
Abstract
We propose zinc fingers and homeoboxes 3 (ZHX3) as new osteogenic markers for mesenchymal stem cells (MSC). ZHX3 mRNA expression was upregulated within 1–6 h after incubation of MSCs in the osteogenic induction medium, and reached maximum levels after 24 h of incubation. Two to 4 days later, ZHX3 mRNA levels had decreased sharply. Maximal mRNA levels were 3- to 5-fold higher than those in the undifferentiated state. In contrast, Runt-related transcription factor2 (RUNX2) mRNA expression was downregulated at 2–4 h after incubation, and levels were only enhanced 1.4-fold after 12 and 24 h of incubation. Further, Osterix mRNA levels increased only 1.6-fold after 4 and 24 h of incubation. Thus, ZHX3 expression may be a better marker of MSC osteogenic differentiation than RUNX2 or Osterix expression at the initial stage of differentiation. Knockdown of ZHX3 using 2 distinct small interfering RNA (siRNA) oligonucleotides had little effect on cell morphology or on MSC proliferation, regardless of the differentiation state of the cells. However, ZHX3 siRNAs suppressed Osterix, but not RUNX2 mRNA expression, within 1 h of osteogenic differentiation, and this suppression was sustained for at least 24 h. The 2 ZHX3 siRNAs also suppressed alkaline phosphatase induction and matrix mineralization (assessed using alizarin red staining), and, further, suppressed the calcium content of the cultures at a later stage of differentiation (days 6–21). The effects of ZHX3 siRNAs on the osteogenic differentiation were comparable to those of RUNX2 and Osterix siRNAs. These findings suggest that ZHX3 is involved in the switch from the undifferentiated state of MSC to an osteogenic program, and that ZHX3 may be useful as an early osteogenic differentiation marker.
Introduction
R
Several transcription factors, including Runt-related transcription factor2 (RUNX2) [5], Osterix [6], msh homeobox1 (MSX1) [7], MSX2 [8], distal-less homeobox5 [9], Krox-20 homolog (Krox-20) [10], Sp3 transcription factor (Sp3) [11], and activating transcription factor4 [12], are involved, directly or indirectly, in bone formation or in the osteogenic differentiation of MSCs. Twist homolog1 (TWIST1) has been reported to suppress osteoblast differentiation [13]. Glucocorticoid-induced leucine zipper (GILZ) suppresses adipogenic differentiation and promotes osteogenic differentiation of MSCs [14,15]. Among these transcription factors, RUNX2 and Osterix play pivotal roles in the osteogenic differentiation of MSCs [16,17]. We thus compared the expression of RUNX2 and Osterix as standard genes. However, other transcription factors that are expressed in the initial stages of differentiation may also play a part in osteogenic differentiation of MSCs.
The zinc fingers and homeoboxes (ZHX) family of transcription factors consists of 3 members: ZHX1, ZHX2, and ZHX3 [18 –20]. All of these proteins contain 2 Cys2-Hys2-type zinc finger motifs and 5 homeodomains, are localized in the nuclei of cells, and act as ubiquitous transcription factors [18 –21]. ZHX3 consists of 956 amino acid residues, and this protein forms homodimers and heterodimers with ZHX1 [19]. Further, ZHX2 and ZHX3 repress the expression of cancer markers in normal hepatocytes [22], and ZHX family members regulate podocyte gene expression during the development of nephritic syndrome [23,24]. However, a role for ZHX3 in osteogenic differentiation has not been investigated.
We have identified novel MSC-specific genes using DNA microarray and real-time RT-polymerase chain reaction (PCR) analyses [25]. In such studies, we noted the upregulation of several transcription factors, including ZHX3, at the initial stage of MSC osteogenic differentiation. In this report, we address the questions of whether and how ZHX3 is involved in the osteogenic differentiation of MSCs.
Materials and Methods
Cell culture
Two different human bone marrow MSC populations from patients with nonbone related disease were obtained from the iliac crest according to a protocol approved by ethics authorities at Hiroshima University, and were maintained in monolayer cultures as described [2]. One MSC line was obtained from Cambrex Bio Science (Walkersville, MD). The MSC lines cultured from both of these sources retained multilineage differentiation potential and did not show any obvious differences in global expression profiles. However, the characteristics of these MSC lines were markedly different from those of differentiated MSCs [25]. These MSCs were seeded at a density of 5 × 103 cells/cm2 in 90-mm tissue culture dishes (BD Biosciences, Franklin Lakes, NJ) and were maintained in Dulbecco's modified Eagle's medium (DMEM; Sigma, St. Louis, MO) supplemented with 10% fetal bovine serum (FBS) and 1% antibiotic–antimycotic (Gibco BRL, Gaithersburg, MD) in the presence of 3 ng/mL fibroblast growth factor-2 (medium A) at 37°C in a humidified incubator in a 5% CO2/95% air atmosphere. Attached cells were provided with fresh medium A every 3 days. Cells were passaged when cultures became confluent, and cells obtained from the third to fifth passages were used for the following experiments.
