growth potential cell clones derived from 3 stromal tissues. Cross-comparison analyses of genes expressed by high growth/multipotential clones derived from bone marrow, dental pulp, and periodontal ligament identified 24 genes that are differentially up-regulated in all tissues. Notably, the transcription factors, E2F2, PTTG1, TWIST-1, and transcriptional cofactor, LDB2, each with critical roles in cell growth and survival, were highly expressed in all stem cell populations examined. These findings provide a model system for identifying a common molecular fingerprint associated with immature mesenchymal stem-like cells from different organs and implicate a potential role for these genes in MSC growth and development.
Introduction
C
Recent studies have identified clonogenic MSC-like populations in dental structures, including adult dental pulp, periodontal ligament, and the pulp of exfoliated deciduous teeth [7 –9] with phenotypic/genotypic characteristics and properties similar to those described for BMSC [10,11]. Clonogenic MSC-like cells from bone marrow, dental pulp, and periodontal ligament are all capable of forming mineralized nodules and adipocytes under defined inductive conditions in vitro. In contrast, human dental pulp stem cells (DPSCs) do not form bone and hematopoietic supportive stroma in vivo, but rather exhibit the capacity to regenerate an ectopic dentin-pulp-like microenvironment following transplantation into immunocompromised mice [7]. Similarly, when transplanted under the same conditions, human periodontal ligament stem cells (PDLSCs) are capable of regenerating calcified structures of cementum impregnated with periodontal ligament-like filaments (Sharpe's fibers) that sprout from the surrounding collagen-rich fibrous tissue [8]. Collectively, these studies infer that adult-derived MSC-like cells are genetically programmed to regenerate the microenvironments from which they were originally derived in vivo. The common stem cell-like properties and heterogeneity exhibited within the different BMSC, DPSC, and PDLSC populations provide a model system for comparative genomic profiling as a means of identifying gene expression patterns that are associated with the growth and development of stromal stem cells and their more committed progeny.
The emergence of high-density gene array technology, in combination with the knowledge obtained from the sequencing of the human genome [12,13], allows the assessment of global patterns of gene expression across multiple experimental samples and enables identification of major genomic differences and unique biological markers specific to the target cell populations [14]. These advances in genomic profiling provide rapid and reliable data sets that define vital processes involved in cellular growth, survival, and development that can be readily applied to different MSC-like populations following immunoselection and ex vivo expansion. The aim of our study was to identify differentially expressed genes unique to immature, high-proliferating, multipotential MSC-like populations that are likely to be used in the treatment of various clinical conditions. To achieve this, we compared the growth and differentiation of stem/progenitor cells and their progeny contained within the BMSC, DPSC, and PDLSC populations using a global microarray analysis strategy. Using this approach, we have identified a common gene set commonly expressed by MSC clones from bone marrow, dental pulp, and periodontal ligament that exhibit high growth capacity and multilineage differentiation potential.
Materials and Methods
Subjects
Human bone marrow aspirates were obtained from the posterior iliac crest of normal adult volunteers (20–35 years old) following informed consent, according to procedures approved by the ethics committee of the Royal Adelaide Hospital, South Australia. Bone marrow mononuclear cells (BMMNCs) were prepared by density centrifugation as previously described [6, 15]. Normal human-impacted third molars were collected from healthy volunteers between 15 and 19 years of age with informed consent, according to procedures approved by the ethics committee of the University of Adelaide, South Australia. Dental pulp and periodontal ligament tissue were isolated and enzymatically digested to generate single cell suspensions as previously described [7,8,16].
Cell isolation and culture
Magnetic-activated cell sorting was performed on BMMNC preparations as previously described [6,15]. In brief, 1−3 × 108 BMMNCs were sequentially incubated with mouse anti-human STRO-1 [17] on ice for 1 h. Following washing, the cells were incubated with goat anti-mouse IgM-biotin (Southern Biotechnology Associates, Birmingham, AL) for 30 min on ice, washed, and then incubated with anti-biotin microbeads (Miltenyi Biotec Inc., Auburn, CA) for 15 min on ice. STRO-1-positive cells were then separated on a Mini MACS magnetic column (Miltenyi Biotec Inc.) according to the manufacturer's recommendations.
