
Abstract
Bidirectional cell trafficking between fetus and mother during pregnancy is a well-established phenomenon observed in placental vertebrates including humans. Although studies have shown that transmigratory fetal cells, also termed pregnancy-associated progenitor cells (PAPCs), can integrate into multiple maternal organs, the integration, long-term survival, and differentiation of PAPCs in the brain has not been extensively studied. Using a murine model of fetomaternal microchimerism, we show that PAPCs integrated and persisted in several areas of the maternal brain for up to 7 months postpartum. Besides expressing neural stem cell or immature neuronal markers, PAPCs were observed to express mature neuronal markers, indicating that PAPCs adopted a neuronal fate. Further, PAPCs also displayed morphologically neuronal maturation by an increasing axonal/dendritic complexity over time. Therefore, PAPCs seem to undergo a molecular and morphological maturation program similar to that observed during adult neurogenesis. We provide evidence that neuronal gene expression of PAPCs was not a consequence of cell fusion with maternal neurons. In addition, in mothers with experimentally induced Parkinson's disease (PD), the frequency of PAPCs within the hippocampus initially increased whereas long-term presence of PAPCs was compromised. Also, the spatial distribution of PAPCs within the hippocampus was altered in mothers with PD. Thus, the disease context influenced the initial attraction, long-term survival, and spatial distribution of PAPCs, which may have wider implications on cell replacement strategies in human neurodegenerative diseases such as PD.
Introduction
C
Here, we show that PAPCs can be found at several anatomical sites within the maternal brain in which they acquired specific neuronal marker gene expression. The expression of neuronal marker genes was not a consequence of PAPCs fusing with maternal neurons, suggesting an intrinsic neuronal differentiation potential of PAPCs. We observed progressive maturation of axonal and dendritic arborizations of PAPCs and the expression of neural stem cell as well as early neuronal differentiation markers. Together, this suggests that PAPCs undergo a molecular and morphological differentiation process that resembles endogenous adult neurogenesis. Further, PAPCs were responsive to a PD environment, as they were initially attracted to the lesioned hippocampus (HC) of a murine PD model. However, long-term survival of PAPCs was compromised, as in older postpartum PD mothers lesser PAPCs were detected and the overall distribution pattern of cells was markedly changed. This suggests that the PD environment did not support long-term homing, engraftment, and survival of PAPCs.
Materials and Methods
Mice strains
Wild-type (WT) young adult virgin female C57BL/6j mice (8–12 weeks old) were used. Males of C57BL/6 Cr Slc TgN (act-eGFP) OsbC14-Y01-FM131 [26] or B6. Cg-Tg (Thy1-YFP) 16Jrs/J mice [27] were mated with WT female mice. Rosa26-lacZ mice (129-Gt(ROSA)26Sor/J) were used as recipients for the cell fusion study. Double transgenic act-eGFP:ROSA26lacZ mice were generated by mating act-eGFP and ROSA26lacZ. All animal experiments were conducted with approval from the Institutional Animal Care and Use Committee.
Quantitative real-time polymerase chain reaction
Maternal mice were perfused with 0.9% saline, and the brain was removed. The olfactory bulb, cortex, striatum, midbrain, HC, and cerebellum were isolated. Genomic DNA was extracted. The primer and probe sequences and detailed protocols are described elsewhere [11]. All samples were run on an ABI PRISM 7000 Sequence Detection System (Applied Biosystems). Data are expressed as mean ± standard error of the mean.
Digestion of maternal brain
Mothers were anesthetized and perfused with saline. Brains were harvested and the olfactory bulb, striatum, cortex, midbrain, HC, and cerebellum were dissected. Each part was chopped into small pieces, and pre-warmed 0.125% trypsin solution was added. Tissues were incubated at 37°C for 5 min followed by aspirations. The aspirate was neutralized with 10 mL of Dulbecco's modified Eagle medium (DMEM) supplemented with 20% fetal bovine serum and was spun for 5 min at 200g. The pellet was re-suspended in 10 mL of fresh medium. The cell suspension was plated in 24- well plates and inspected for eGFP+ve cells under an epifluorescence microscope.
Y-chromosome fluorescence in situ hybridization analysis of PAPCs
In situ hybridization was done as described elsewhere [11]. Briefly, sections were incubated with methanol and acetic acid and incubated with sodium thiocyanate solution (1 M). Sections were denatured with 60% formamide and hybridized with the probes. Sections were counterstained with DAPI and mounted with an anti-fade mounting medium.
Detection and quantification of eGFP PAPCs in the maternal brain
Mice were perfused with saline and 4% PFA at prenatal stages E12, E16, and postnatal stages postpartum day 0 (P0), P7, P60, and P120. Coronal cryosections (20 μm) from olfactory bulb to cerebellum were mounted. Ten to 12 serial sections from each area of the brain were collected, and eGFP+ cells were quantified.
Immunohistochemistry
Serial consecutive coronal hippocampal sections (35 μm) were cut on a cryostat and evenly distributed into 10 sets containing 5 sections each. Mothers (n = 6) were used for quantifying the expression of neural marker genes. For the quantification of total fetal cells, 1 set of sections of the HC of each mother was analyzed, and the total amount of fetal cells was determined in each set by epifluorescence microscopy. Immunohistochemistry was performed following standard procedures. Primary antibodies used were nestin, poly-sialic acid neural cell adhesion molecule (PSA-NCAM), NeuN, β3-tubulin, microtubule-associated protein 2 (MAP2), myelin-associated glycoprotein, glial fibrillary acidic protein (GFAP) (all Chemicon), doublecortin (DCX) (Santa Cruz), and Calbindin-D28 (gift from Dr. Liang, National University of Singapore). For the analysis of marker gene expression, 9 sets of sections (5 sections each) from the HC were used. Each set was stained with 1 antibody (see above) and was analyzed. Representative double-labeled cells were recorded with a confocal microscope (Zeiss LSM). Morphological maturation of PAPCs: hippocampal sections (20μm) of mothers at P7, P60, and P120 were screened for eGFP+ cells. z-stacks were obtained from individual eGFP cells using confocal microscopy. eGFP+ cells were then manually projected on 2D drawings and branching points, and branching complexity was quantified.
