
Abstract
Germ line cell-derived pluripotent stem cells (GPSCs) are similar to embryonic stem (ES) cells in that they can proliferate intensively and differentiate into a variety of cell types. Previous studies have revealed some inherent differences in gene expression between undifferentiated mouse ES cells and GPSCs. Our aims were to generate functional hepatocytes from mouse GPSCs in vitro and to investigate whether the differences in gene expression may impact on the hepatocyte differentiation capacity of the GPSCs compared with ES cells. Mouse GPSCs and ES cells were induced to differentiate into hepatocytes through embryoid body formation, with very high efficiency. These hepatocytes were characterized at cellular, molecular, and functional levels. The GPSC-derived hepatocytes expressed hepatic markers and were metabolically active as shown by albumin and haptoglobin secretion, urea synthesis, glycogen storage, and indocyanine green uptake. We also performed an unprecedented DNA microarray analysis comparing different stages of hepatocyte differentiation. Gene expression profiling demonstrated a strong similarity between GPSC and ES cells at different stages of induced hepatic differentiation. Moreover, Pearson correlation analysis of the microarray datasets suggested that, at late hepatic differentiation stages, the in vitro-derived cells were closer to fetal mouse primary hepatocytes than to those obtained from neonates. We have shown for the first time that adult GPSCs can be induced to differentiate into functional hepatocytes in vitro. These GPSC-derived hepatocytes offer great potential for cell replacement therapy for a wide variety of liver diseases.
Introduction
E
Several strategies have been used to differentiate mouse ES cells into functional hepatocytes in vitro [2], but their use in human has been hampered by ethical problems. Adult stem cells may provide an alternative cell source for therapy [3]. Hepatocyte-like cells have been derived by directed differentiation from bone marrow, adipocytes, and mesenchymal stem cells [3]. However, when these cells are cryopreserved, a loss in viability is observed [4,5].
During the last 5 years, the developmental plasticity of pluripotent germ cells has solicited much interest. Spermatogonial stem cells have been isolated from both human and murine testis [6 –12]. Cultured spermatogonial stem cells derived from adult murine testis can undergo a spontaneous conversion process at low efficiency into pluripotent stem cells [6,10 –12]. These germ line cell-derived pluripotent stem cells (GPSCs) are similar in appearance to mouse ES cells. GPSCs can be continuously passaged under appropriate culture conditions without decline in colony-forming ability or change in karyotype [13]. These cells can also be cryopreserved and thawed with no loss in proliferation or differentiation capacity. And most importantly, no peculiar manipulation is required for “reprogramming” these adult stem cells. The GPSCs have been induced to differentiate into functional cardiomyocytes, neurons, and glia in vitro [14,15]. GPSCs are of prime importance in tissue regeneration due to the common features with both ES cells and adult stem cells. Several groups have reported the expression of the early hepatic marker, α–fetoprotein (AFP), in embryoid bodies (EBs) generated from GPSCs [10,16]. Recently, Loya et al. reported the directed differentiation of testis-derived pluripotent stem cells into hepatocyte-like cells. They demonstrated that these cells expressed hepatocyte-specific genes as seen by real-time polymerase chain reaction (RT-PCR) and secreted albumin, but no other functional characterization was shown [17]. In view of the in vivo application of GPSC-derived hepatocytes, it is important to better characterize these cells at molecular and cellular levels, as well as their functionality.
Despite sharing common properties with ES cells, GPSCs show some inherent differences as indicated by gene expression studies [6,10,12,13,18]. These 2 cell types are also different in the DNA methylation pattern of imprinted genes in that ES cells show a somatic imprinting pattern while GPSCs show an androgenic imprinting pattern [11]. It is not yet clear whether these pluripotent cells, when induced to differentiate into specific cell types, may generate cells that differ in their gene expression patterns, and hence, functionality.
In this study, we generated hepatocytes from mouse GPSCs with high efficiency and characterized their functionality. We also demonstrated that these cells could differentiate into hepatocytes as efficiently as ES cells as pointed out by large-scale gene expression profiling, and were closer to fetal hepatocytes compared with those obtained from neonates.
Materials and Methods
Culture of ES cells and GPSCs
GPSCs were derived from in vitro culture of spermatogonial stem cells isolated from 129Sv/C57B mice as described in Supplementary Methods. The cells were maintained in 5% CO2 at 37°C. After ∼3 months culture, the ES-like colonies (GPSCs) were picked and further expanded in ES cell culture media [19]. Two independently derived cell lines (GPSC#1 and GPSC#2) were generated. ES (E14) cells were used for comparison.
EBs formation and hepatocyte differentiation
To induce differentiation of ES cells and GPSCs, the method by Chinzei et al. was employed with the following modifications [20]. To form EBs, 300 cells in Iscove's modified Dulbecco's medium (IMDM), the differentiation medium containing 9% serum, were added to each well of an ultra-low attachment 96-well plate (Corning, Lowell, MA). Where indicated, partially trypsinized EBs were replated on growth factor-reduced Matrigel (BD Biosciences, Buccinasco, Italy) or in 1.5 mg/mL collagen gel (BD Biosciences; rat tail tip collagen type I; see Supplementary Methods for further details).
Culture of mouse primary hepatocytes
Primary hepatocytes (PH) from embryonic Day 16 (E16) and Day 1 neonate (PN1) livers were prepared as previously described [21]. Primary adult hepatocytes were isolated from 8-week-old mice by the 2-step collagen perfusion method [22].
Analysis of hepatic gene expression
EBs were collected at Days 2, 7, 14, 21, and 28 and total RNA extracted according to the manufacturer's protocol (Invitrogen, San Giuliano Milanese, Italy). Complementary DNA (cDNA) was synthesized from 1 μg RNA as previously described [23] and PCR was performed using the primers and conditions listed in Supplementary Table 1 (Supplementary materials are available online at
Quantitative RT-PCR (qRT-PCR) was performed to analyze Cyp7a1 and delta-like 1 (Drosophila) (Dlk1) expression using primers and probes listed in Supplementary Table 2 (Supplementary materials are available online at
Lentiviral infection of EBs and flow cytometric analysis
The lentivirus pCCL.ET.GFP.sin, which expresses GFP under the control of the enhanced transthyretin promoter, was produced in 293FT cells as previously described [24,25]. MOI was determined by infecting Hepa 1–6 cells. EBs were infected with viral supernatant (MOI = 50) containing 8 μg/mL polybrene (Sigma-Aldrich, St. Louis, MO). Positivity for GFP was assessed by flow cytometry (BD Biosciences). Data were analyzed using the CellQuest software. Noninfected EBs were used as negative control.