Dynal bead cell sorting was performed on single cell suspensions of dental pulp and periodontal ligament as previously described [15]. In brief, each cell suspension was incubated with mouse anti-human STRO-1 for 1 h on ice. Following washing, the cells were incubated with rat anti-mouse IgM-conjugated Dynabeads (Dynal Biotech, Oslo, Norway) at 4 beads per cell on a rotary mixer at 4°C for 2 h. STRO-1-positive cells were isolated with Dynal MPC-1 magnetic particle concentrator (Dynal Biotech) according to the manufacturer's recommendations.
Primary BMSC, DPSC, and PDLSC cultures were established in α-minimum essential media (α-MEM; Sigma-Aldrich, St. Louis, MO) supplemented with 20% fetal calf serum (FCS; Thermo Electron, Melbourne, Australia) and 100 μM
Assessment of population doublings
Single cell-derived colonies obtained from primary cultures of BMSC, DPSC, and PDLSC populations were seeded at 8 × 103 cells/cm2 and allowed to expand to ∼90% confluency before being harvested by trypsin/EDTA (JRH Biosciences, Lenexa, KS) treatment. Representative cell samples for each clone were passaged at a seeding density of 8 × 103 cells/cm2 into 24-well plates until they had reached in vitro cellular senescence. Cell counts were performed at each passage and population doublings were calculated using the formula (log2 final cell number/log2 seeding cell number). The final population doubling value for each colony was represented as the sum of population doubling values obtained at each passage. The population doubling times per cell passage were calculated using the formula: Generation time = (log2 × days in culture)/(log final cell number—log seeding cell number).
Differentiation assays
Mineralization was induced as previously described [6,18]. In brief, BMSCs, DPSCs, and PDLSCs at passage 6 were seeded at 8 × 103 per cm2 in 24-well plates and cells were cultured in α-MEM supplemented with 5% FCS, 100 μM
Gene expression profiling (microarray analysis)
Total cellular RNA from 5 × 105 cells of 3 highly proliferative/bipotential clones and 3 less proliferative clones of BMSCs, DPSCs, and PDLSCs at passage 6 was isolated using phenol–chloroform (TRIzol; Invitrogen, Carlsbad, CA) extraction and purified using column chromatography (RNeasy® MinElute Cleanup Kit; Qiagen, Doncaster, Victoria, Australia) according to the manufacturer's instructions. Microarray analysis was performed on a commercial Illumina WG-6 platform, containing bead chips targeting 48,000 human transcripts. Samples of 1 pg of total RNA at a concentration of 100 ng/μL were provided for labeling, hybridization, scanning, and analysis that were performed by Dr. Stephen Wilcox, at the Australian Genome Research Facility (AGRF), VIC, Australia. The results obtained were normalized by quantile normalization method using lumi package in R statistical software and were subsequently imported into GeneSpring GX for further statistical analysis.
Real-time PCR analysis
Total RNA isolated from subpopulations submitted for microarray analysis (5 × 105 cells of 3 high proliferative/multipotential clones and 3 low proliferative clones of BMSCs, DPSCs, and PDLSCs at passage 6) was used as a template for cDNA synthesis and the expression of 24 transcripts (primer sets outlined in Table 1)was assessed by polymerase chain reaction (PCR) amplification as previously described [22]. In order to identify the differences present in cells of varying differentiation and proliferation potential, comparative genomic analysis was performed on 3 biological replicates of cell clones exhibiting low proliferation potential and high proliferation/differentiation potentials, for each tissue type.