Osteogenic differentiation
MSCs were seeded at a density of 5 × 103 cells/cm2 in tissue culture dishes and were provided with medium A until confluent. Thereafter, the cells were maintained in minimum essential medium alpha (Gibco BRL) supplemented with 10% FBS, 1% antibiotic–antimycotic, 100 nM dexamethasone (Sigma), 10 mM β-glycerophosphate (Sigma), and 50 μg/mL ascorbic acid-2-phosphate (Sigma; medium B). Cultures were provided with medium B every 3 days.
Adipogenic differentiation
MSCs were seeded at a density of 5 × 103 cells/cm2 in tissue culture dishes and were grown to confluence in medium A. Cells were treated with adipogenic induction medium (DMEM-high glucose containing 10% FBS, 1 μM dexamethasone, 0.2 mM indomethacin, 10 μg/mL insulin, and 0.5 mM 3-isobutyl-1-methylxanthine) for 3 days and were then switched to adipogenic maintenance medium (DMEM-high glucose containing 10% FBS plus 10 μg/mL insulin) for another 3 days.
Chondrogenic differentiation
MSCs were seeded at a density of 5 × 103 cells/cm2 in tissue culture dishes and were grown to confluence in medium A. Cells were treated with chondrogenic induction medium (minimum essential medium alpha containing 1 μL/mL ITS+ Premix, 100 nM dexamethasone, 50 μg/mL ascorbic acid-2-phosphate, 100 μg/mL sodium pyruvate, 3,500 μg/mL, and 10 ng/mL Recombinant Human Transforming Growth Factor-β3) every 3 days, and were then cultured with monolayer [26].
RNA isolation
Total RNA was isolated from cells using the RNeasy Mini kit (Qiagen, Tokyo, Japan) according to the manufacturer's instructions.
Quantitative real-time reverse transcription-polymerase chain reaction
First-strand cDNA was synthesized from 1 μg of total RNA using the High Capacity cDNA Reverse Transcription kit (Applied Biosystems, Foster City, CA). Real-time PCR was performed using the Universal ProbeLibrary System (Roche Diagnostics, Manheim, Germany) and the LightCycler 480 (Roche Diagnostics). Primer sets were designed using ProbeFinder version 2.45 for Human (Roche Diagnostics) as follows: ZHX3 left primer, 5′-gagccctttgacacatcgag-3′; right primer, 5′-tcagatcaaattcagtctgtttcg-3′; probe number, 64 (Universal ProbeLibrary System). RUNX2 and Osterix primers and probes were designed using DNASIS® version 2.08 (Hitachi Software Engineering Co., Ltd., Tokyo, Japan): RUNX2 left primer, 5′-ctccaacccacgaatgcacta-3′; right primer, 5′-cggacataccgagggacatg-3′; and probe 5′-ccacctttacttacaccccgccagtcac-3′; and Osterix left primer, 5′-atgagctggagcgtcatgtg-3′; right primer, 5′-aggtggtcgcttcgggtaa-3′; and probe 5′-tcacctgcctgctctgctccaagc-3′. In pilot studies during MSC osteogenesis, we found that peptidylprolyl isomerase A mRNA expression is much more stable than GAPDH or β-actin mRNA expression, which are commonly used as internal controls. This has also been reported previously [27]. mRNA signals were therefore normalized using peptidylprolyl isomerase A as an internal control (Roche Diagnostics).
RNA interference
The small interfering RNAs (siRNAs) used in this study were designed as described by Ui-Tei and others and Reynolds et al. [28 –31], and were synthesized by RNAi Co., Ltd. (Tokyo, Japan). Four siRNAs were designed for ZHX3, RUNX2, and Osterix. These siRNAs were analyzed with regard to their efficiency of knockdown of ZHX3, RUNX2, and Osterix mRNAs, respectively, in MSCs. Based on this analysis, the following effective siRNAs were selected for use in experiments: siZHX3-1, 5′-cccauuaacacuguguguuca and 5′-aacacacaguguuaaugggug; siZHX3-2, 5′-ccaugacaugacccaauuugu and 5′-aaauugggucaugucauggga; siRUNX2, 5′-ccagccaccuuuacuuacacc and 5′-uguaaguaaagguggcuggau; siOsterix, 5′-cuagaucuggugccuagaagc and 5′-uucuaggcaccagaucuagca. Silencer Select Negative Control #2 siRNA (obtained from Applied Biosystems) was used as a control siRNA (siNC). MSCs were seeded at a density of 5 × 104 cells/well in 12-well tissue culture plates (BD Biosciences) for assay of cell number, and at a density of 9 × 104 cells/well in 12-well tissue culture plates (BD Biosciences) for other experiments. The siRNAs were transfected into MSCs when the cultures reached 50% or 70% confluence, using Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. To achieve efficient transfection, cultures were incubated with the siRNA transfection medium for 24 h. Thereafter, the cultures were provided with medium A for 2 days. When the cultures became confluent, cells were then exposed to the osteogenic induction medium.