Blastocyst transfer
WT females were time mated with eGFP males. Three days after plug formation, blastocysts were isolated from the uterus of these females. Twelve to 16 blastocysts were implanted into individual Rosa26 (129-Gt(ROSA)26Sor/J) pseudo-pregnant females. Mothers with successful pregnancies having eGFP+ve pups were sacrificed by anesthetic overdose and perfusion with 4% PFA at indicated time points. Serial coronal brain sections (20 μm) were cut, and anti-β-Gal immunohistochemistry was performed. Stained sections were screened for double-labeled cells.
Induction of PD in a murine model
C57BL/6j females were injected with 6-OHDA (2 μL of a 4 μg/μL in 0.02% ascorbic acid solution) into the right striatum using Bregma coordinates (AP: + 1.0; ML: +1.5; DV: +3.0). Apomorphine was subcutaneously injected (0.5 mg/kg), and rotations were recorded for 45 min using a Rotometer (TSE Systems) with automated recording software (TSE systems). Mice with >5 rotations min−1 were selected and mated with eGFP males. Fetal cell presence in the HC was quantified as described earlier.
Statistical analysis
Data are expressed as means ± standard error of the mean. For comparison between groups, the Student's t-test was performed.
Results
Presence and quantification of PAPCs in maternal brain
To characterize PAPCs in maternal brain, male mice expressing eGFP under the chicken actin (CAG) promoter were mated with WT females, and eGFP expressing PAPCs were tracked within maternal brains. Hippocampi (HC) of P7 mothers (n = 4) were microdissected, dissociated into single-cell suspensions, and screened for the presence of eGFP+ cells. PAPCs were detected in 2 out of 4 HC, whereas no eGFP+ cells were found in nonpregnant controls (data not shown). Fluorescence in situ hybridization using a Y-chromosome-specific probe was performed to detect male fetal cells in mothers. Male cells were observed in HC (Fig. 1A) as well as in 5 other areas of the brain (Supplementary Fig. S1, available online at

PAPCs are detected in the maternal brain.
Time course of integration, long-term survival, and spatial distribution of PAPCs in the HC
We focused our analysis on the HC, as it is a well-studied anatomical and functional brain structure using the direct eGFP-detection method. The time course of integration was examined by analyzing mothers at specific time points postcopulation (E12 and E16) and at P0 (n = 6), P7 (n = 9), P30 (n = 7), P60 (n = 9), P120 (n = 7), and P210 (n = 3) (Fig. 2A and Supplementary Table S1, available online at
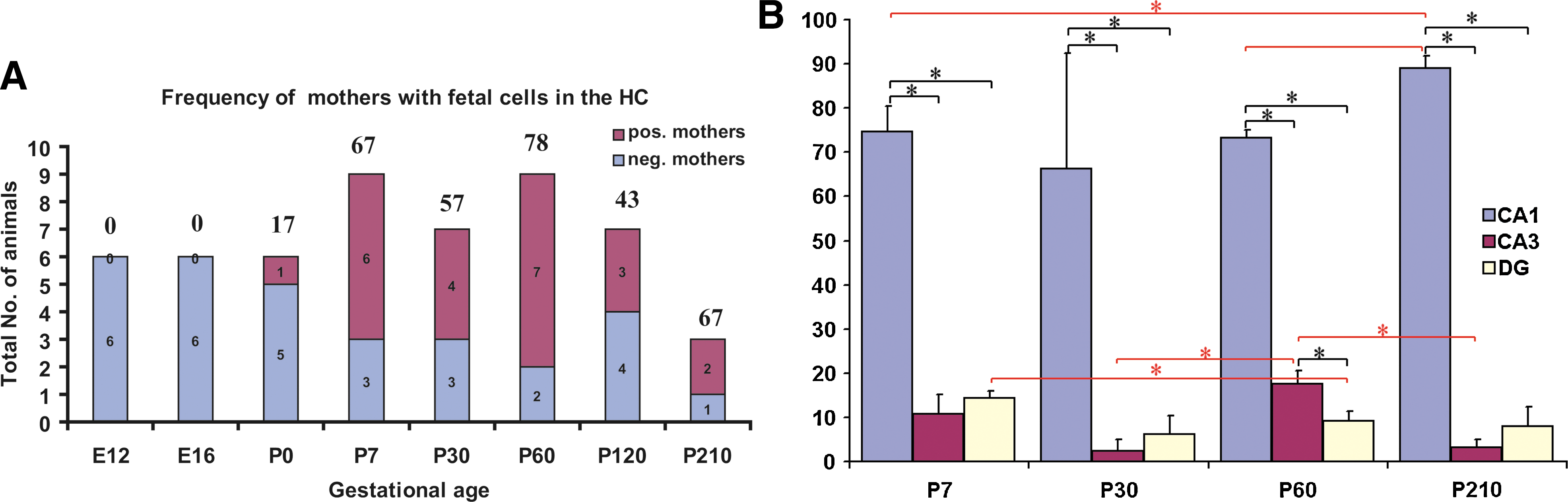
Temporal and spatial analysis of PAPC frequency and distribution within the HC.
eGFP+ fetal cells were found at the 3 major anatomically sites within HC, that is, CA1/2, CA3, and the dentate gyrus (DG). Detailed quantification of cellular distribution in the substructures of the HC was performed at P7 (n = 3), P30 (n = 3), P60 (n = 3), and P210 (n = 2) and it was found that on average, PAPCs were predominantly located in the CA1/2 (82.0% ± 10%) and to a lower frequency in the CA3 area (9.7% ± 7.6%) and in the DG (8.4% ± 4.7%) (Fig. 2B). This distribution pattern remained relatively unchanged across the different time points examined.