Characterization of GPSCs and EBs
GPSCs were stained for Oct4, Nanog (Abcam, Cambridge, UK) and analyzed by confocal microscopy (Leica Microsystems, TCS SP5 II, Italy). Flow cytometry was performed after staining of GPSCs with stage-specific embryonic antigen-1 (SSEA-1). Teratoma was generated in NOD/SCID mice using GPSC#1. Teratoma sections were stained with hematoxylin and eosin, and with anti-mouse Liv2 antibody (MBL International, Woburn, MA) and revealed with biotinylated anti-rat antibody and the ABC complex (DAKO, Glostrup; see Supplementary Methods for details).
Immunofluorescence staining of EBs
EBs were stained for albumin, cadherin, and Dlk1 (Abcam) as described in Supplementary Methods.
Albumin and haptoglobin secretion
Albumin was measured in single EB culture supernatant at various time points with a mouse albumin enzyme-linked immunosorbent assay (ELISA) quantitation kit (Bethyl laboratories, Montgomery, TX) according to the manufacturer's instructions and normalized to protein content of single EBs. Haptoglobin (Hp) was analyzed by western blotting as previously described [26].
Urea production
One EB was plated per well on Matrigel and 5 mM ammonium chloride was added for 24 h. Urea production was determined at various time points using a colorimetric assay (Quantichrom, Hayward, CA) following the manufacturer's instructions and normalized to protein content of single EBs.
Periodic acid Schiff staining
EBs were stained with Periodic acid Schiff (PAS) reagents according to the manufacturer's instructions (Bio Optica, Milan, Italy). Mouse embryonic fibroblasts (MEFs) were used as negative control.
Indocyanine green uptake study
Indocyanine green solution (final concentration 1 mg/mL, Sigma-Aldrich) was added to the EBs as described previously [27].
RNA extraction and microarray analysis
Total RNA was extracted from ES cells and GPSC#1 and GPSC#2 during hepatocyte differentiation at indicated time points, and from E16 and PN1 PH using miRNeasy kit (QIAGEN, Milan, Italy). DNA microarray analysis was performed as described in Supplementary Methods using MOUSEWG-6_V2 Beadchips (Illumina, San Diego, CA). Data clustering was performed with the GEDAS software [28]. qRT-PCR was employed for experimental validation (Supplementary Table 2).Data were normalized to 18S expression (Applied Biosystems, Monza, MI).
Gene ontology and KEGG pathway enrichment analysis
Clusters of coexpressed genes and sets of genes highly expressed in the late differentiation stage (E16 PH, ES cells, and GPSCs on Day 28) were analyzed for statistically significant enrichment in Gene ontology (GO) annotations [29] and KEGG pathways [30,31] by computing P values according to 1-tailed Fisher's exact test. Only annotations with a false discovery rate (FDR) ≤5% were accepted. For more details, see Supplementary Methods.
DNA methylation analysis
H19 differentially methylated region (DMR) methylation of ES cells and GPSCs as well as Oct4 and Nanog promoter methylation analyses of spermatogonial stem cells versus GPSCs were determined by bisulfite sequencing as described in Ko et al. [11].
Statistical analysis
Data are expressed as mean ± SD. Statistical differences were determined by a 2-tailed Student's t-test (*P < 0.05, **P < 0.01, ***P < 0.001). All experiments were performed independently at least 3 times.
Results
Characterization of GPSCs and hepatocyte differentiation
Thy1+/αv-integrin− spermatogonial stem cells were sorted from juvenile mouse testis and cultured on a MEF feeder layer. The ES-like cells or GPSCs (Fig. 1A and Supplementary Fig. 1A; Supplementary materials are available online at

In vitro characterization of germ line cell-derived pluripotent stem cell (GPSC)-derived hepatocytes. (
In order to induce differentiation of GPSCs into hepatocytes, EBs were formed by static culture and a specific hepatocyte differentiation protocol was employed (see Materials and Methods section). During the course of directed differentiation, the EBs were analyzed morphologically. At Day 7, typical polygonal-shaped cells with granular cytoplasm were observed, and on Days 14 and 21, binucleated cells with prominent cytoplasm, characteristic of hepatocytes, were present in the EB outgrowths (Fig. 1A).
Expression of liver-specific genes in the GPSC-derived EBs
To assess the differentiation of GPSCs into hepatic lineages, we analyzed the mRNA expression of liver-specific genes albumin, AFP, tyrosine amino transferase (TAT), hemopexin (Hx), Hp as well as the pluripotency gene Oct4 by RT-PCR. As shown in Figure 1B, albumin was expressed on Day 14 and lasted till Day 28, the latest time point analyzed. AFP mRNA was expressed as early as Day 2 in EBs, showing endodermal differentiation and fetal hepatic formation and, continued to increase until Day 21, after which a slight decrease was observed. Expression of the enzyme TAT, characteristic of maturing hepatocytes, was observed as from Day 21. Hx and Hp, which are highly expressed in hepatocytes in vivo, were expressed as from Days 14 and 21, respectively. No expression of albumin, AFP, TAT, Hx, or Hp was noted in the undifferentiated GPSCs. Moreover, the expression of the liver-specific gene Cyp7a1 confirmed hepatic differentiation in EBs derived from GPSCs (Fig. 1C). On the other hand, Oct4 mRNA gradually decreased during the course of directed differentiation. A similar expression pattern was obtained with ES cells (data not shown). These results show that GPSCs expressed hepatic genes during induced differentiation.