Results
Differential growth potential of BMSC, DPSC, and PDLSC clones in vitro
To determine the proportion of high growth/multipotential clones within each of the cell populations, individual plastic-adherent, MSC-like colonies derived from human bone marrow, dental pulp, and periodontal ligament were isolated using cloning rings as previously described [6,11]. A total of 260 colonies obtained from 9 BM donors were expanded in continuous culture until cellular senescence was achieved. A small proportion of clones (31/260; 12%) exhibited high growth potential that exceeded 20 population doublings (mean generation time of 0.48 PD per day ± 0.05 SD). However, the majority of clones (179/260; 69%) exhibited a limited to moderate proliferation potential and were unable to expand beyond 20 population doublings with a mean generation time of 0.29 PD per day ± 0.03 SD (Fig. 1).
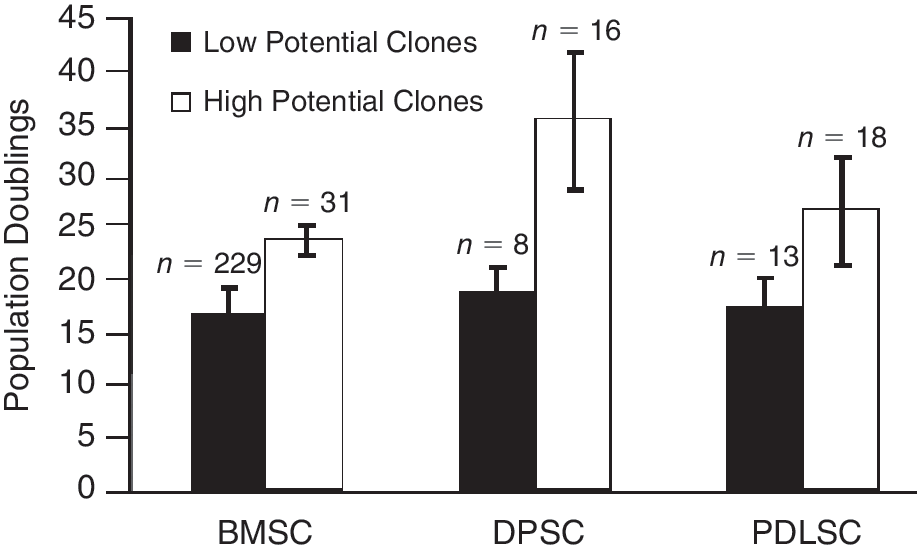
Differential growth potential of mesenchymal stem/stromal cell (MSC)-like clonal populations. Individual clonal cell populations of adult bone marrow stem cell (BMSC), dental pulp stem cell (DPSC), and periodontal ligament stem cell (PDLSC) exhibit varying growth potentials as determined by the total number of population doublings for each clone. The BMSC population contained a smaller proportion of long-lived clones in comparison with DPSC and PDLSC populations.
Similar studies were performed for clonogenic DPSC and PDLSC, previously reported to have higher proliferative potentials and life spans compared with their bone marrow counterparts [8,10]. A total of 24 clones obtained from 4 dental pulp donors were serially passaged until they had reached cellular senescence as described earlier. The majority of clones (16/24; 67%) exhibited high growth potential, ranging from 25 to 45 population doublings (mean generation time 1.07 PD per day ± 0.25 SD). However, the remainder of the colonies (8/24; 33%) were found to possess a low growth potential with a mean generation time of 0.29 PD per day ± 0.01 standard deviation (SD) (Fig. 1). Similarly, a total of 31 clones obtained from 3 periodontal ligament donors were expanded in culture to the point of senescence. Approximately 58% (18/31) of the clones exhibited high levels of growth ranging from 21 to 37 population doublings (mean generation time 0.69 PD per day ± 0.13 SD), while the remaining clones demonstrated low growth potential (13/31; 42%) with a mean generation time of 0.30 PD per day ± 0.01 SD (Fig. 1).
Differential potential of long-lived BMSC, DPSC, and PDLSC clones in vitro
BMSC, DPSC, and PDLSC clones that exhibited high levels of growth were subsequently assessed for their potential to differentiate into cells of osteogenic, adipogenic, and chondrogenic lineages. After 4 weeks of osteoinduction, all of the BMSC clones assessed (31/31, 100%) produced Alizarin Red staining mineralized nodules [6, 18]. Furthermore, 90% of the clones (28/31) also exhibited adipogenic differentiation potential, indicated by the formation of Oil Red O-positive lipid clusters (Fig. 2).