PAPCs express neuronal cell markers
PAPCs showed axonal and dendritic morphologies reminiscent of neurons, suggesting that they acquired neuronal cell fate. Therefore, we investigated whether PAPCs in the HC of mothers (n = 6) expressed neuronal and glial-specific markers. We found that PAPCs could express markers such as β3-tubulin (31.2%, n = 900; Fig. 3A–C), MAP2 (5.6%, n = 900; Fig. 3D–F) NeuN (28.5%, n = 1,056; Fig. 3G), and calbindin (5.7%, n = 1,056; Fig. 3H), suggesting that they acquired neuronal-specific marker gene expression. PAPCs in other brain areas also expressed neuronal-specific markers, demonstrating that PAPCs generally could acquire neuronal marker gene expression (Supplementary Fig. S2, available online at
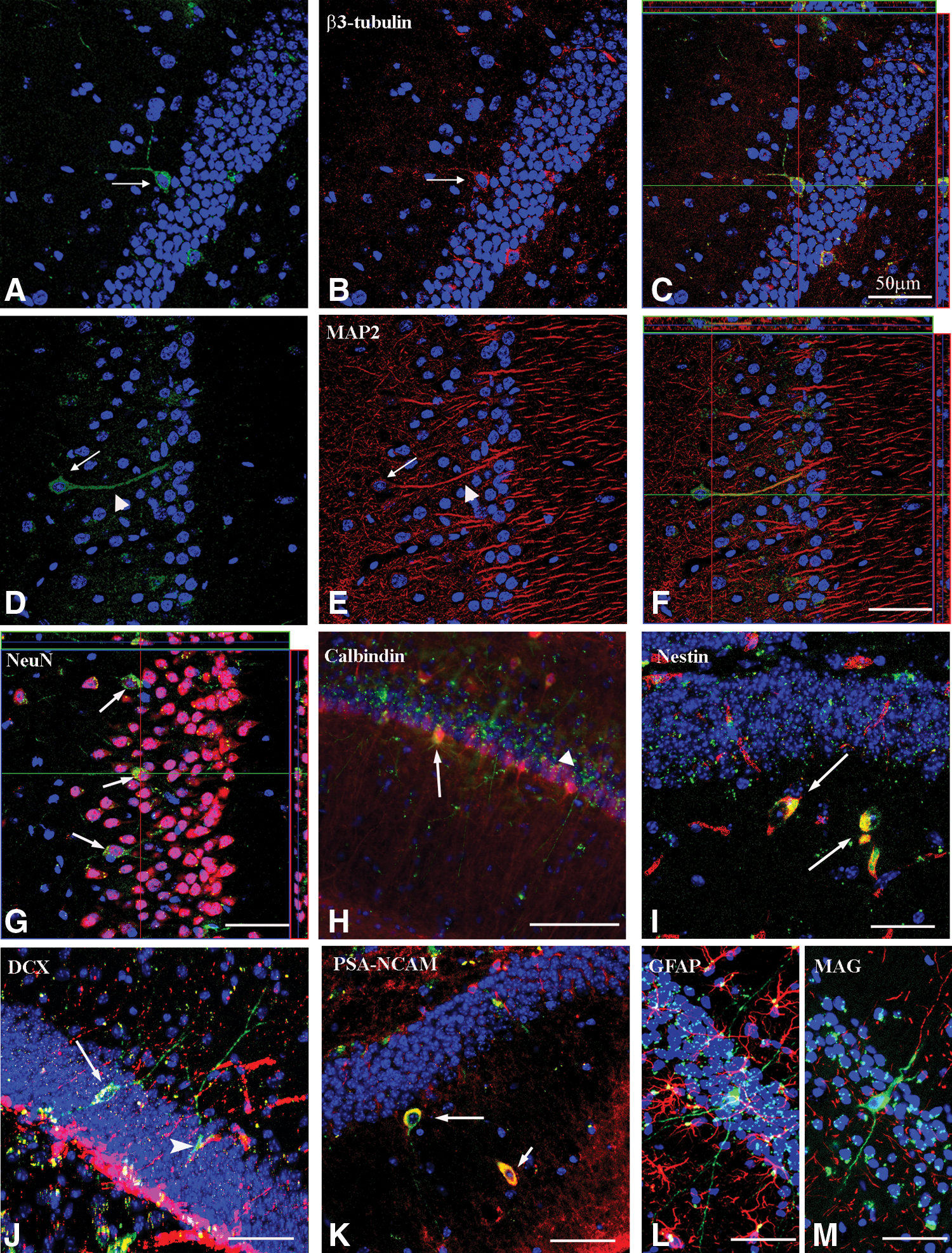
PAPCs can express stem cells as well as early and late neuronal differentiation markers.