Percentage of hepatocytes in GPSC-derived EBs
In order to evaluate the percentage of cells differentiating into hepatocytes, EBs were infected with lentivirus expressing GFP under the hepatocyte-specific enhanced transthyretin promoter. Analysis by fluorescence microscope revealed clusters of polygonal-shaped cells that were highly positive for GFP (Fig. 2A). Our cultures also consisted of other cell types including neuron-like cells that were not positive for GFP. Flow cytometric analysis showed that on Day 21, 70.16% of cells in the GPSC-derived EB cultures were GFP-positive, comparable with those obtained from ES cells (data not shown). Interestingly, this percentage increased with time hence showing high efficiency of hepatocyte conversion of GPSCs (82.68% on Day 35, Fig. 2B). Only 7.7% of the nondifferentiated cells were GFP-positive, showing that at high MOI (MOI = 50), enhanced transthyretin promoter drove a low but detectable level of expression after high vector input [24].
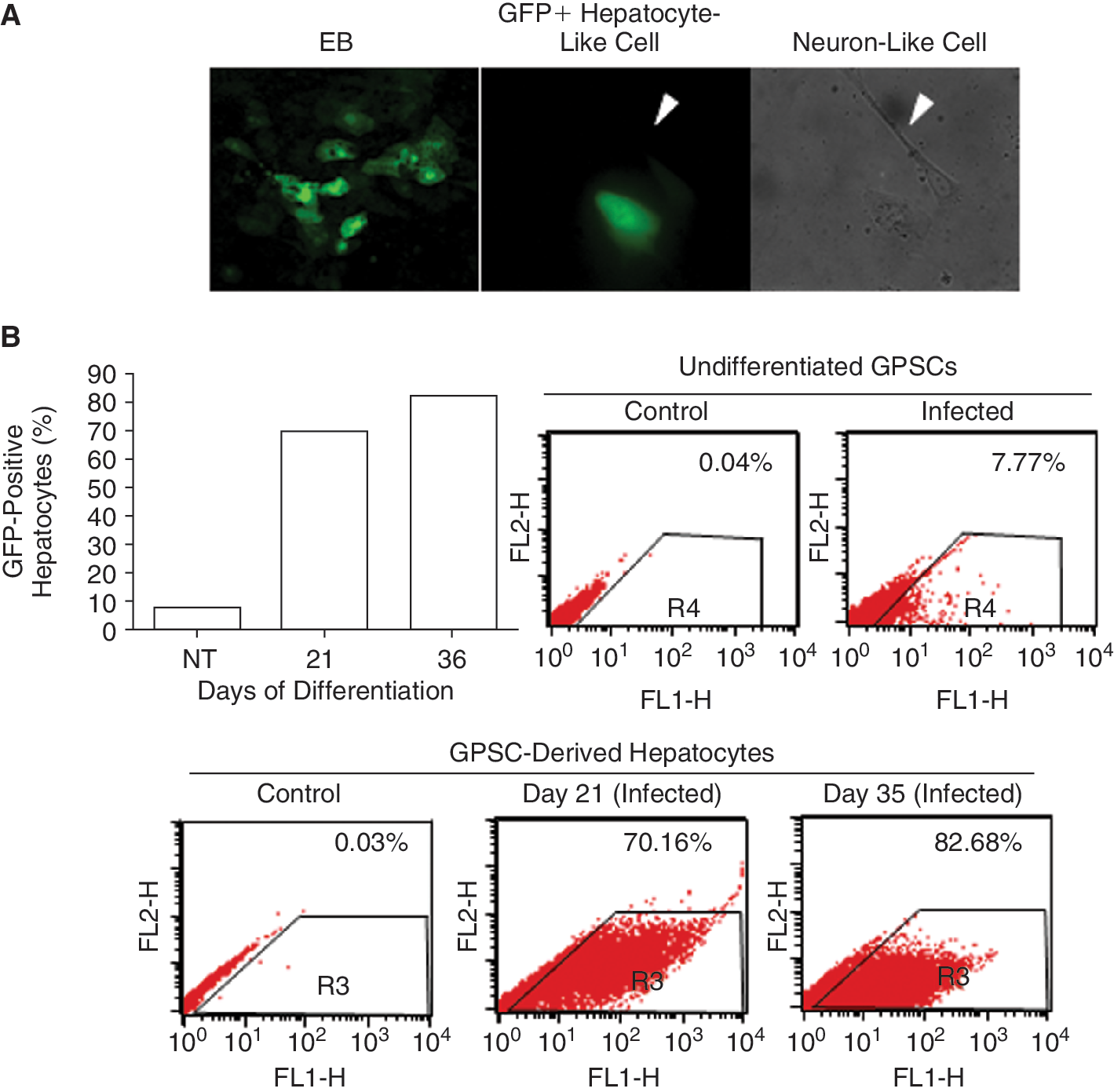
Percentage of hepatocytes in germ line cell-derived pluripotent stem cell (GPSC)-derived embryoid bodies (EBs). (
Immunofluorescence analysis of hepatocyte markers
Expression of the hepatocyte markers, albumin on Day 14, Dlk1 and E-cadherin on Day 13 was analyzed by immunofluorescence (Fig. 3A) in the GPSC-derived EBs. Albumin-positive staining was observed in the cytoplasm of the monolayer cells around the EBs. These cells were also positive for Dlk1 and E-cadherin. Albumin and Dlk1 staining could be seen more clearly in the larger cells of the EB outgrowths (Fig. 3B). Interestingly, qRT-PCR analysis revealed that Dlk1 was still highly expressed in the Day 28 EBs (Fig. 3C) suggesting that the EB cultures still contained hepato-cyte precursors.
