
Abstract
Lesions to the fimbria fornix (FiFx) plus cingulate bundle (CB), the principal routes of communication of forebrain cholinergic regions, produce lasting impairment of spatial learning and memory in mice. We report that extensive neurogenesis takes place in the FiFx, CB, and basalis magnocellularis following FiFx plus CB transection. Immunofluorescence revealed that nestin-expressing cells were present in all 3 areas following lesion; the majority of nestin-positive cells were also positive for 5-bromo-2-deoxy-uridine, a marker of DNA synthesis. Nestin-positive proliferative cells were almost entirely absent from unlesioned tissue. Neurospheres cultured in vitro from lesioned FiFx displayed the characteristics of neural stem cells—proliferation, expression of embryonic markers, and multipotential differentiation into neurons, astrocytes, and oligodendrocytes. At early stages after transection, a small number of immature and migrating doublecortin-immunopositive neurons were detected in lesioned FiFx, where neuronal cell bodies are normally absent. At later stages, postlesion immature neurons developed into β-tubulin III–positive mature neurons. Lentivirus labeling assay implied that the injury-induced neurogenesis in FiFx may be from local neurogenic astrocytes but not from dentate gyrus. These results demonstrate that insult to cholinergic tracts can stimulate neural stem cell proliferation and neuronal regeneration not only in innervated regions but also in the projection pathways themselves. Ectopic neurogenesis in cholinergic system-related areas provides an additional mechanism for repair of cholinergic innervation following damage.
Introduction
N
The cingulate bundle (CB) and the fimbria fornix (FiFx) are the main routes traversed by forebrain cholinergic projections that innervate the hippocampus and neocortex from the septal/diagonal band complex and nucleus basalis of Meynert (NBM) in the mammalian brain [25 –29]. Lesions of the FiFx plus CB, therefore, markedly reduce cholinergic innervation of the hippocampus and cortex [29] and induce lasting impairment of spatial learning and memory [30]. In previous studies, we have demonstrated that neurogenesis takes place in denervated hippocampus following transection of the FiFx and CB. The environment of the deafferented hippocampus was found to stimulate neuronal and cholinergic differentiation of either endogenous or exogenous NSCs, which then migrated into the granular cell layer of DG [31,32]. However, the possibility that such regenerative processes might take place within the cholinergic projections to the hippocampus and cortex has not been addressed previously.
In the present study, we report that, in response to FiFx plus CB lesion, robust neurogenesis takes place in the FiFx, the CB, and the basalis magnocellularis (MBN), the rodent homolog of the human NBM. Proliferating NSCs were positive for 5-bromo-2-deoxy-uridine (BrdU) incorporation and nestin expression and were able to differentiate into neurons, astrocytes, and oligodendrocytes in vitro. At early stages after transection, immature and migrating neurons positive for doublecortin (DCX) were found in the lesioned FiFx, where prior to lesion there are only neuronal fibers and no cell bodies. At later stages, the immature neurons became mature neurons positive for β-tubulin III (BT3). Ectopic neurogenesis in the cholinergic pathway induced by nerve tract lesion, therefore, provides a new mechanism for regeneration of cholinergic innervation following damage. Lentivirus labeling assay implied that the injury-induced neurogenesis in FiFx may be from local neurogenic astrocytes but not from DG. These findings argue that manipulation of endogenous neurogenic cells is a potential therapeutic strategy for brain repair in conditions such as Alzheimer disease (AD), which are characterized by loss of cholinergic innervation.
Materials and Methods
Animal and surgery
Adult female Sprague–Dawley rats (220–250 g) were purchased from the experimental animal center of Nantong University. The animals were maintained in a temperature-controlled environment (23°C ± 2°C) on a 12:12-h light:dark cycle in an approved facility with free access to food and water. All animal experiments were conducted according to protocols approved by the United States National Institutes of Health Guide for the Care and Use of Laboratory Animals. All efforts were made to minimize the number and suffering of animals used in this study.
Unilateral FiFx lesions were performed by an adaptation of the method of Hefti [33]. Briefly, following intraperitoneal (i.p.) Chlorpent anesthesia (2 mL/kg body weight), a specially designed knife was lowered into left hemisphere of brain (antero-posterior [AP], −1.3; medio-lateral [ML], 1.0; and dorso-ventral [DV], 4.4) according to the atlas of Paxinos and Watson [34], moved laterally to 4.0 mm lateral to the midline and retrieved at this position. Nissl staining and acetylcholinesterase histochemistry [35] were used to verify the extent of the lesions (data not shown; referenced in our previous study [31]).