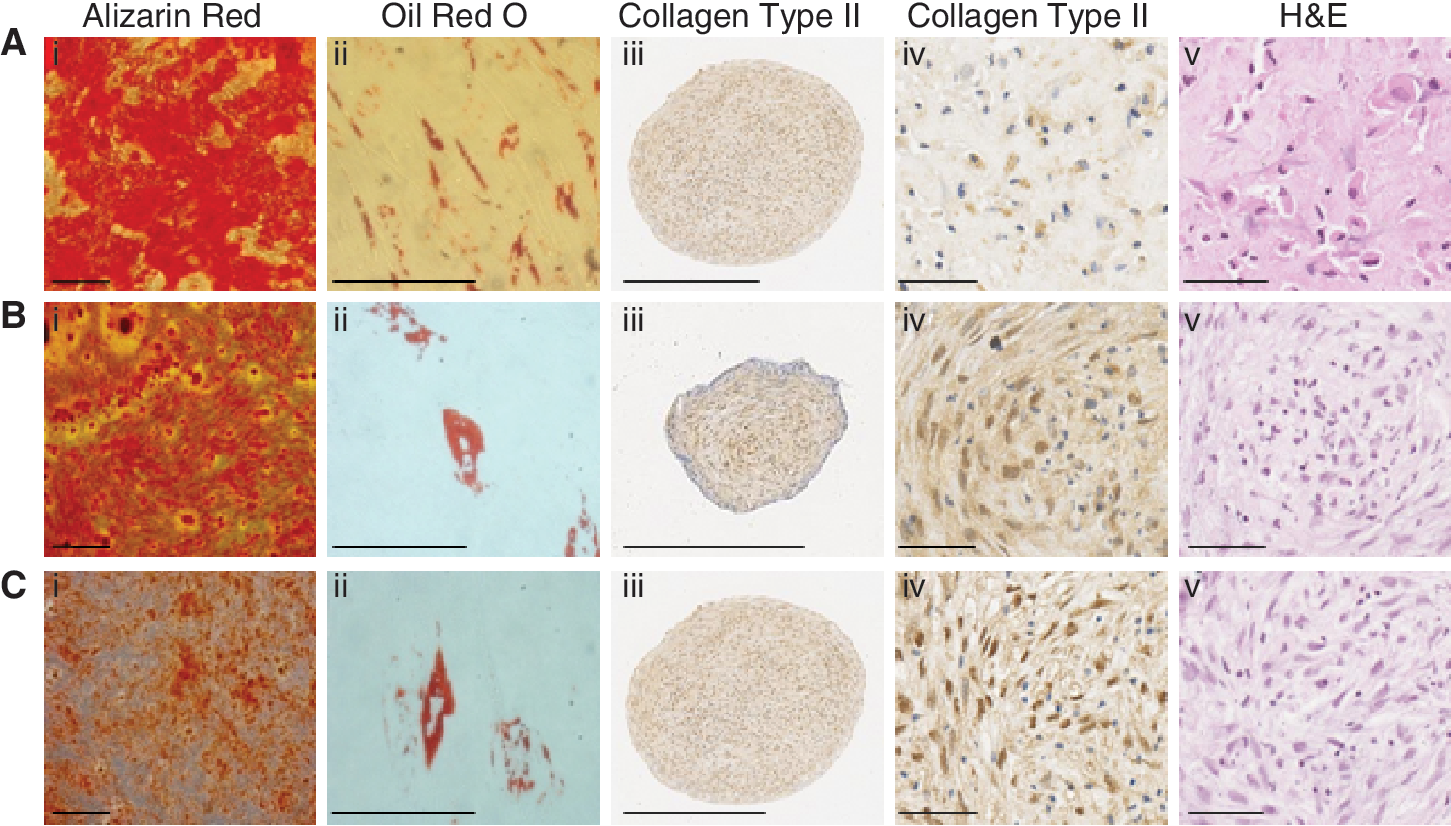
Developmental potential of long-lived mesenchymal stem/stromal cell (MSC)-like populations. Individual clonal cell populations of long-lived adult bone marrow stem cell (BMSC), dental pulp stem cell (DPSC), and periodontal ligament stem cell (PDLSC) exhibit varying levels of mineralization, adipogenic, and chondrogenic differentiation potential. (i) Alizarin Red (scale bar = 200 μm) and (ii) Oil Red O (scale bar = 50 μm) staining of clonal (
In parallel studies, 16 high proliferative DPSC clones exhibited the capacity to form Alizarin Red-positive mineralized nodules, while 31% (5/16) also displayed the ability to form Oil Red O-positive lipid-laden globules (Fig. 2). All of the PDLSC clones (18/18; 100%) demonstrated potential to form mineralized deposits, while 37% (7/18) of these clonal lines also formed adipocytes (Fig. 2).