DNA quantification
Cells were washed twice with physiological saline, and were disrupted on ice using a Vibra-cell (Sonics & Materials Inc., Danbury, CT) for 10 s in a solution containing 350 mM 2-amino-2-methyl-1-propanol (Sigma), 1 mM zinc sulfate (Wako Pure Chemical Industries, Ltd., Osaka, Japan), 2 mM magnesium sulfate (Katayama Chemical Industries, Co., Ltd., Osaka, Japan), and 2 mM ethylenediaminetetraacetic acid (Sigma; pH 10.4). Lysed cells were centrifuged at 12,000 g for 5 min. Supernatant was mixed with PicoGreen Reagent (PicoGreen dsDNA Quantitation kit; Molecular Probes, Boston, MA), and absorption was measured using a multilabel counter (Arvo-SX; PerkinElmer, Inc., Wellesley, MA) at 480 nm excitation and 520 nm emission wavelengths.
In pilot studies, cell number and DNA of similar cultures were simultaneously determined using a Particle counter (Z1 Coulter; Beckman Coulter K. K., Tokyo, Japan). The amount of DNA per MSC (7.85 pg/cell) was estimated and this value was used to calculate cell number.
Alkaline phosphatase activity
Supernatants of lysed cells were obtained as described above. Supernatant was incubated with 16 mM PNPP (SIGMA FAST p-nitrophenyl phosphate tablet sets) (Sigma), for 10–30 min at 37°C, and the reaction was then stopped by the addition of 1 N NaOH. The absorbance of the solution at 405 nm was measured using a spectrophotometric microplate reader (Immuno Mini NJ-2300; Nalge Nunc International, Rochester, NY). Alkaline phosphatase (ALP) activity measured at 4, 6, and 8 days was normalized against DNA content in the same well, and is expressed as ALP activity/min/ng DNA.
Alizarin red staining
Cell layers were washed twice with a solution containing 10 mM Tris-HCl, pH 7.6, and 0.9% NaCl, and were then fixed in 95% ethanol at room temperature for 1 h. Cell layers were stained with 1% Alizarin-red S (Chroma-Gesellschaft Schmid GMBH&Co., Kongen, Germany) for 5 min at room temperature in the dark. Alizarin red staining was performed at day 21 after induction of osteogenic differentiation.
Calcium content
Cells and extracellular matrix were washed twice with physiological saline, 10% trichloroacetic acid (Katayama Chemical Industries, Co., Ltd.) was then added to the cell layer, and the cells were disrupted using a Vibra-cell for 10 s. Lysed cells were centrifuged at 12,000 g for 5 min to obtain supernatant. Calcium content was determined using a Ca C-test Wako kit (Wako Pure Chemical Industries, Ltd.) and a spectrophotometric microplate reader (Immuno Mini NJ-2300) at 570 nm. Calcium content was quantified at day 21 after induction of osteogenic differentiation. Calcium content is expressed as Ca content/ng DNA. DNA content was quantified using Pico green.
Results
ZHX3 mRNA expression during osteogenic differentiation
To determine the role of the transcription factor ZHX3 in osteogenic differentiation, we analyzed ZHX3 mRNA expression during the early stages of MSC differentiation. Levels of ZHX3 mRNA expression were then compared to those of Osterix and RUNX2, transcription factors known to be involved in osteogenesis. MSCs from 3 different donors were separately incubated with osteogenic induction media, and ZHX3 mRNA levels were assayed by quantitative real-time reverse transcription-polymerase chain reaction (RT-PCR). At 1, 3, 6, 9, 12, and 24 h and 2, 4, 7, 14, and 28 days after osteogenic induction, total RNA was extracted for the analysis of ZHX3 mRNA expression, and at 1, 2, 4, 12, and 24 h after osteogenic induction, total RNA was extracted for analysis of RUNX2 and Osterix mRNA expression. In all cultures, ZHX3 mRNA levels increased within 1–6 h of induction and reached a maximum at 24 h (Fig. 1A). Maximal levels of ZHX3 mRNA expression were 3- to 5-fold higher than the levels in undifferentiated MSCs (Fig. 1B). Thereafter, ZHX3 mRNA levels decreased sharply until day 2, and low expression levels were then maintained until day 28. In contrast, Osterix mRNA levels only slightly increased over the first 24 h, whereas RUNX2 mRNA levels gradually decreased over the first 4 h and then gradually increased between 12 and 24 h after osteogenic induction. However, the changes in expression of Osterix and RUNX2 mRNA were much less than those observed for ZHX3 mRNA expression (Fig. 1C). Much stronger upregulation of RUNX2 and Osterix mRNA was observed several days after osteogenic induction (data not shown).