To track the migration of DG cells, lentivirus subcloned with GFP-coding sequence (provided by Biomics Biotechnologies Co.) was injected into bilateral DG at 3 days before FiFx lesions. Coordinates of injections according to the atlas of Paxinos and Watson [34] were 1.0 mm lateral to midline, 3.0 mm posterior to bregma, and 4.5 mm inferior to upper surface of skull bone. Each side received 1 μL GFP lentivirus (5 × 108 TU/mL).
Cell culture
At days 3 and 7 following left FiFx lesion, the contralateral and lesioned FiFxs behind the transection sites were carefully dissected under an anatomic microscope. Tissues were mechanically dissociated into single-cell suspensions following 0.25% trypsin digestion. The cell number and viability were determined by staining a small volume of cell suspension with 0.4% trypan blue. The cell suspensions were then maintained in NSC medium comprising Dulbecco's modified Eagle's medium (DMEM) and Ham's F12 medium (1:1, v/v) supplemented with 2% B27, 10 ng/mL epidermal growth factor, and 10 ng/mL basic fibroblast growth factor (bFGF). The cells were plated into flasks and incubated at 37°C in a 95% air/5% CO2 humidified atmosphere incubator for 12 h. When a proportion of the cells adhered to the flask surface, the remaining suspended cells were transferred to a new flask followed by incubation in NSC medium as above. Cultures were passaged every 7–8 days by dissociation of bulk neurospheres with Accutase (Innovative Cell Technologies).
The astrocyte culture was performed according to Zhang et al. [36]. Briefly, the cell suspensions from lesion and normal FiFx were prepared as above and plated in DMEM/F12 (1:1, v/v) containing 10% fetal bovine serum (FBS). After 7 days in culture, the flasks were agitated to dislodge the layer of cells covering the astroglial monolayer, predominantly yielding type-1 astrocytes with a flat morphology. The medium was replaced by fresh complete medium every 2–3 days.
BrdU labeling
To label proliferating progenitors in vivo, BrdU (Roche) was injected i.p. (single dose, 50 mg/kg) from day 2 after FiFx transection once each day for 5 consecutive days. To detect cell proliferation in vitro, BrdU was added into the medium at a concentration of 5 nM for 24 h.
Differentiation of NSCs
Neurospheres derived from the FiFx were digested into single-cell suspensions in Accutase and subsequently plated onto poly-
Immunofluorescence
Animals with left FiFx lesions were perfused with 0.9% (w/v) NaCl and 4% (w/v) paraformaldehyde in 0.1 M phosphate buffer (PB). Coronal sections through the hippocampus (−2.8 to −4.4 mm from the bregma) were prepared using a cryostat (Leica CM1900), blocked in 10% goat serum in PBST [0.01 M sodium phosphate buffer, pH 7.4, containing 0.05% (v/v) Tween 20] for 1 h at room temperature (RT). The cultured cells were fixed with 4% paraformaldehyde in 0.1 M PB for 1 h and blocked in 5% goat serum in PBST for 30 min at RT. The sections or cells were then incubated with rat or mouse monoclonal anti-BrdU (1:200); mouse anti-CD15 (1:100; Abcam); mouse or rat monoclonal antibodies directed against nestin, MAP2 (1:300), or 2′,3′-cyclic nucleotide 3′ phosphodiesterase (CNP, 1:500); chicken anti-BT3 (1:800); rabbit anti-glial fibrillary acidic protein (GFAP; 1:500); or guinea-pig anti-DCX (1:500) antibodies (Millipore) overnight at 4°C, followed by incubation with Alexa Fluor 568-conjugated goat anti-rat or chicken IgG (Molecular Probes), fluorescein isothiocyanate-conjugated goat anti-mouse or rabbit IgG (Chemicon), or 488-conjugated goat anti-guinea pig IgG (invitrogen). To detect BrdU, sections were pretreated in 2 N HCl at 37°C for 20 min and neutralized with sodium borate buffer (0.1 M, pH 8.5) at RT for 10 min before incubation with antibody. Immunofluorescence signals were visualized at excitation/emission wavelengths of 578/603 nm (Alexa Fluor 568) or 495/520 nm (fluorescein isothiocyanate), respectively, under a fluorescence microscope (Leica DMIRB).