Morphological maturation of eGFP cells in maternal brain
To study morphological maturation of PAPCs, maternal HC were collected and analyzed at P7 (n = 11), P60 (n = 5), and P120 (n = 7); and axonal/dendritic complexity of individual PAPCs was quantified (Fig. 4). At P7, we observed fewer total numbers of branches, and the majority of the processes were of lower branching order (Fig. 4A, C–E) as compared with P60 when PAPCs exhibited a higher axonal/dendritic complexity (Fig. 4B, C–E). We did not observe a major difference between PAPCs' axonal/dendritic complexity between P60 and P120 mothers, suggesting that PAPCs might mature and integrate into the mother's circuitry at about P60.

Axonal-dendritic maturation of PAPCs.
PAPCs do not fuse but differentiate into neurons
The expression of mature neuronal markers in PAPCs could result from cell fusion with mature maternal neurons. To exclude this possibility, we undertook a blastocyst transfer experiment (see Materials and Methods section) in which blastocysts derived from crosses between WT females and eGFP+ fathers (eGFP: fetal marker) were transferred to Rosa26-lacZ (β-galactosidase: maternal marker) foster mothers. The rationale was that in the case of cell fusion, PAPCs would express both genetic markers, whereas the absence of lacZ expression in eGFP+ PAPCs would rule out a fusion event. We screened for eGFP+ cells in ROSA26-lacZ recipient mothers, and eGFP+ cells were detected in 4 out of 10 Rosa26-lacZ mothers. This demonstrated that this experimental procedure led to PAPC transfer in a frequency comparable to normal pregnancies. As expected, β-galactosidase antibody staining was negative in WT females (not shown). Controls of Rosa-26-lacZ females showed the typical ROSA26-lacZ expression pattern (Fig. 5A), whereas eGFP+ animals stained with α-βGal did not show any lacZ staining (Fig. 5B). Further, eGFP:ROSA26-lacZ double transgenic animals demonstrated that lacZ expression could be found within the eGFP expression domain of eGFP-positive cells, highlighting that cells carrying both transgenic markers can be readily detected (Fig. 5C, arrows). We then examined 55 eGFP+ cells in transfer mothers by recording z-stack confocal images that we carefully analyzed for any coexpression of the markers in 3D. We found that 45 cells (82%) did not express the maternal marker β-galactosidase (Fig. 5D). Ten of the 55 cells (18%) showed lacZ staining in the periphery of the eGFP+ labeling. However, closer examination of the relevant z-stack recordings revealed that most of these β-galactosidase speckles did not overlap with eGFP and, therefore, β-galactosidase expression most likely was not within the domains of the eGFP expressing PAPCs. We conclude that PAPCs unlikely fused with maternal neurons within the HC and PAPCs acquired their neuronal cell fate by a process of differentiation.
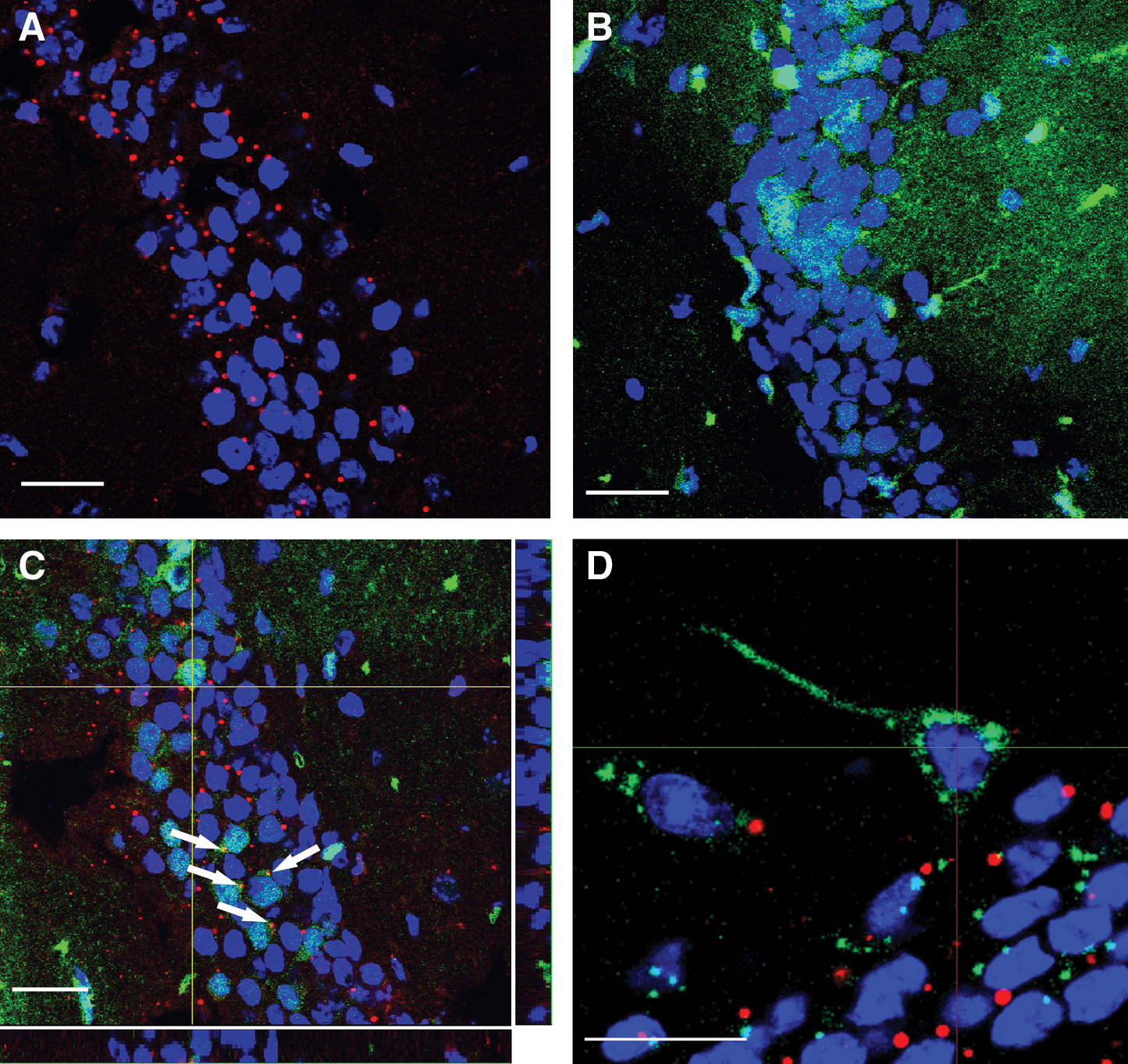
PAPCs do not fuse with maternal neurons.