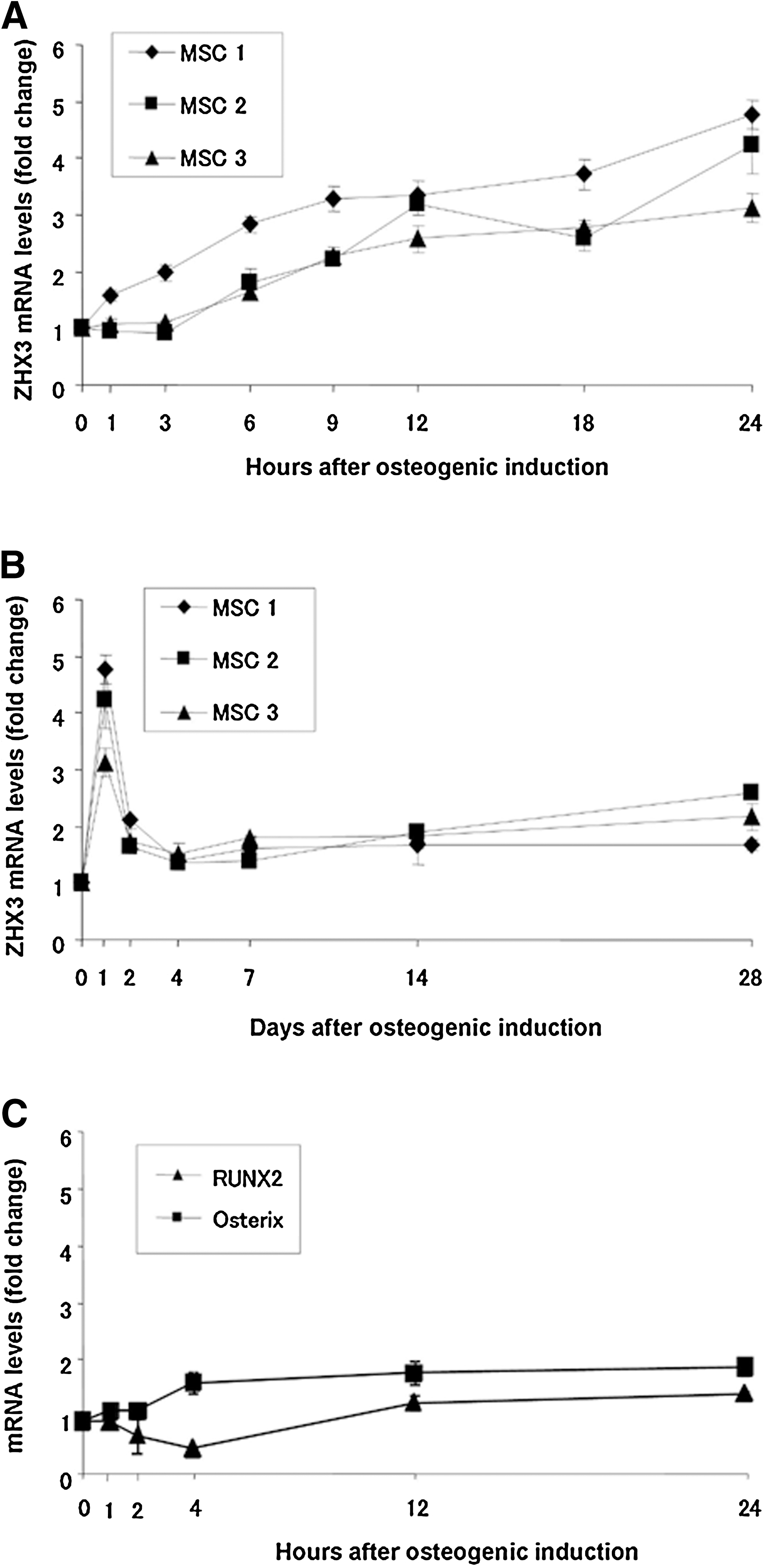
Zinc fingers and homeoboxes 3 (ZHX3), Runt-related transcription factor2 (RUNX2), and Osterix mRNA expression in bone-marrow-derived mesenchymal stem cells (MSCs) during osteogenic differentiation. Osteogenic induction was initiated after the indicated MSC lines (MSC1, 2, and 3) reached confluence. Total RNA was extracted at the indicated times and was used to analyze the expression levels of ZHX3, RUNX2, and Osterix mRNA using quantitative real-time reverse transcription-polymerase chain reaction (RT-PCR). ZHX3 mRNA expression was analyzed over 24 h
ZHX3 mRNA expression during chondrogenic or adipogenic differentiation
To determine whether upregulation of ZHX3 mRNA is specific for osteogenic differentiation, we also assayed ZHX3 mRNA levels in the 3 MSC lines following incubation with adipogenic or chondrogenic induction medium. At 1, 3, 12, and 24 h after adipogenic or chondrogenic induction, total RNA was extracted. No obvious changes in ZHX3 mRNA expression were observed over the first 24 h of chondrogenic and adipogenic induction (Fig. 2A, B).