Upon identification of their differentiation potential into cells of osteogenic and adipogenic lineages, 3 bipotential clones were selected from each of the 3 tissue types and assessed for their chondrogenic differentiation potential. The selected cell clones showed capacity to differentiate into cells of the chondrogenic lineage, indicated by immunohis-tochemical staining of cell pellets with anti-collagen type II antibody (Fig. 2). Quantitative analyses demonstrated similar multipotentials between different MSC clones for each tissue type (Supplementary Fig. 1; Supplementary materials are available online at
Genomic profiling of BMSC, DPSC, and PDLSC clonal lines exhibiting differential growth and developmental potentials
To identify genes involved in MSC maintenance, growth, and development, we used large-scale gene expression profiling of different clonal lines of varying differentiation and proliferation potentials. Comparative genomic analysis was performed using a commercial Illumina WG-6 platform on 3 biological replicates of clonal cell lines exhibiting low growth potential compared with 3 clones exhibiting high growth/multidifferentiation potentials, for BMSC, DPSC, and PDLSC populations (Fig. 3). The microarray analyses were based on 3 biological replicate clones for each tissue that displayed similar differentiation potentials as shown in Figure 3.

Hierarchical sample clustering of BMSC, dental pulp stem cell (DPSC), and periodontal ligament stem cell (PDLSC) clones. Representative dendrograms illustrate patterns of gene expression levels, of significant differentially expressed genes within the samples analyzed for (
Lumi package in R statistical software and GeneSpring GX software were used for statistical analysis, where a cutoff point with a P value of 0.05 was used to identify statistically significant differentially expressed genes with greater than 2-fold differences in expression. These bioinformatic analyses identified 968, 603, and 842 differentially expressed genes between low growing clones and high growth/multidifferentiation potential clones for BMSC, DPSC, and PDLSC, respectively (Fig. 3).
Identification of common gene sets expressed by long-lived, multipotent BMSC, DPSC, and PDLSC clonal lines
Further cross-comparison analyses were performed to identify differentially expressed genes that were commonly expressed amongst long-lived, multipotent cell populations derived from the 3 different stromal tissues. Interrogation of the data resulted in the identification of 24 common differentially up-regulated genes that could be grouped into categories based on their involvement in different cellular processes, including cell cycle, mitosis, and cell division, as well as DNA repair and replication and cellular differentiation (Table 2).Within this gene data set, the transcription factors, E2F2, PTTG1, TWIST-1, and transcriptional cofactor, LDB2, were found to be commonly up-regulated in long-lived, multipotent cells derived from all 3 tissues. Confirmatory studies using real-time PCR analysis were performed to validate the microarray findings for those genes involved in transcriptional regulation (Fig. 4). However, comparative studies found that the expression of LDB2, PTTG1, and TWIST-1 was higher in multipotential BMSC, DPSC, and PDLSC, while E2F2 was lower in comparison with highly proliferating human foreskin fibroblasts (Supplementary Fig. 2). RT-PCR analysis was also performed to verify the differential gene expression patterns observed for the other 20 highly expressed stem cell-associated genes (Supplementary Fig. 3).