Statistical analysis
The positive cells in consecutive coronal brain sections and in vitro random view fields were counted by single blind method. Statistical analysis was carried out using GraphPad software (GraphPad Prism v4.0; GraphPad Software). Comparison of differences between the experimental groups was performed using either 1-way analysis of variance test followed by Dunnet test or Student's t-test for unpaired values. P < 0.05 was considered significant.
Results
Nestin expression induced by FiFx transection
We previously reported that endogenous NSC proliferation takes place in hippocampus after surgical denervation [32]. We therefore investigated whether FiFx plus CB lesion promoted neuroregenerative processes in the region of the lesion itself. Hippocampus denervation surgery was performed by FiFx transection (Fig. 1A) with extensive damage of CB; Nissl staining confirmed that the left FiFx and CB remained disconnected at day 7 postsurgery (Fig. 1B). The sections through hippocampus posterior to the site of transection were examined for immunofluorescence with antibody against nestin, an intermediate filament protein predominantly expressed by NSCs. Denervation surgery by FiFx transection damages other structures above the FiFx, and in addition to the FiFx, we examined the CB and MBN. As shown in Fig. 1C and D, all 3 structures on the FiFx-transected side were enriched in nestin-positive cells, whereas such cells were almost completely absent from the contralateral (unoperated) site. Nestin expression suggests that NSC proliferation takes place in these brain regions following surgical lesion.

Transection of fimbria fornix (FiFx) and cingulate bundle (CB) induces local nestin expression.
NSC regeneration triggered by surgery
To determine whether the nestin-positive cells represent proliferating cells, BrdU was used to label dividing cells in vivo. Double staining for BrdU and nestin revealed that about 60% of nestin-positive cells in the CB, 70% in the FiFx, and 30% in the MBN were immunoreactive for anti-BrdU antibody (Fig. 2A, C, E), demonstrating that these cells were active for DNA synthesis. Quantification of the labeled cell numbers in consecutive sections revealed 583 ± 73.4 nestin+ BrdU+ cells in the CB, 910 ± 87.1 in the FiFx, and 245 ± 56.7 in the MBN per hemisphere (n = 5) (Fig. 2B, D, and F). Almost no double-positive cells were seen in the contralateral FiFx, CB, or MBN (Fig. 2A, C and E). Nestin+ BrdU+ cells, which were predominantly only found on the lesioned side of the brain, are therefore strong candidates for NSCs. The nestin− BrdU+ cells were about 197 ± 73.5 in CB, 93 ± 34.7 in FiFx, and 570 ± 93.8 in MBN (n = 5), which are presumed to be proliferating glial cells. The nestin+ BrdU− cells were about 367 ± 56.4 in CB, 395 ± 68.8 in FiFx, and 552 ± 77.2 in MBN (n = 5), which may represent resting neural progenitors that did not enter mitosis.

Proliferative neural stem cells (NSCs) in FiFx, CB, and MBN following FiFx plus CB transection. Following surgery, dividing cells were labeled by intraperitoneal administration of 5-bromo-2-deoxy-uridine (BrdU; single dose, 50 mg/kg) once a day for 5 consecutive days. Consecutive coronal sections were immunostained with anti-BrdU and anti-nestin antibodies at day 7 following surgery. BrdU-positive (red), nestin-positive (green), and double-positive cells (red nucleus with green body and processes) in CB
Immunostaining for GFAP, a marker of NSCs and of mature astrocytes [37,38], revealed that almost all nestin-positive cells were also immunoreactive for GFAP (Fig. 3A). Additionally, in the FiFx, the nestin-positive cells were predominantly CD15 positive, a known marker of NSCs (Fig. 3B). However, in other regions, the GFAP-positive cells were not nestin immunoreactive (Fig. 3C), and these are presumed to be mature astrocytes. Together these results indicate that extensive FiFx transection stimulates NSC proliferation in CB, FiFx, and MBN.

Glial fibrillary acidic protein (GFAP) expression is associated with nestin immunoreactivity in cholinergic and noncholinergic regions.