ZHX3 mRNA expression during adipogenic
Effects of ZHX3, RUNX2, and Osterix knockdown using siRNAs on cell morphology and cell proliferation
We next generated siRNAs against ZHX3, RUNX2, and Osterix to further analyze their roles during osteogenic differentiation. The siRNAs used (siZHX3-1, siZHX3-2, siRUNX2, and siOsterix) decreased the mRNA levels of their respective target mRNAs in MSC to similar extents (∼50%), as assessed by quantitative real-time RT-PCR. This mRNA suppression was sustained for at least 4–7 days (Fig. 3). Knockdown of ZHX3, RUNX2, and Osterix mRNA using these siRNAs had little effect on cell morphology (Fig. 4A–K), and on cell proliferation (Fig. 4L, M) on day 3 after transfection when compared with control siRNA transfection on day 3, or to cells on day 0, indicating that neither ZHX3 nor RUNX2, Osterix do not affect MSC proliferation in undifferentiated and osteogenic conditions.
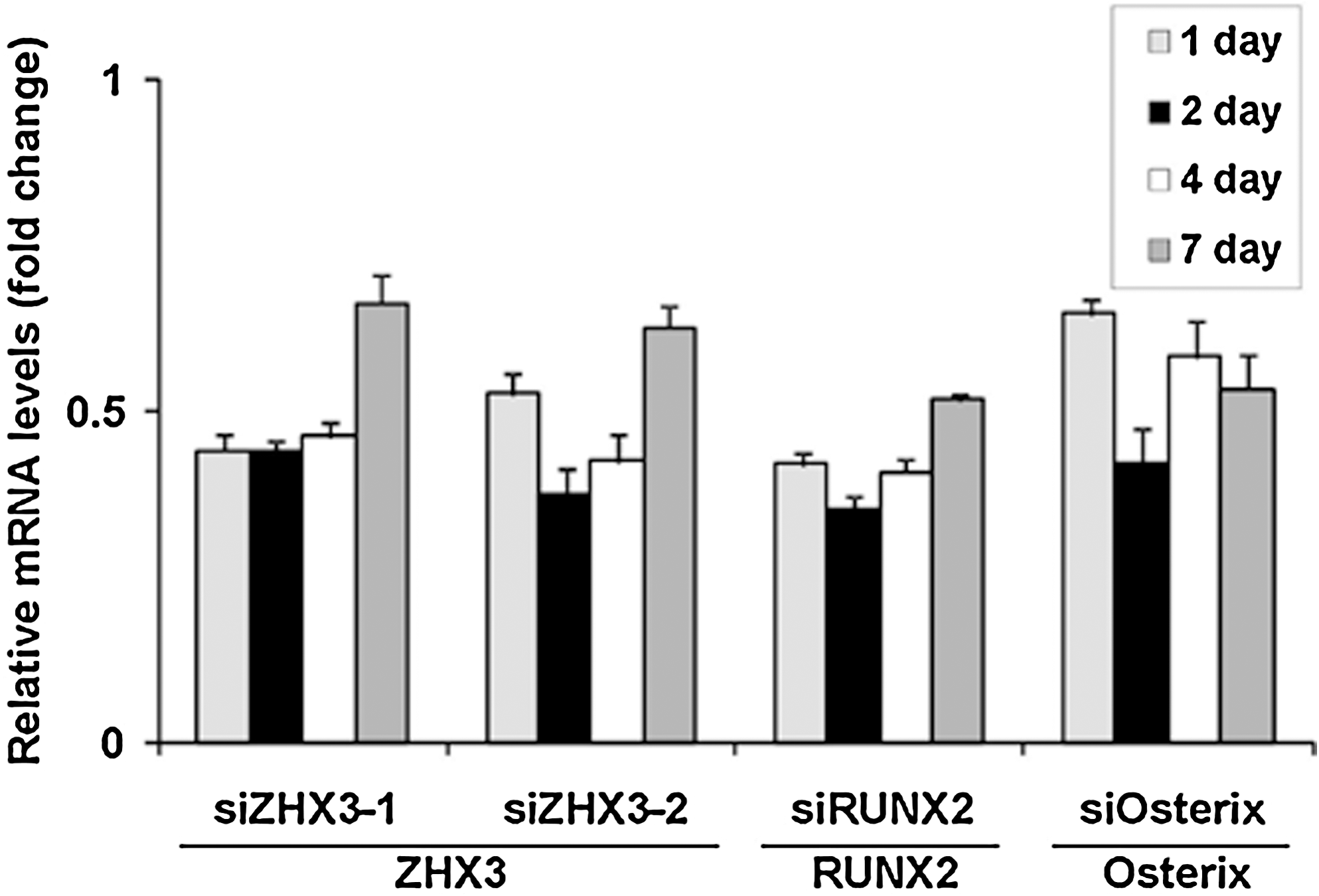
Knockdown efficiency of the small interfering RNA (siRNA) oligonucleotides. MSC cultures at 70% confluence were transfected with siZHX3 (siZHX3-1 or siZHX3-2), with siRUNX2, with siOsterix, or with the siNC. Cultures were incubated in siRNA transfection medium for 24 h after transfection. Thereafter, the cultures were provided with medium A at 1, 2, 4, and 7 days after transfection, total RNA was extracted, and ZHX3, RUNX2, and Osterix mRNA levels were determined by real-time PCR. The Y-axis shows the fold change in mRNA expression levels relative to their expression in siNC-transfected undifferentiated MSCs. Values represent the means ± SD of triplicate cultures.