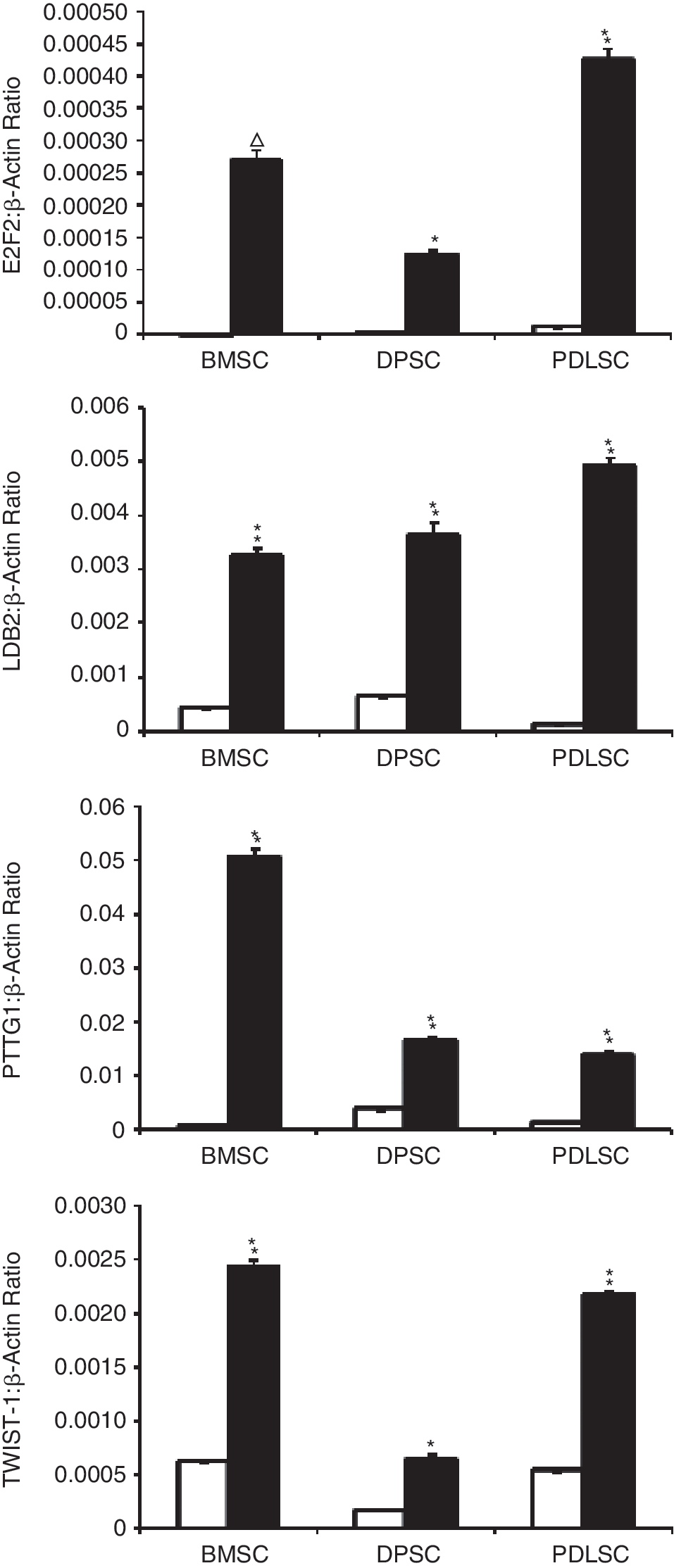
Confirmation of microarray data for genes associated with transcription by RT-PCR. Representative graphs of real-time PCR analysis of expression levels of regulators of transcription, E2F2, LDB2, PTTG1, and TWIST-1, confirm up-regulation of these genes in high growth/multidifferentiation potential clones (solid histograms) in comparison with low growth potential clones (open histograms), as primarily identified in microarray analysis. The data represent the mean values
BMSC, adult bone marrow stem cell; DPSC, dental pulp stem cell; PDLSC, periodontal ligament stem cell.