FiFx neuronal regeneration
FiFx is predominantly composed of glial cells and neuronal fibers and it is thought that FiFx itself contains few or no neuronal cell bodies. Because NSC proliferation was detected specifically in FiFx, CB, and MBN regions adjoining the surgical lesion, we examined whether newborn and mature neurons were generated. The sections at 7 or 35 days following FiFx transection were stained with antibodies directed against DCX and BT3. DCX, a microtubule-associated protein expressed almost exclusively in developing and migrating neurons, is increasingly employed as a marker of neurogenesis and/or immature neurons [39,40]. DCX-positive cells at a total number of 156 ± 33.6 (n = 5) per hemisphere were observed in the lesioned FiFx at day 7 after transection, whereas no DCX-positive cells were detected in the contralateral FiFx (Fig. 4A, B). The DCX-positive neurons per hemisphere in the CB and MSN of the lesion side numbered about 10 times that of the control (Supplementary Fig. S1; Supplementary Data are available online at
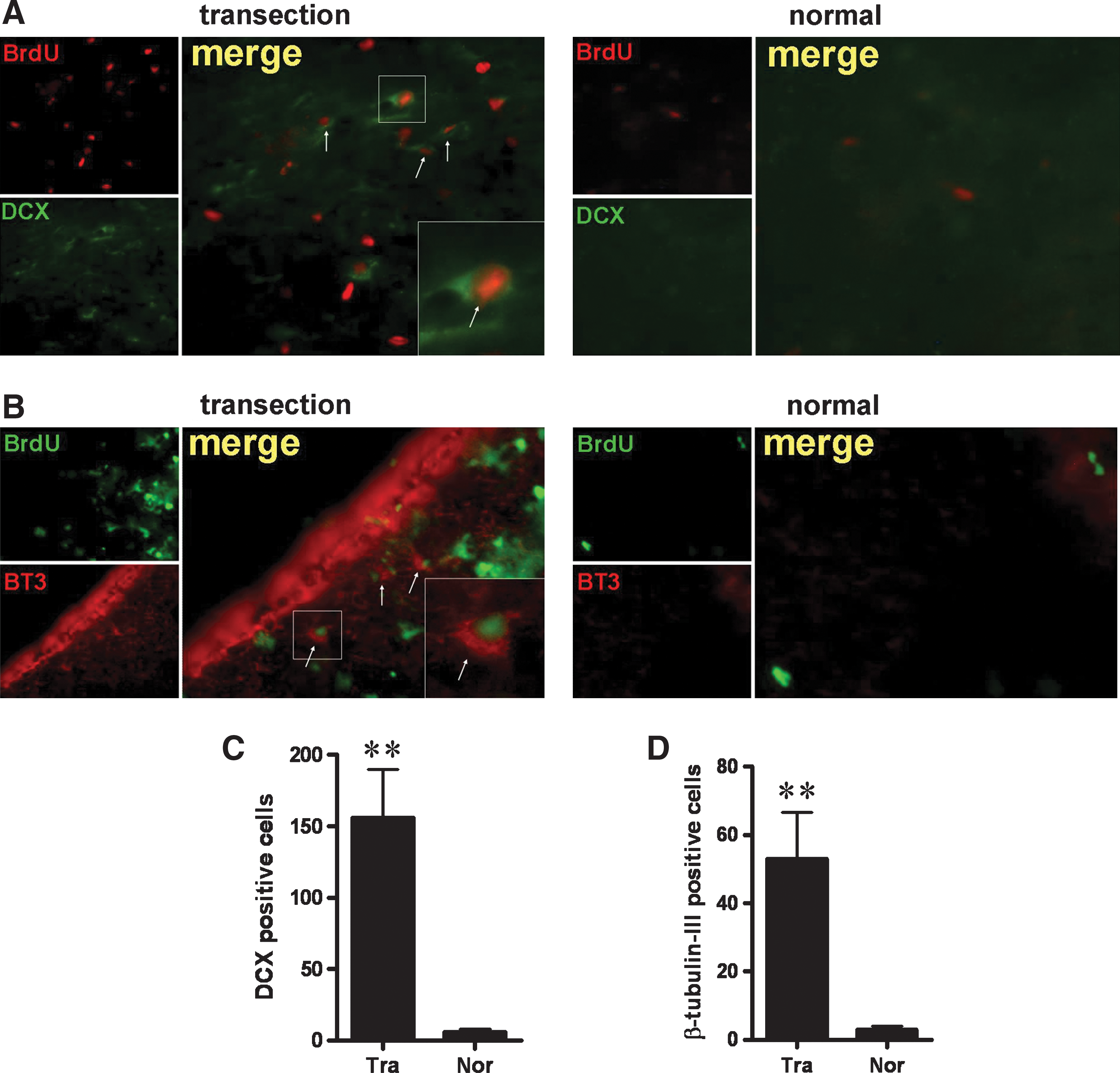
Neuronal development is stimulated in lesioned FiFx.