Effects of ZHX3 and RUNX2 siRNA oligonucleotides on the morphology and proliferation of MSCs. MSC cultures at 50% confluence were transfected with siZHX3-1 or 2, with siRUNX2, with siOsterix, or with the siNC. Cultures were then incubated in siRNA transfection medium for 24 h. Thereafter, cells were transferred into medium A
Effects of siRNAs on RUNX2 and Osterix expression
We then tested the effects of ZHX3 siRNAs on the expression of RUNX2 and Osterix mRNAs, which are known to be induced during osteogenic differentiation of MSCs. MSCs were transfected with ZHX3, RUNX2, or Osterix siRNAs 3 days before incubation with osteogenic induction medium (∼70% confluence), and RUNX2 and Osterix mRNA levels were then determined using quantitative real-time RT-PCR. ZHX3 or Osterix knockdown using siZHX3-1, siZHX3-2, or siOsterix had little effect on RUNX2 mRNA expression throughout the experimental period (24 h), whereas siRUNX2 decreased RUNX2 mRNA levels by ∼50% at each time point following differentiation, as compared to control siRNA (Fig. 5A). However, both siZHX3-1 and siZHX3-2, as well as siRUNX2 and siOsterix, decreased the induction of Osterix mRNA expression at each time point (1–24 h), as compared to control siRNA (Fig. 5B).
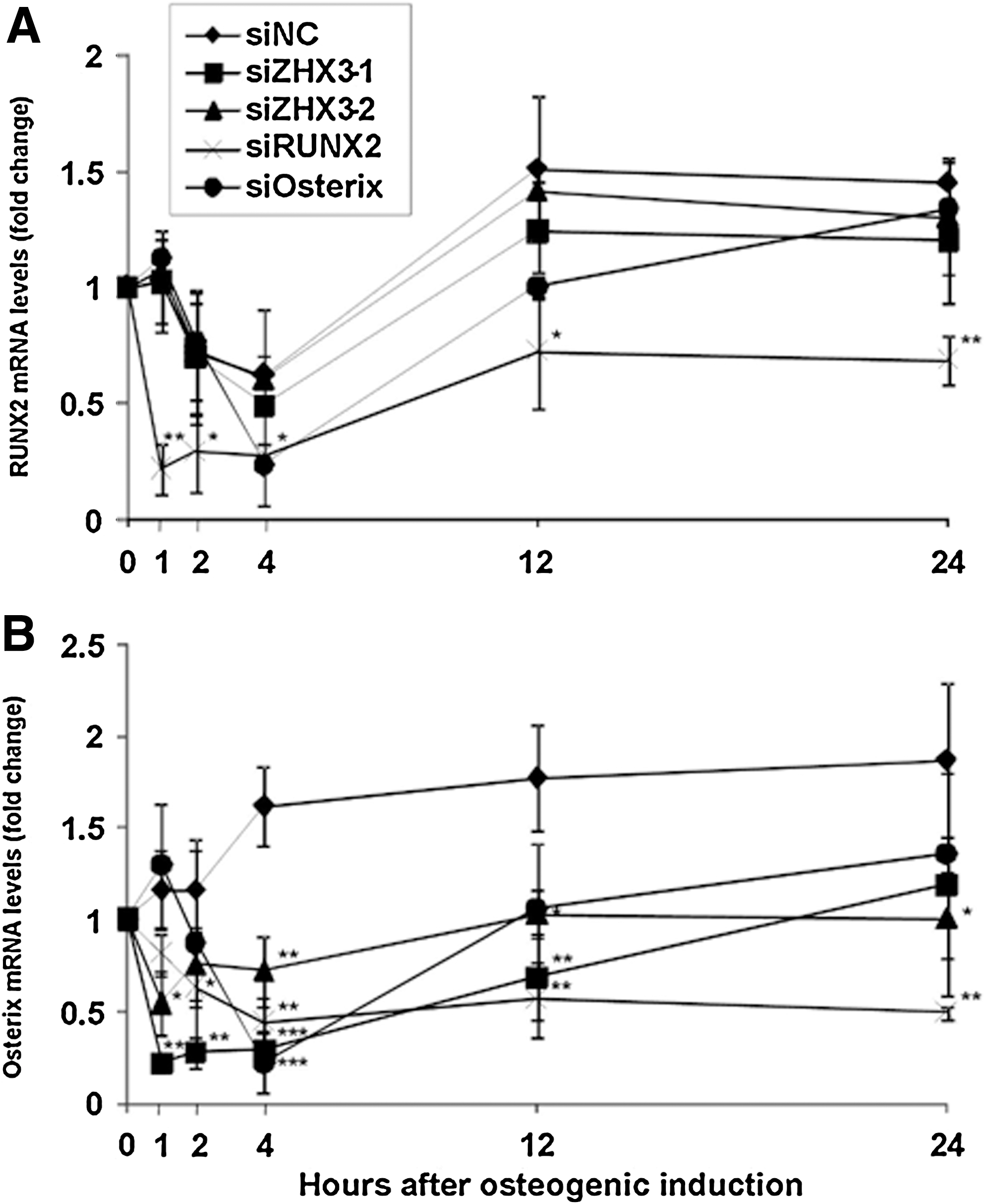
Effects of ZHX3, RUNX2, and Osterix siRNA oligonucleotides on RUNX2 and Osterix mRNA expression during the initial stage of osteogenic differentiation. MSC cultures at 70% confluence were transfected with siZHX3-1 or -2, siRUNX2, siOsterix, or with the siNC. Cultures were incubated in the siRNA transfection medium for 24 h, followed by medium A for 2 days. After the cultures became confluent, cells were transferred into the osteogenic induction medium. RUNX2
Effects of ZHX3, RUNX2, and Osterix siRNAs on ALP activity and calcification of MSCs
We finally assayed the effects of ZHX3, RUNX2, and Osterix siRNA transfection on ALP activity, matrix calcification, and calcium content of the cultures, which are all markers of osteogenic differentiation. The effects of transfection of siZHX3-1, siZHX3-2, siRUNX2, or siOsterix into MSCs on the activity of ALP were measured at 4, 6, and 8 days after osteogenic induction using a colorimetric assay. All ZHX3, RUNX2, and Osterix siRNAs tested suppressed ALP activity on