Discussion
In the present study we analyzed the genetic profiles of clonally expanded colony-forming unit-fibroblast (CFU-F) populations derived from 3 different human stromal tissues including bone marrow, dental pulp, and periodontal ligament. Our preliminary experiments focused on characterizing the differentiation and proliferation potentials of the clonally expanded cell lines isolated from each of the 3 tissue types. Proliferation studies clearly showed heterogeneity in the proliferation potentials of clonal MSC-like cells residing within bone marrow, dental pulp, and periodontal ligament tissues in accord with previous findings [5,6,8,10]. Our data suggests that only a small fraction of BMSC exhibits high proliferative potential and can undergo >20 population doublings. In contrast, approximately two-third of the DPSCs and PDLSCs exhibited an extensive proliferation capacity and an ability to exceed 20 population doublings. Essentially all the BMSC clonal lines that showed evidence of high growth potential exhibited a capacity to differentiate into cells of osteogenic, chondrogenic, and adipogenic lineages. This frequency of multipotential BMSC clones was higher than that observed for DPSCs and PDLSCs, where only 50% of the highly proliferative DPSC and PDLSC clones also exhibited multipotential differentiation potential. Data obtained from functional analysis performed on clonal populations isolated from the bone marrow, dental pulp, and periodontal ligament confirm the general hypothesis that ex vivo-expanded CFU-F from different tissues contain a heterogeneous mix of cells with varying differentiation and proliferation potentials and support previous assumptions of the hierarchical nature of stromal populations from different tissues [23]. These investigations provide a model system for identifying a common molecular fingerprint associated with immature mesenchymal stem-like cells from different organs.
Assessment of gene expression profiles comparing cells of low and high growth capacity and varied differentiation potential within the BMSC, DPSC, and PDLSC populations resulted in the identification of differentially expressed gene sets for each of the tissue types. Bioinformatic interrogation of the microarray data sets identified several hundred differentially expressed genes associated with high growth/multidifferentiation potential clonal populations. Further cross-comparative analyses of the differential gene expression patterns across all 3 stromal tissues identified 24 differentially expressed genes common to high growth/multidifferentiation potential BMSC, DPSC, and PDLSC, which were confirmed by real-time PCR. As anticipated, a large proportion of these genes have been reported to be involved in cell cycle, cell division, and mitosis, as well as in other cellular processes including DNA repair and replication and cellular differentiation. Interestingly, a small subset of the common MSC-like genes was found to be well-characterized transcription factors (E2F2, PTTG1, and TWIST-1) and a transcriptional cofactor (LDB2). As E2F family members play a major role in regulating cellular proliferation, we expected and confirmed high expression of E2F2 in high proliferative human foreskin fibroblasts that lack the capacity to differentiate into bone, fat, and cartilage. In contrast, PTTG1, TWIST-1, and LDB2 were more highly expressed by MSC-like cells in comparison with human foreskin fibroblasts. Although the role played by these transcriptional regulators in the maintenance of high growth/multidifferentiation potential MSC remains to be determined, recent studies suggest that E2F2, PTTG1, and TWIST-1 may play critical roles in BMSC growth and development.
Previous studies have shown that the transcription factors of the E2F family regulate cellular proliferation by controlling the transcription of numerous genes involved in DNA replication, DNA repair, mitosis, and cell cycle progression [24 –29]. Interestingly, studies from Won and colleagues [30] suggest that E2F1, E2F2, and E2F3 activate hTERT in normal somatic cells suggesting that activation of E2F2 in high proliferative MSC clones may maintain expression levels of hTERT and, hence telomerase activity, previously shown to greatly prolong the lifespan of culture-expanded BMSC [31 –33].