In vitro culture of FiFx NSCs
The above findings argue that surgical denervation induces NSC proliferation and differentiation in CB, FiFx, and MBN. We therefore used in vitro culture to confirm that surgical injury led to an increase in the number of proliferating NSCs. Brain tissues corresponding to the FiFx on both the lesioned and contralateral sides (blue areas in Fig. 5A) were carefully dissected under a microscope; care was taken to not include hippocampal tissue (Fig. 5B). These tissues were then cultured in vitro under conditions appropriate for NSC proliferation and differentiation (Materials and Methods section). No neurospheres were obtained from either the lesioned or contralateral FiFx tissues at day 3 following surgery. The in vitro culture of lesioned FiFx at day 7 following surgery gave rise to a significant number of neurospheres (Fig. 5C), whereas the culture of contralateral (normal) FiFx tissue failed to generate neurospheres. FiFx-derived neurospheres were slow growing, and culture for a period of ∼14 days was required to form neurospheres with 100–200 cells. The slow development of FiFx neurospheres contrasts with the relatively rapid in vitro development of neurospheres from either hippocampus or the SVZ [31,36].
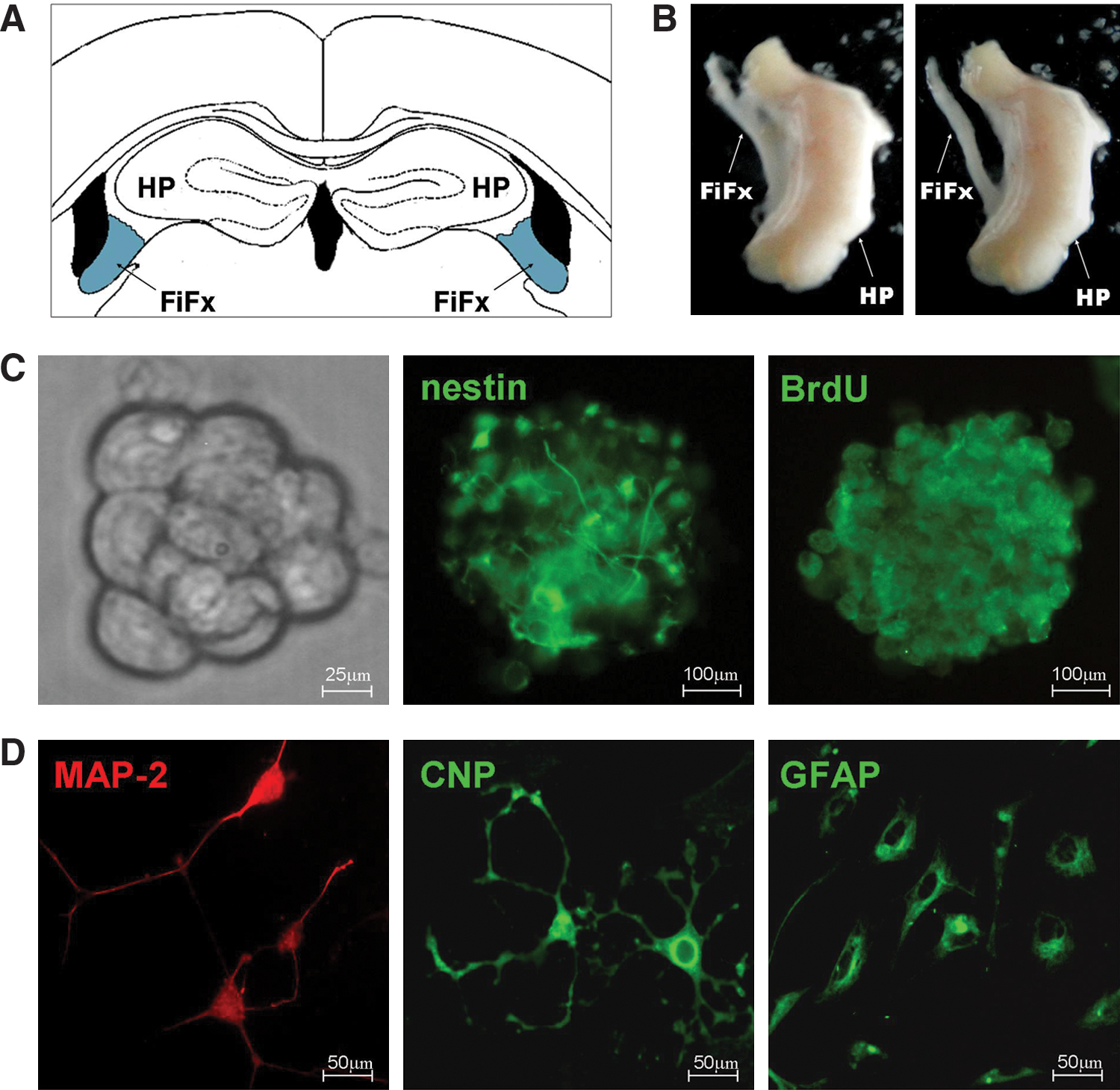
In vitro neurospheres from FiFx display characteristics of NSCs. The 7-day lesion and normal FiFx were dissociated and cultured in vitro. Neurospheres generated were evaluated for proliferation and differentiation.