Frequency and distribution of PAPCs are changed in HC of experimental PD animals
We generated experimental Parkinsonian mice via unilateral injection of 6-OHDA into the right striatum. This procedure results in the depletion of dopaminergic neurons within the treated hemisphere. Lesioned mice, which exhibited the typical behavioral phenotype (>5 rotations/min) on apomorphin injection, were mated with eGFP+ fathers. We analyzed the maternal HC, where robust PAPC integration and differentiation can be observed. eGFP+ PAPCs were generally found in P7 and P60 PD mothers (Fig. 6A, B). In nonlesioned control animals, 6 out of 9 mothers at P7 (66.7%) and 7 out of 9 mothers at P60 (77.8%) had PAPCs (total: 13/18 mothers, 72%). In lesioned animals, all P7 mothers (n = 4; 100%) and 4 out of 5 P60 mothers (80%) were positive (total: 8/9 mothers, 88%), suggesting that the experimental lesion increased the frequency of positive mothers. In histological sections of P7 nonlesioned mothers, an average of 12.7 ± 4.3 cells per hippocampal section was observed. A similar number (13.5 ± 4.2) was found in nonlesioned mice at P60. We did not find any difference between right and left HC at P7 (R: 6.2 ± 2.0; L: 6.5 ± 2.4) or right and left HC at P60 (R: 6.7 ± 2.0; L: 6.8 ± 2.1) (Fig. 6C). Thus, fetal cell frequency and distribution were similar in both P7 and P60 nonlesioned control mothers.
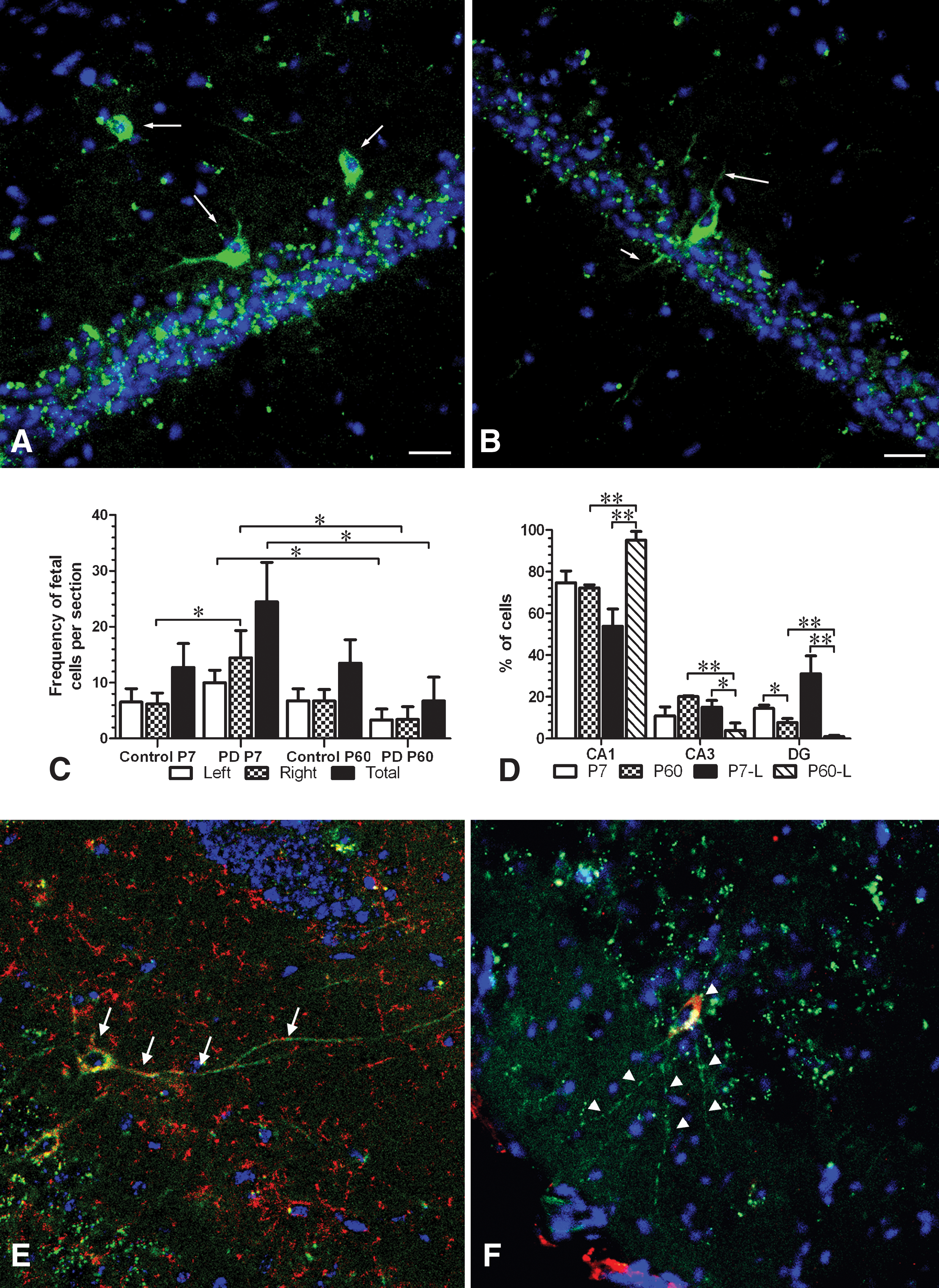
Fetal cell frequency and distribution in mothers with PD.