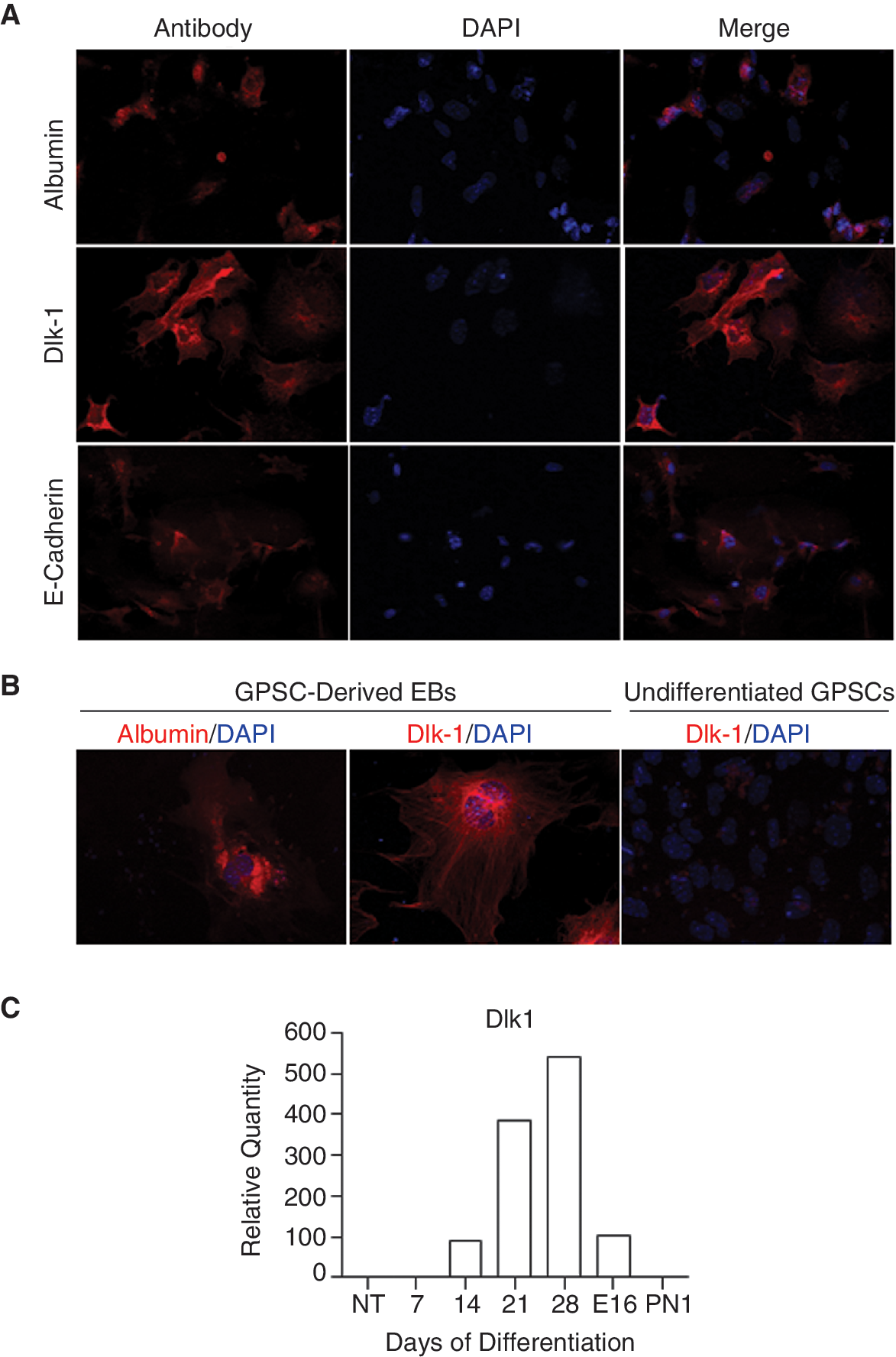
Analysis of hepatocyte markers in the differentiated germ line cell-derived pluripotent stem cells (GPSCs). (
Protein secretion by GPSC-derived hepatocytes
Albumin secretion was analyzed in culture supernatants of single GPSC-derived EBs plated on Matrigel on Days 7, 11, 14, 18, and 21. A statistically significant increase in albumin secretion was observed on Day 11, followed by a gradual decrease to Day 18. Interestingly, when EBs were trypsinized on Day 17 and replated in collagen gel, albumin secretion was restored (Fig. 4A). Hp was also secreted from the EBs on Day 28 of differentiation (Fig. 4B). Similar results were obtained with ES cells (data not shown). Hence, these results demonstrated that GPSC-derived hepatocytes could secrete liver-specific protein when cultured in a 3D setting.
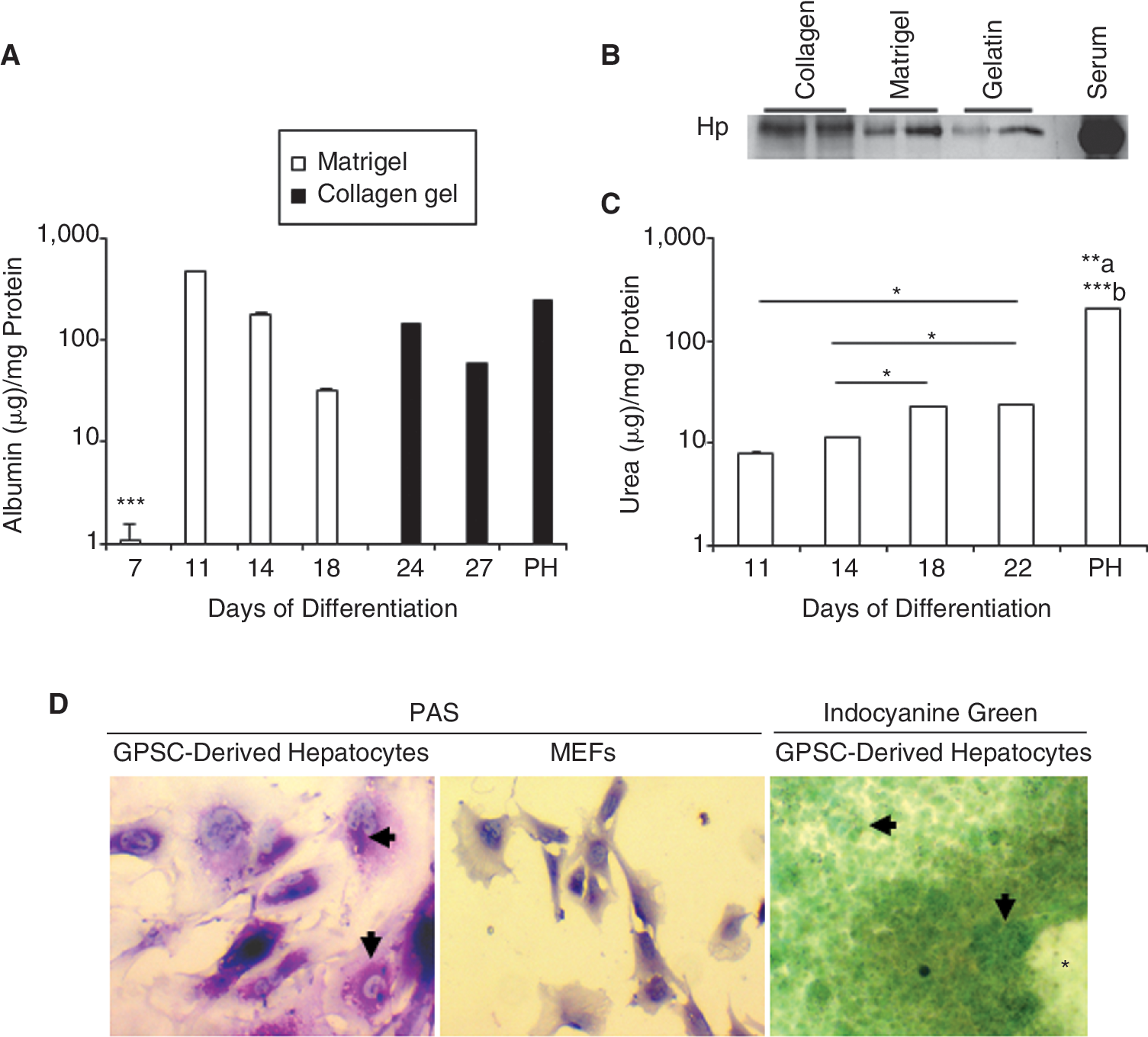
Functional characterization of germ line cell-derived pluripotent stem cell (GPSC)-derived hepatocytes. (
Hepatocyte metabolic activity
We assessed for the production of urea in order to determine the functionality of the GPSC-derived hepatocytes. Substantial urea synthesis was first observed on Day 11 and increased significantly up to Day 22, the last time point analyzed (Fig. 4C). Moreover, PAS staining showed intra-cellular glycogen deposits in the cytoplasm of hepatocytes derived from GPSCs but not in MEFs (Fig. 4D). Indocyanine green-positive cells were also seen as green clusters in the GPSC-derived EBs, most of which were adjacent to the cardiac muscle-like cells (Fig. 4D). These results show that the GPSC-derived hepatocytes were functional.