To confirm that the cell aggregates developing in vitro were indeed neurospheres, we performed BrdU labeling and nestin staining. This revealed that the cell clusters were systematically BrdU and nestin positive (Fig. 5C). To assess the pluripotency of the presumptive neurospheres, the cell clusters were cultured in 1% FBS medium lacking epidermal growth factor and bFGF to promote differentiation. After 10 days in differentiation culture, the cells were analyzed for MAP2, GFAP, and CNP immunostaining. This revealed that a proportion of cells were MAP2 positive (and are therefore presumed to be neurons) and some were GFAP positive (astrocytes), whereas others were immunoreactive for CNP (oligodendrocytes) (Fig. 5D). Together these findings argue that the neurospheres derived from the lesioned FiFx tissue are generated from proliferative multipotent NSCs corresponding to the nestin+ BrdU+ cells seen in the sections of in vivo tissue from the lesioned FiFx (Fig. 3).
Glial cell proliferation in FiFx
Proliferation of glial cells including astrocytes typically takes place following brain damage and is thought to contribute to repair. To determine if FiFx plus CB lesion is accompanied by proliferation of glia, immunohistochemistry was performed on lesioned and contralateral brain using anti-GFAP antibody. On postlesion day 7, extensive GFAP immunoreactivity was detected in the lesioned FiFx, and the extent of staining was significantly greater than in the contralateral FiFx (Fig. 6A, B); the same increase in GFAP positivity was seen in CB (data not shown). To confirm that FiFx lesion promotes glial cell proliferation, FiFx tissues from the lesioned and contralateral sides of brain were cultured in vitro under conditions promoting astrocyte