In lesioned mothers, the frequency of PAPCs was 24.5 ± 7.1 and 6.75 ± 4.2 at P7 and P60, respectively, which represents a statistically significant (P < 0.05) decrease. A significant decrease was also observed between the right HC of P7 and P60 (P7: 14.5 ± 4.9; P60: 3.4 ± 2.3) as well as the left HC of P7 and P60 mothers (P7: 10.0 ± 2.3; P60: 3.3 ± 2.0; Fig. 6C). Although more total PAPCs were detected in P7 lesioned mothers (24.5 ± 7.1) compared with nonlesioned mothers (12.7 ± 4.3), the difference was not significant. Likewise, the decrease in total PAPCs in lesioned mothers at P60 (6.8 ± 4.2) as compared with nonlesioned P60 mothers (13.5 ± 4.2) was also not significant. However, when comparing the right HC (the lesioned side) of P7 mothers to the right HC of nonlesioned mothers, numbers of PAPCs significantly increased by 2.3-fold (lesioned: 14.5 ± 4.9; nonlesioned: 6.2 ± 2.0; P < 0.05; Fig. 6C). The left HC of lesioned and nonlesioned mothers at both P7 and P60 were not significantly different. Taken together, our data demonstrated a significant increase in the initial recruitment of PAPCs into the right (lesioned) HC of P7 mothers as well as a significant decrease of PAPCs in both the left and right HC between P7 and P60 of lesioned animals.
Further, the overall distribution of PAPCs was changed in lesioned HC (Fig. 6D). In P7 mothers, the frequency of PAPCs in the CA1/2 area (53.9% ± 8.2%), the CA3 area (15.1% ± 3.2%), and the DG (31.1% ± 8.6%) seemed different as compared with nonlesioned mice (CA1/2, 74.7% ± 5.7%; CA3, 10.9% ± 4.3%; DG, 14.5% ± 1.6%); but these differences were not significant (P > 0.05) (Fig. 6D). However, in P60 lesioned mice, PAPCs were predominantly located to the CA1/2 area (95.2% ± 4.1%; CA3: 3.9% ± 3.5%, P < 0.05; DGL: 0.9% ± 0.6%, P < 0.05; Fig. 6D). This change in PAPC distribution is significant compared with nonlesioned mothers (CA1/2, 72.2% ± 1.5%, P < 0.01; CA3, 20.1% ± 0.4%, P < 0.01; DG, 7.7% ± 1.9%, P < 0.01; Fig. 6D). The change in PAPC distribution was also significantly different when compared with all 3 hippocampal areas in lesioned P7 mothers (CA1/2, P < 0.01; CA3, P < 0.05; DG, P < 0.01). Thus, besides the age-specific differences of PAPC frequency, we observed a net shift of PAPCs to the DG in P7 and to the CA1/2 area in P60 animals.
The axonal/dendritic complexity of PAPCs in mothers with PD at P7 was comparable to those found in nonlesioned brains. However, PAPCs in the lesioned brains of P60 mice had a tendency to be less complex than their healthy controls, although these changes were not statistically significant. In agreement with our previous findings, most of the PAPCs found in PD animals also differentiated toward the neuronal lineage, as they expressed neuronal-specific markers such as β3-tubulin and DCX (Fig. 6E and F, respectively).
Discussion
PAPCs in maternal brain
Cell trafficking between fetus and mother has been a long known phenomenon associated with clinical complications during human pregnancies [18,19]. However, it has been recently shown that PAPCs can engraft into a number of different maternal tissues [20,28] and can persist for >3 decades in the maternal blood [8,29]. Using a murine model system, we showed that PAPCs integrate into the brain of mothers with long-term engraftment capabilities of up to 7 months postpartum. PAPCs were identified in all 6 studied brain areas, suggesting that PAPCs have the intrinsic capability to widely integrate into the unmanipulated brain. Our findings might suggest that PAPCs are responsive to regional-specific attractive cues facilitating homing into different brain areas. Alternatively, fetal cell homing is stochastic and does not require any particular mechanism.
PAPCs are undetectable in the HC before or at delivery. This finding is consistent with a previous report which suggested that in average, fetal cells could not be found in the brain at prenatal stages [10]. We detected PAPCs at about the time of delivery in approximately one-sixth of the mothers (17%), whereas at P7 and later on PAPCs were consistently found in about half of the mothers or more (43%–78%). Although our observations somewhat differ from the study by Fujiki et al. [10], they are consistent with previous observations by Tan et al. [11]. However, neither of both reports analyzed the presence of fetal cell beyond P6/P7 [10] or P90 after delivery of the last litter of pups of ex-breeders with multiple pregnancies [11]. Therefore, our study provided first evidence for long-term engraftment of PAPCs in the murine brain. This long-term engraftment is consistent with another study reporting the long-term presence and survival of fetal cells [8] in humans.