Hepatocytes derived from GPSCs and ES cells in vitro show similar expression profile to fetal PH
In order to compare the genome-wide gene expression profile of hepatocytes derived in vitro from 2 independent GPSC lines (GPSC#1 and GPSC#2) and ES cells at different stages of differentiation to those isolated from mice livers (E16 and PN1), DNA microarray analysis was performed. The first analysis was done with an unsupervised approach. Clustering analysis of the 1,443 probes (out of 17,482) displaying a standard deviation >1 (an average fold change >2; see Supplementary Methods) indicated a striking similarity in the gene expression profile among GPSC#1, GPSC#2, and ES cells. Clusters 1–4 show genes that were up-regulated at late differentiation times in the EBs from the 3 cell lines, but were differentially expressed in the PH isolated from E16 and PN1 mice (Fig. 5A). Clusters 5–7 represent genes that were up-regulated in the EBs at late differentiation times as well as in the PH, showing a liver-like response (Fig. 5A). Clusters 8–10 also contain genes that showed a similar expression pattern in the EBs in the early, late, and very late stages of differentiation, respectively, with respect to PH (Supplementary Fig. 2A). Conversely, Clusters 11–15 show genes that were unaffected by differentiation in all 3 cell lines, but were differentially expressed in the PH (Fig. 5B and Supplementary Fig. 2B and Supplementary Table 3; Supplementary materials are available online at
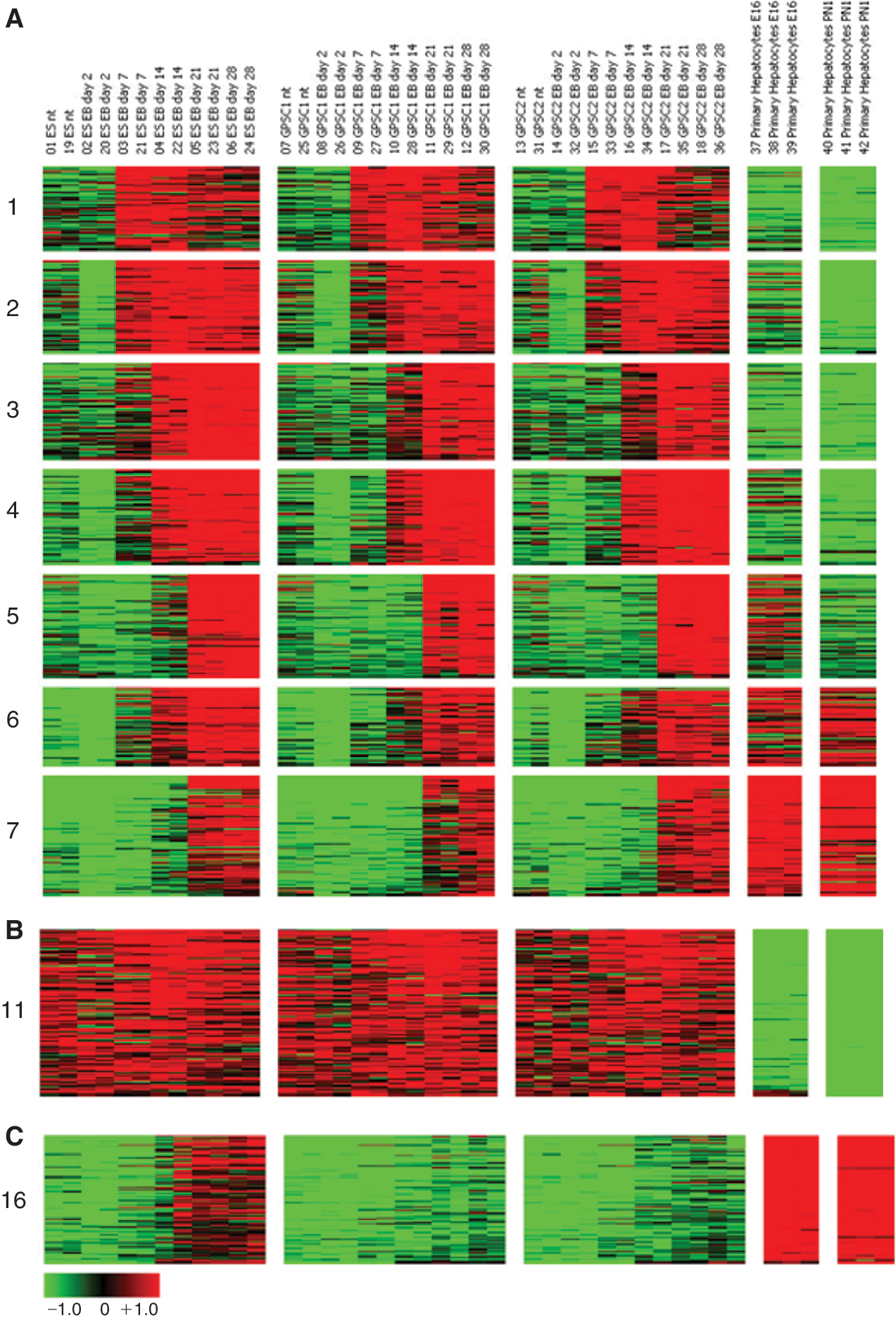