days 6 and 8, at the midstage of osteogenic differentiation, as compared to control siRNA (Fig. 6A). Matrix calcification was determined using alizarin red staining and calcium content at 21 days after osteogenic differentiation (Fig. 6B). Both assays indicated that cultures transfected with ZHX3, RUNX2, or Osterix siRNAs had lower calcium content on day 21, which is a late stage of osteogenic differentiation, as compared to control siRNA-transfected cultures.

Effects of ZHX3, RUNX2, and Osterix siRNA oligonucleotides on osteogenic differentiation. MSC cultures were transfected with the indicated siRNAs and were maintained as described in the legend to Fig. 5. On days 4, 6, and 8 after the onset of osteogenic differentiation, alkaline phosphatase (ALP) activity was quantified using a colorimetric assay and was expressed as ALP activity/min/ng DNA
Discussion
Differentiation of MSCs into osteoblasts is usually induced by osteogenic induction medium containing dexamethasone, ascorbic acid, and β-glycerophosphate. However, dexamethasone is also a component of chondrogenic and adipogenic induction media, and ascorbic acid is a component of chondrogenic induction medium. Therefore, osteogenic induction medium may also upregulate genes in cells of other lineages. For example, GILZ, which is induced by dexamethasone, was strongly upregulated by chondrogenic, adipogenic, and osteogenic induction media on days 1 and 28 after induction, respectively (data not shown). It has been reported that GILZ overexpression induces MSC osteogenic differentiation, but reciprocally reduces adipogenic differentiation [15]. It has been reported that RUNX2, which is induced by the osteogenic medium, is essential for chondrogenic and osteogenic maturation of MSCs. However, RUNX2 has a role in eliminating the potential of MSC to differentiate into chondrocytes from osteoblasts [16]. Identification of transcription factors that are specifically upregulated during osteogenesis is important for understanding how cell lineages are regulated. In this study, we identified the transcription factor gene ZHX3, as being upregulated during osteogenic differentiation of MSCs, but not upregulated during chondrogenic or adipogenic differentiation, and as playing an important role in osteogenic differentiation.