proliferation (Materials and Methods section). As shown in Fig. 6C and D, a significantly greater number of GFAP-positive astrocytes were generated in the primary cultures from the lesioned FiFx than from the contralateral unoperated FiFx. Together these results demonstrate that FiFx transection promotes local astrocyte proliferation.
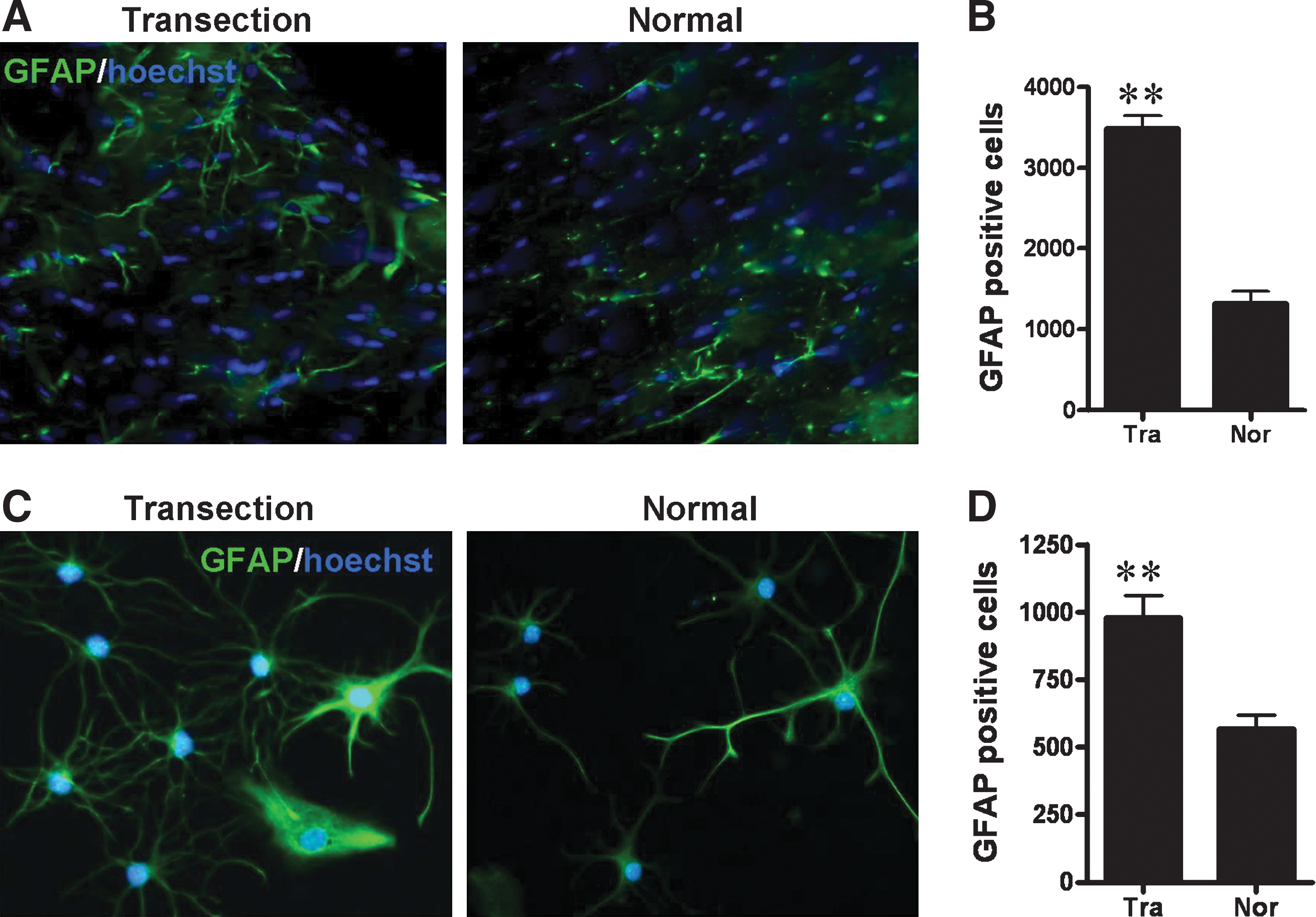
Astrocyte reactivity in FiFx.
Origin of FiFx neuroregeneration
Although a significant neuroregeneration in FiFx, CB, and MBN was detected in the lesion side, the sources initiating NSC regeneration in these areas remained unclear. To demonstrate whether NSCs in lesion FiFx were from the adjacent DG, GFP-subcloned lentivirus was used to label cells in DG, so as to track their migration. Seven days after FiFx plus CB transection, coronal sections through hippocampus were directly observed under fluorescence microscope. The results showed that the lentivirus-labeled cells in the lesion side migrated a longer distance in the subgranular cell layer than those in the normal side where migration was observed as well (Fig. 7A, B). However, few or no GFP-labeling cells were found in either lesion or normal FiFx at this time point (Fig. 7C) when significant NSC regeneration was observed, that is, these NSCs in FiFx stimulated by lesion did not originate from DG.
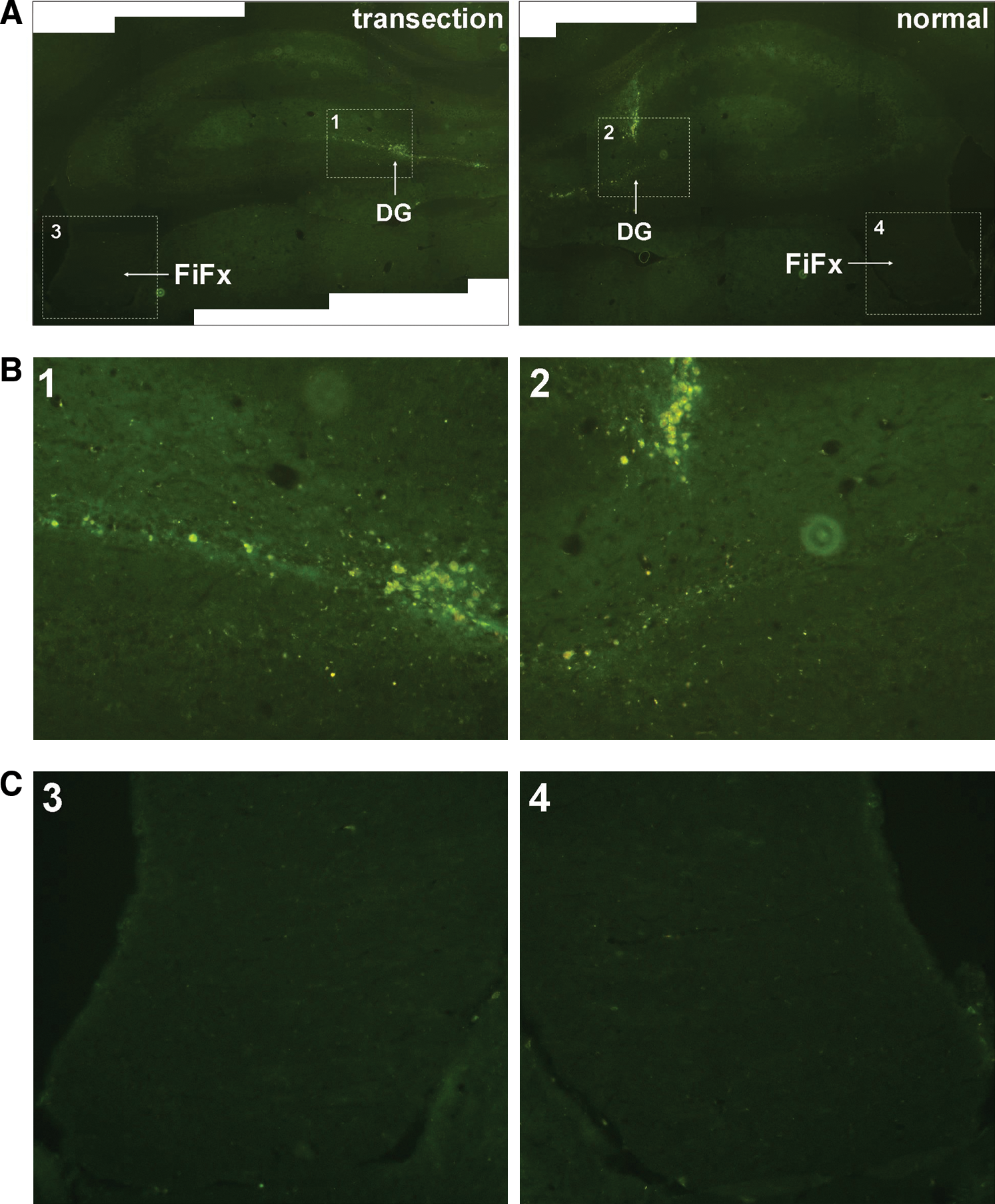
In vivo tracking of dentate gyrus (DG) cells transfected with GFP lentivirus. Lentivirus subcloned with GFP-coding sequence is injected into DG at 3 days before unilateral FiFx transection. Seven days after transection, GFP-labeled cells are detected under fluorescence microscope.
Discussion