Pituitary tumor-transforming gene 1(PTTG1) was originally isolated from rat pituitary cells [34] and was termed Securin upon its latter discovery in human [35]. This transcription factor is involved in many cellular processes including mitosis, cell cycle progression, DNA repair, and maintenance of chromosome stability [35 –38]. Due to its role in chromosomal aggregation, the up-regulation of PTTG1 leads to chromosomal misaggregation and hence genomic imbalance. These events account for the oncogenic potential of up-regulated PTTG1 expression and has been associated with a variety of endocrine-related tumors and non-endocrine-related cancers [39 –46]. While the role of PTTG1 does not appear to be critical for normal fetal development, mouse embryonic fibroblasts that lack PTTG1 expression grow abnormally in culture, even though PTTG1 null mice are essentially normal [47]. However, a role for this gene in postnatal BMSC maintenance and proliferation has recently been proposed in studies describing PTTG1 deletion reduces BMSC proliferation in vitro, associated with an increased sensitivity to hypoxia and cellular senescence [48].
TWIST-1 is a basic helix-loop-helix (bHLH)-containing transcription factor [49 –52] found to be crucial for correct patterning of the skeleton [50,52 –54], where it is down-regulated during endochondral and intramembranous fetal bone development [55]. The expression of TWIST-1 appears greatest at sites of immature bone cells and decreases in more mature bone cell populations during calvaria and suture development [56,57]. The importance of TWIST-1 in bone development is highlighted in both mice and human where TWIST-1 heterozygosity leads to premature osteoblast differentiation and cranial suture fusion in the skull. These abnormalities result in craniosynostosis (Saethre-Chotzen syndrome), a condition that is often associated with dysmorphic facial findings and various limb abnormalities [58 –60]. Studies using double murine heterozygotes containing TWIST-1/Runx2 or DERMO-1/Runx2 deletions showed that both TWIST-1 and DERMO-1 (TWIST-2) negatively regulate Runx2-induced bone formation during murine development [58]. Previous studies have reported that TWIST-1 expression can be identified in different murine osteoblastic cell lines, and that this bHLH gene is down-regulated during osteogenic differentiation in vitro [55,58,61 –63]. Furthermore, work by Glackin and colleagues has shown a reduced capacity of osteosarcoma cell lines to undergo osteoblastic maturation as a result of enforced expression of either TWIST-1 or DERMO-1, whereas an increase in bone cell maturation was observed due to overexpression of TWIST-1 or DERMO-1 antisense, in these cell lines [50,52]. Recent work in our laboratory has identified a potential role for both TWIST-1 and DERMO-1 in BMSC proliferation potential and adipogenic commitment while inhibiting osteogenic/chondrogenic development [21]. Collectively, these findings implicate E2F2, PTTG1, and TWIST-1 as potential key molecules that mediate maintenance, growth, and development of mesenchymal stem-like cells derived from different anatomical sites.
A comprehensive knowledge of the genes that regulate stem cell self-renewal and differentiation potential will assist in the development of stem cell-based cell therapies. In this study, we have isolated clonal populations of BMSCs, DPSCs, and PDLSCs with high growth/multidifferentiation potential that exhibited unique genomic profiles. Among these gene data sets was a cohort of genes associated with stem cell-like characteristics. Ongoing investigations seek to examine the significance of this gene set as a signature of early mesenchymal stem cell populations derived from different tissues. This work lays the foundation for identifying novel mesenchymal stem cell markers and/or key regulators of self-renewal, growth, and commitment.
Footnotes
Acknowledgments
We would like to acknowledge the technical assistance of Mr. Krzysztof Mrozik and Dr. Naohisa Wada and bioinformatics support of Mr. Yoganand Sundaravadanam and Mr. Mark Van Der Hoek. This study was funded by National Health and Medical Research Council project grants (ID#453497 and ID#453599), Australian Dental Research Foundation Grant (62/2008), DAWES, and Betty Fanning Postgraduate Scholarships.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.