Several transcription factor genes, including the gene encoding ZHX3, were selected in an earlier gene filtering study as genes that are induced during osteogenic differentiation, but not during chondrogenic or adipogenic differentiation. Among the well-known transcriptional factors that are reportedly involved in osteogenic differentiation, Osterix and Sp3 were upregulated by all differentiation induction media at an early stage. Distal-less homeobox5 was strongly upregulated by the chondrogenic induction medium at a late stage and was weakly upregulated by the osteogenic induction medium at both stages. MSX1 was upregulated by chondrogenic and osteogenic induction media at both stages. Activating transcription factor4 was strongly upregulated by chondrogenic and adipogenic induction medium at an early stage. Krox-20 was upregulated by osteogenic and chondrogenic induction media at a late stage, and TWIST1 was upregulated by osteogenic, chondrogenic, and adipogenic induction media at an early stage. We were not able to detect any obvious changes in MSX2 expression with any induction media at these time points (microarray data, data not shown). Of the transcription factors identified, ZHX3 was consistently and robustly induced after 24 h in all MSC lines exposed to the osteogenic induction medium. This suggests that ZHX3 is a novel marker gene for the osteoblast lineage. However, it is unclear why ZHX3 is not upregulated during chondrogenic or adipogenic induction, whose medium also contains dexamethasone, or dexamethasone plus ascorbic acid, respectively. It is possible that ZHX3 upregulation might be inhibited by other medium components in chondrogenic or adipogenic induction media. A second possibility is that β-glycerophosphate, which was present only in the osteogenic induction medium, may induce ZHX3 expression.
Our data suggest that ZHX3 plays a role in the early stages of osteogenic differentiation. A time-course study showed that ZHX3 is transiently upregulated within 24 h after the onset of osteogenic differentiation. ZHX3 expression was then rapidly downregulated within 2–4 days. One potential role for ZHX3 during early osteogenic differentiation may be to regulate the expression of the RUNX2 and/or Osterix gene, which have been shown to be essential for osteoblast differentiation. We showed that knockdown of ZHX3 suppressed the expression of Osterix but not the expression of RUNX2 (Fig. 5). It has been reported that Osterix is downstream of RUNX2 in mesenchymal cells [17]; thus, ZHX3 may act later than RUNX2 and may be upstream of Osterix in the MSC osteogenic process. It remains unclear whether ZHX3 acts directly on osteogenic differentiation or indirectly through Osterix, or both. To clarify the role of ZHX3 during MSC osteogenic differentiation, further investigation is needed. However, ZHX3 may not be involved in the maintenance or further maturation of osteoblasts in later stages, because upregulation of ZHX3 appears to be limited to a sharp peak in the first 24 h of MSC osteogenic differentiation. The observed suppression of the late stage of MSC osteogenic differentiation by ZHX3 siRNA may be caused by siRNA disruption of the early stage of differentiation, which may result in a delay in the sequence of osteogenic differentiation.
In this study, we used siRNAs to knockdown ZHX3, RUNX2, and Osterix gene expression. The knockdown efficiencies of the siRNAs were ∼50% and mRNAs were maintained at this suppressed level for 4 days after transfection, but began to increase again on day 7 after transfection (Fig. 3). One limitation of this study was that, as we were unable to transfect siRNA into confluent cultures, we had to transfect siRNA into cells that were ∼70% confluent. This meant that we had to wait 3 days after transfection for the cultures to become confluent before they could be transferred into the osteogenic induction medium. This protocol for siRNA transfection meant that knockdown at high efficiency could only be maintained 1–4 days after osteogenic induction. Nevertheless, this transient knockdown did suppress MSC osteogenic differentiation at later stages of differentiation (days 6–21) (Fig. 6). As the siRNAs used in this study had little effect on cell shape or cell proliferation, their observed suppression on Osterix expression, ALP activity, and calcification is not likely to be due to nonspecific actions of siRNA. Further, control siRNA did not modulate osteogenic differentiation. These findings strongly suggest that ZHX3 is essential for the switch from a nondifferentiated state to an osteogenic differentiation state. However, further studies using ZHX3 knockout or transgenic mice are necessary to understand the physiological role of ZHX3 in osteogenesis.
In summary, we suggest that ZHX3 is a novel and useful functional marker for MSC osteogenic differentiation. As most transcription factors that are upregulated after osteogenic differentiation of MSCs, including RUNX2 and Osterix, are upregulated <2-fold, ZHX3, which is upregulated by 3–5-fold, may be useful for predicting the osteogenic potential of individual MSC lines within the first 24 h of differentiation.
Footnotes
Acknowledgments
This work was supported in part by the Ministry of Education, Culture, Sports, Science, and Technology of Japan (Grants 19390497, 21791939, and 22390367) and by a Grant-in-Aid for Scientific Research from Nagasaki University, Japan.
Author Disclosure Statement
No competing financial interests exist.