Gene expression changes across multiple cell lines and hepatic differentiation stages. Undifferentiated (nt) embryonic stem (ES) cell, germ line cell-derived pluripotent stem cell (GPSC)#1, GPSC#2 versus Days 2, 7, 14, 21, 28 of embryoid body (EB) differentiation versus E16 and PN1 primary hepatocytes. For sake of clarity, only representative clusters as visualized by GEDAS are shown. Color code as shown in scale bar. See Supplementary Figure 2 for the remaining clusters. Color images available online at
Importantly, to determine the differentiation status of the hepatocytes derived in vitro, we used all 17,482 probes (see Supplementary Methods) detected to compare the expression profiles of EBs derived from the 3 cell lines at different stages of hepatic differentiation with those of the PH from E16 or PN1 livers, using a Pearson's correlation analysis. The results indicated that the hepatocytes obtained in vitro at Day 28 of differentiation were closer in gene expression profile to the E16 hepatocytes (Fig. 6A) compared with PN1 hepatocytes (Fig. 6B).
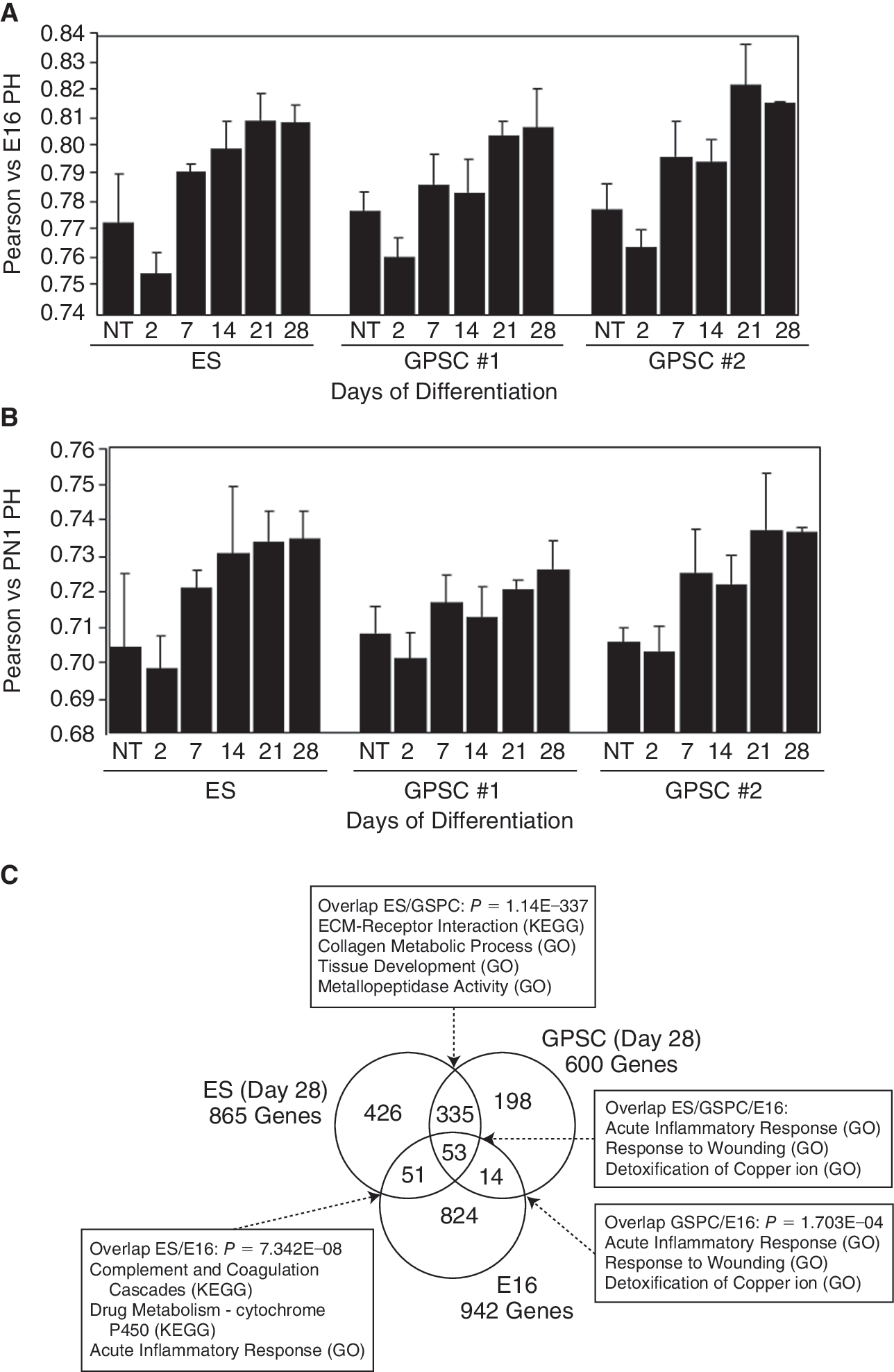
Comparison of global gene expression. (
The similarity between ES and GPSCs at late differentiation stages is exemplified when comparing exclusively the genes having at least 2-fold higher expression in E16 PH, and ES and GPSC at Day 28 of differentiation than their average expression level over all samples (see Supplementary Methods). As shown in Figure 6C, the number of genes highly expressed in both ES and GPSC on Day 28 is extremely significant (P = 1.14 × 10−337).