The increase in detectable cells between P0 and P7 suggests that PAPCs in the brain increased postpartum, in the absence of fetal donors, but reached a stage of equilibrium at around P7. It is not known why PAPCs are not detected in the pre-partum brain although PAPCs are generally found in the peripheral circulation at this stage. Possible explanations could be that the maternal brain during pregnancy is inert to fetal cell invasion, maybe as a consequence of the hormonal status of the mother during pregnancy and a resulting relative robustness of the blood brain barrier. On the other hand, the delivery process and the physiological breakdown of the endothelial barrier between mother and fetus in the placenta could also result in a decreased efficiency of the blood brain barrier so that PAPCs could more easily penetrate the brain as a consequence. It is also tempting to speculate that the process of delivery could lead to microinjuries to the brain, leading to the release of cytokines recruiting PAPCs to the brain. Also, a surge of fetal cell infiltration to mothers is observed around the time of delivery; and it could well be that increased numbers of fetal cells in the brain are merely a reflection of an increased availability of PAPCs in the peripheral blood system. Interestingly, our observation of increasing cell numbers in the brain at P7 coincides with a general disappearance of PAPCs in the peripheral blood after delivery [10]. PAPCs could also be recruited from a yet unknown fetal cell pool(s) within the mother. It has been shown that maternal cells stably engraft into the fetal bone marrow in the murine systems for at least up to 10–12 weeks postpartum [6]. It is possible that PAPCs could be recruited from the known stem cell niche bone marrow in the mother to the brain. Also, PAPCs might proliferate in their respective locations before differentiation, although so far there is no direct evidence for clonal expansion of PAPCs in mothers.
It is not known how PAPCs transfer to the mothers, but the most intuitive route would be via the fetal cord blood-placenta-maternal blood axis [30]. This hypothesis finds some support in the presence of fetal cells in the maternal blood circulation at about E11-14. According to another report [10] as well in our own unpublished observation (data not shown), PAPCs are found in the blood at the same time or slightly before cells are detected in solid organs such as the lung, liver, and spleen. This might indicate that the blood is one of the earliest populated “organs.” However, it is unclear whether PAPCs found in the blood are relatively immature fetal progenitor cell types temporarily circulating in the blood during their passage to maternal target organs or whether they have already undergone some level of, for example, hematopoietic differentiation and, as a consequence, are an intrinsic part of the blood system.
Neural differentiation of PAPCs
In accordance with a published report [11], we found that the PAPCs express immature neuronal marker β3-tubulin as well as mature markers such as MAP2, NeuN, and calbindin, thus suggesting that PAPCs acquired a gene expression profile which is typical for neurons. With regard to calbindin expression, PAPCs in the HC adopted either a calbindin-positive or a calbindin-negative phenotype. Calbindin expressing cells are either principal cells or interneurons of the HC, of which about 10%–12% are estimated to be GABAergic interneurons [31]. Endogenous hippocampal neurons are also either positive or negative for calbindin. This suggests that PAPCs are capable of responding to local hippocampal differentiation cues in a manner similar to endogenous differentiating neurons. Interestingly, we found that PAPCs could express either the neural stem cell (NSC) marker nestin or immature neuronal markers such as DCX and PSA-NCAM. PAPCs expressed neuronal-specific marker genes as early as P7, which implies that they adopted an intermediate to a mature neuronal phenotype at this stage. PAPCs could have transferred to the mother as early as the blastocysts implant into the uterus at E4.5. However, PAPCs are only detectable in the mother's circulation at around E10–11 in quantifiable numbers [10]. This implies that PAPCs could be within the brain for a maximum period of 18.5–24 days (from either blastocyst implantation or E10 to about P7). The time frame of about 3 weeks is consistent with the maturation of endogenous NSCs into mature neurons, which normally requires between 14 and 28 days in which neural stem cells in the adult HC progress to neurons through developmental stages defined by the expression of specific marker proteins [32]. The early putative neural stem cell is characterized by the expression of Nestin and GFAP. Neuronal development proceeds by cells extinguishing nestin expression and by upregulating DCX, which labels a more lineage-determined putative progenitor cell. Around the same stage, cells also upregulate PSA-NCAM expression. PSA-NCAM is a marker commonly found on migratory early neurons. Such cells then withdraw from the cell cycle to become postmitotic. This is followed by cells starting to express early neuronal markers such as β3-tubulin. The stage of expression of immature markers also coincides with the development of network connections as an indication of integration into the pre-existing circuitry. Last, neurons mature into fully differentiated neurons, and this coincides with the expression of markers such as NeuN and calbindin. Due to technical limitations, we have not studied whether individual PAPCs convert from cells expressing a NSC marker to immature and then mature neurons. However, the presence of PAPCs with a stem cell or an immature neuronal phenotype is consistent with the notion that PAPCs transit from a neural stem-cell-like stage through an intermediate differentiation stage to fully matured neurons. Such transition has been described for adult neurogenesis [32]. Neuronal maturation of PAPCs is also supported by the observation of an increasing axonal/dendritic complexity between P7 and P60, although functional integration of PAPCs into the existing maternal circuitry still needs to be demonstrated.
Expression of glial markers by PAPCs could not be detected, indicating that in our experiments PAPCs predominantly adopted a neuronal fate. A previous study has reported glial differentiation of PAPCs [11]. One possible explanation for this discrepancy is that this study used healthy, unmanipulated animals; whereas Tan et al. examined a hippocampal acute lesion model particularly for the glial differentiation. The experimental lesion could have possibly led to changes in the local brain microenvironment to which PAPCs could have responded by producing glial cells under such conditions. It is well known that acute injury to the brain stimulates reactive astrogliosis [33], thus resulting in massive proliferation of resident astrocytes as well as in major morphological and biochemical changes in astrocyte physiology. It is conceivable that such mechanisms also could contribute to the de novo generation of glia from PAPCs.