Identification of genes differentially expressed during hepatocyte differentiation
As an additional supervised approach, genes with similar expression in the E16 and PN1 PH as well as in the EBs on Days 21 and 28, but differentially expressed in the undifferentiated cells (GPSCs and ES cells) were searched for. The results of a t-test (P value = 0.00001) and a fold change analysis (log2 ratio of ±2), performed independently, were combined and resulted in 266 genes with both significant and robust expression change. The genes that were up-regulated in E16 and PN1 PH as well as in the EBs at late differentiation time were grouped as shown in Figure 7A. Genes up-regulated during differentiation in the 3 cell lines comprised hepatocyte metabolic enzymes like alcohol dehydrogenase 1 (Aldh1), UDP glycosyltransferase 1 family, polypeptide A10, hydroxysteroid 11-β-dehydrogenase 1, serum proteins like lipocalin 2, Hp, serum amyloid A3 (SAA3), serine protease inhibitors like Serpina3n and Serpina3g, hormones like insulin-like growth factor 1 (IGF-1), and iron-storage proteins like ferritin heavy chain 1 (FtH1) (Fig. 7A). Interestingly, gene expression profiling showed that Dlk1 was expressed in the EBs during hepatocyte differentiation from Day 7 and Day 14 in the ES- and GPSC-derived EBs, respectively. The expression of this gene was still high on Day 28, the last time point analyzed (Cluster 4 in Fig. 5A and Supplementary Fig. 3A; Supplementary materials are available online at
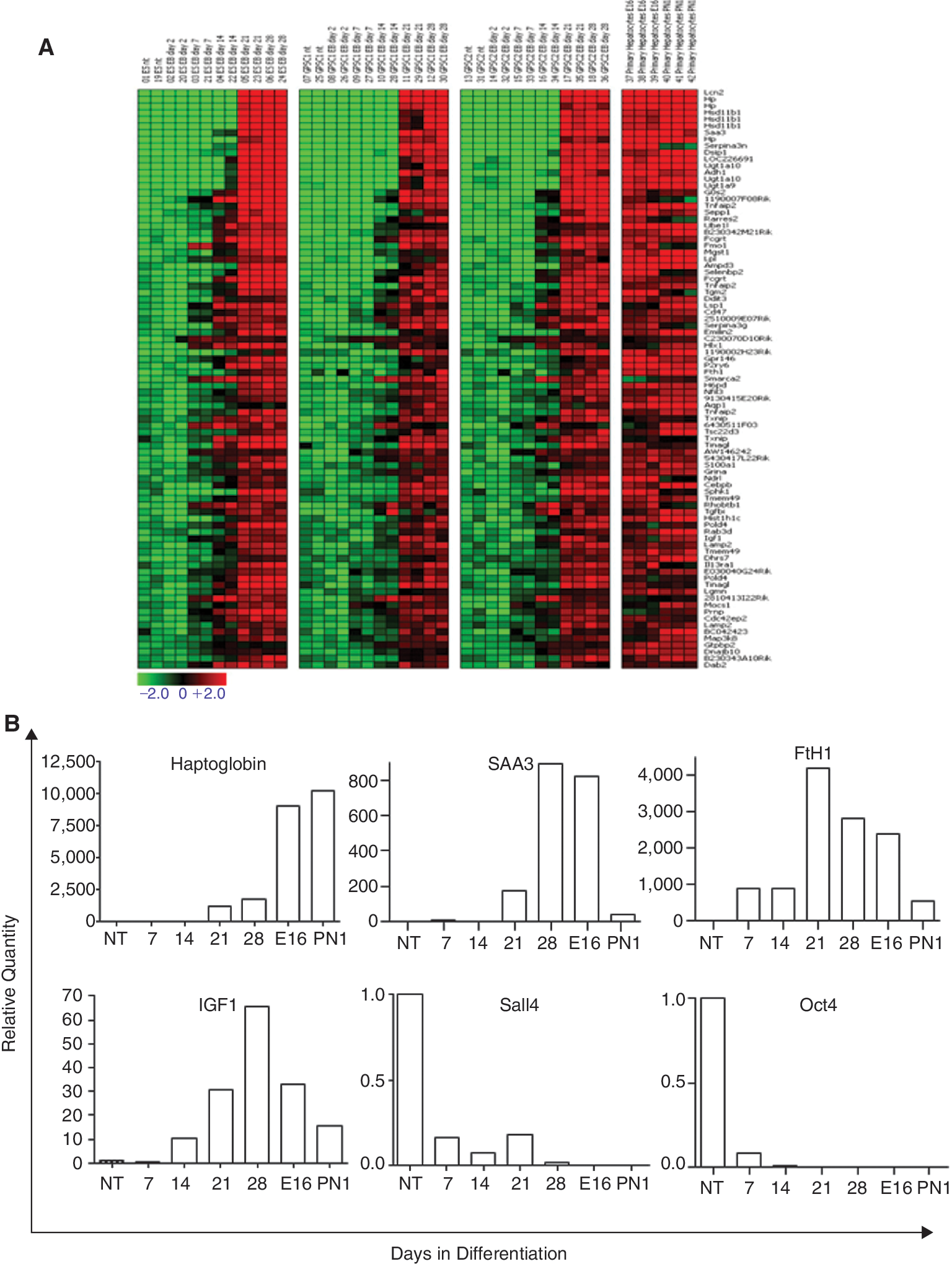
Gene expression profile of genes up-regulated during induced hepatic differentiation of embryoid bodies (EBs). (
We also compared our microarray data with those of Guo et al. [33], who identified by gene expression profiling a transcriptional signature discriminating mouse fetal liver at different developmental stages (E11.5, E14.5, E15.5, and 3 dpp). A Pearson's correlation analysis for the 845 genes of this signature mapped on our dataset (see Supplementary Methods) revealed that the gene expression profile of ES cells as well as GPSCs on Days 21 and 28 of hepatocyte differentiation was much closer to 3 dpp livers, where the hematopoietic component becomes negligible (Supplementary Fig. 4C; Supplementary materials are available online at
Functional analysis of gene clusters
To gain further insight into the biological significance of the microarray results, we performed functional analysis of Clusters 1–16 by identifying significant enrichments in GO annotations and KEGG pathways. Detailed results are listed in Supplementary Tables 5 and 6 (Supplementary materials are available online at
The liver-like response is best evidenced by the KEGG pathway enrichments of Cluster 7 (Supplementary Table 6), which include different metabolic pathways (eg, androgen and estrogen, retinol, pentose and glucuronate, starch and sucrose, and xenobiotics by cytochrome P450) as well as antigen processing and presentation.