Cell fusion between stem cells and differentiated cells has been described as a possible reason for expression of differentiation markers in stem cells; for example, for bone marrow mesenchymal stem cells (MSCs) [34 –38] and brain cells [39]. We explored whether PAPCs fused to maternal neurons, and we demonstrated that the majority of PAPCs with neuronal morphology did not express the maternal marker. This suggests that PAPCs did not fuse with maternal neurons. Also, we did not detect PAPCs with neuronal morphology or neuronal marker gene expression containing 2 distinct nuclei as an indication for heterokaryon formation. Our results are consistent with the notion that PAPCs differentiated into neurons rather than acquiring neuronal fate through fusion with preexisting maternal neurons.
PAPCs are initially attracted to the HC of PD mothers, but long-term integration is impaired
We investigated the frequency and distribution of PAPCs in a murine PD model system. Our study showed that frequency of PAPCs in the lesioned side of the brain is initially increased. This is consistent with the interpretation that the experimentally manipulated tissue attracted PAPCs. It has been reported that injury can attract PAPCs to the brain [11], liver [22], and skin [21]. Similarly, migratory properties to injury sites have been described for several stem cell types including neural stem cells (NSCs) [40], hematopoietic stem cells (HSCs) [41], and mesenchymal stem cells [42]. In some instances, the SDF-1/CXRX4 receptor axis has been linked to the chemotactic attraction of cells including stem cells [43] to injury sites. It can be speculated that similar chemokine-related mechanisms may play a role in the initial attraction of PAPCs to healthy and, particularly, to diseased tissues. Interestingly, some organs respond to tissue injury through upregulation of SDF-1 expression and release. This facilitates physiological responses such as motility, chemotactic responses, adhesion, and secretion of matrix metalloproteases of cells carrying the cognate receptor CXCR4 [44]. Such mechanisms could account for the attraction of PAPCs to injury sites, their capability of crossing endothelial barriers of the vasculature and homing to injured organs.
PAPCs were found in lower total frequency in P60 PD mothers, suggesting that the disease environment did not support their long-term maintenance or survival. This could be due to injury-related changes in the local microenvironment impairing on cell survival, possibly as a consequence of lack of maintenance/survival cues in these injury sites.
We also observed that PAPCs were differently distributed in the HC in lesioned and nonlesioned mothers. There were no major differences between the location of PAPCs in HC of P7 lesioned and nonlesioned mothers except for a trend of increasing numbers located in the DG. This is an interesting observation, because the DG is an endogenous stem cell niche within the adult brain. In contrast, at P60, we noticed significant changes in fetal cell distribution in the HC. Significantly lower numbers of PAPCs were found in the DG and the CA3 area, which was accompanied by a significant increase in cell numbers in the CA1/2 area as compared with P7 lesioned mothers and P60 nonlesioned mother. This shift could be due to either an active relocation of cells to the CA1/2 area or their relative enhanced survival. We tend to favor the latter, as PAPCs were found in significant lower numbers in older P60 mothers.
Experimental lesions had been performed in the striatal areas, resulting in the typical loss of DA neurons in the substantia nigra. It remains unknown why fetal cell attraction as well as long-term retention in the HC, a part of the brain normally not known to be affected in this disease model, is altered. Recently, 2 studies have reported host-to-transplant disease progression in PD in which transplanted fetal DA neurons have been shown to undergo similar pathologies as observed in native tissue of these patients [45,46]. Although the precise mechanism of disease progression in PD and propagation of disease to transplants require further elucidation, mechanisms such as inflammation, oxidative stress, and excitotoxicity as well as loss of neurotrophic support have been shown to play a role [47]. In addition, neurogenesis and consequentially endogenous recovery mechanisms of stem cell activation are negatively affected under experimental PD conditions [48]. Thus, the disease microenvironment might play a crucial role, for example, in differentiation and/or cell survival of endogenous stem cells as well as transplanted cells, which might impact the outcome of any cell-based regenerative strategy. More studies are required to understand microenvironmental changes of the diseased brain and how this might influence cell recruitment, survival, integration, and differentiation in the context of cellular therapies.
In summary, we showed that PAPCs engraft into the maternal brain and undergo neural differentiation in the absence of cell fusion. PAPCs can express stem cell markers as well as immature neuronal markers, suggesting that they might go through a similar cellular and molecular maturation process as is observed during adult neurogenesis. Also, PAPCs increase axonal-dendritic complexity over time, supporting the notion of a process of cell maturation and time-dependent integration into the brain. We suggest that the model of fetomaternal microchimerism in the brain can be used as a model to study cell homing, survival, and integration into the existing circuitry of the brain. Regenerating connectivity still represents a major hurdle particularly for cell therapies in neurodegenerative diseases. We suggest that the murine fetomaternal microchimerism model can be utilized as a noninvasive cell transplantation model, as it mimics an allogenic transplantation setting. Finally, PAPCs might constitute a novel class of stem or progenitor cells with tissue-specific homing and differentiation capabilities, which might be exploitable for cellular therapies in the future. In this context, the major hurdle remains the isolation, in vitro propagation, and differentiation of PAPCs into specific cell types for use in cell transplantations in animal disease models.
Footnotes
Acknowledgments
This work was supported by the Institute of Medical Biology and a joint grant from the Juvenile Diabetes Research Foundation, USA, and the Biomedical Research Council, Singapore. The authors thank Esther Wong, Xie Huijuan, and Manickam Arun Kumar of the Biological Resource Center (BRC) for the excellent technical assistance during the blastocyst transfer experiment.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.