Cluster 12 is enriched in genes involved in the immune system, complement cascade, and coagulation pathway (Supplementary Table 5), concording with the ability of the liver to synthesize acute phase proteins as well as complements and coagulation factors [34,35]. These genes were not up-regulated during ES or GPSC differentiation (Supplementary Fig. 2B). Overall, the gene expression patterns of ES cells and GPSCs, during hepatocyte differentiation, showed strong similarity (see also Fig. 6C).
Discussion
Adult stem cells have demonstrated great differentiative potential and may provide an unparalleled opportunity for the treatment of a wide range of liver diseases [36]. The similarity between mouse ES cells and GPSCs offer the possibility of shifting to GPSCs for therapeutic studies, thus circumventing ethical problems. In this study, we generated hepatocytes from GPSCs through the formation of EBs, which are known to recapitulate early embryogenesis in vitro, and by addition of stage-specific growth factors. The resulting polygonal-shaped and binuclear cells expressed a panel of hepatic genes. To our knowledge, this is the first report showing that metabolically active hepatocytes, capable of urea synthesis, glycogen storage, and indocyanine green uptake can be derived from GPSCs in vitro. GPSCs differentiated into hepatocytes with high efficiency (70.16% on Day 21), which was comparable with that of ES cells. A further increase in number of GFP-positive hepatocytes was observed with time (82.68% on Day 35) in our system, as opposed to Loya et al. who noted a decrease in hepatic gene expression after Day 3 [17]. We also demonstrated that GPSC-derived hepatocytes bore secretive activity as shown by the presence of albumin and Hp in the culture supernatants. When the EBs were cultured in monolayer, albumin secretion decreased with time, but when these were partially trypsinized were replated in collagen gel, protein secretion was restored. This indicates that the 3D setting is important for the cells to maintain their metabolic activity. Protein secretion by stem cell-derived hepatocytes is an important aspect especially when replacement therapy aims at restoring liver functions, as for example, albumin secretion in cirrhotic patients.
Previous gene expression studies have searched for differences between undifferentiated mouse ES cells and GPSCs. During the transition from spermatogonial stem cell to GPSC, the latter acquire the expression of ES cell-specific genes, like Oct4, Nanog, Sox2, SSEA-1, although differences are evident in the expression of genes like Dazl, Gdf3, Stra8 [6,10,12,37]. We thus wanted to investigate whether this difference in gene expression may impact on the hepatocyte differentiation capacity of the GPSCs. To our knowledge, no large-scale comparative analysis has been performed on the hepatocytes derived from these 2 cell types during the different stages of hepatocyte differentiation. DNA microarray analysis conducted during our study suggested that GPSCs generated hepatocytes as efficiently as ES cells and with similar kinetics. Genes involved in different hepatocyte functions, like serum protein production, detoxification, hepatocyte specification, were up-regulated by at least 4-fold in ES cells and GPSCs at late differentiation stages, as well as in the PH (See Results section and Fig. 7A). On the other hand, we observed at least 4-fold decrease in the expression of Oct4, Sall4, Dppa5, Utf1, Zfp42 on Day 14 of differentiation of GPSCs and ES cells into hepatocytes (Supplementary Table 4)consistent with their known function in pluripotency maintenance.
Besides these similarities, we also observed some differences between ES cells and GPSCs when induced to differentiate into hepatocytes. For instance, Cluster 13 shows genes that were expressed in GPSCs but not in ES cells and includes genes like testis-expressed 9 (Tex9) (Supplementary Table 7; Supplementary materials are available online at
Studies performed on rats have shown that cells positive for the transmembrane protein and hepatoblast marker, Dlk1, contained repopulating activity [40]. We observed that the expression of Dlk1 was still high on Day 28 in the EBs derived from GPSCs, as well as in those from ES cells. In agreement, AFP, an endodermal and early hepatic marker, was still up-regulated on Day 28 indicating that a high percentage of cells might still be in early stages of hepatic differentiation and might be highly proliferative as also suggested by the enrichment of Cluster 1, and particularly, Cluster 11. The Pearson's correlation analysis performed on all 17,482 DNA microarray probes detected also indicated a closer similarity between GPSC-derived hepatocytes and fetal hepatocytes compared with those from neonates. This property may be crucial for liver colonization when the GPSC-derived hepatocytes are transplanted in vivo, as immature hepatocytes have increased proliferative activity compared with mature ones, as reported by Sandhu et al. [41].
In the mouse fetal liver, at E15.5 there is an important switch toward liver-related functions [33]. When induced to differentiate into hepatocytes, the ES cells and GPSCs at Days 21 and 28 were closer to fetal hepatocytes (E16 onward), which were less hematopoietic than E15.5, indicating a strong tendency toward liver development.
In conclusion, we show for the first time that functional hepatocytes can be derived from GPSCs in vitro with very high efficiency. Future studies will show whether GPSC-derived hepatocytes have the ability to restore hepatic function in disease models, and hence offer the opportunity to produce immune-matched hepatocytes for patient-specific treatment.
Footnotes
Acknowledgments
We thank Luigi Naldini and Alessio Cantore for providing pCCL.ET.GFP.sin lentiviral vector; Maddalena Ianicella and Antonellisa Sgarra for technical assistance; Annalisa Camporeale for help with flow cytometry; Elisa De Luca for confocal microscopy; Ferdinando Di Cunto for helpful discussion; Rinaldo Pellicano for helpful discussion and critical reading of the manuscript; and Guido Tarone for critical reading of the manuscript. This work was supported by the following grants to F.A.: Piattaforma Regione Piemonte Pi-Stem, Compagnia San Paolo, Telethon (GTF05010), and ex 60% MIUR.
Author Disclosure Statement
No competing financial interests exist.
The raw and normalized microarray data have been deposited on the Gene Expression Omnibust database (accession number: GSE19044).
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.