Endogenous NSCs within the adult mammalian brain are known to undergo enhanced proliferation and differentiation in response to neurodegeneration and brain injury including aspiration or transection lesions, inflammatory or chemical demyelination, and percussion trauma [10,41 –43]. The present and previous reports have demonstrated that transection of cholinergic projections stimulates neuroregeneration in forebrain cholinergic regions. The basal forebrain cholinergic system is of major importance to human brain function [44 –46]. The cholinergic basal forebrain comprises magnocellular hyperchromic neurons within the septal/diagonal band complex and the NBM; these provide the predominant cholinergic innervation to the hippocampus, amygdala, and neocortex in the mammalian brain.
Cholinergic input plays an important role in cognition and attentional behaviors, and cholinergic dysfunction is a prominent feature of dementias including AD. Hippocampal afferents from the septal area, the locus coeruleus, and the raphe nuclei are routed via 3 anatomically distinct pathways, namely the CB, the FiFx, and a ventral pathway whose exact anatomical location is not well defined but is thought to reach the hippocampus after passing in the vicinity of the amygdalar complex. Afferents via the FiFx and CB provide the hippocampus with a large proportion of its cholinergic, extrinsic GABAergic, noradrenergic, and serotonergic inputs, whereas efferents carrying fibers from hippocampal pyramidal CA2–CA4 cells project to the anterior thalamic nucleus, medial mamillary nucleus, cingular gyrus, and the NBM [25]. Cholinergic projections comprise a complex neural network that supports higher brain functions, and the FiFx and CB are the principal cholinergic pathways that communicate between the basal forebrain and hippocampus and cortex. Lesions of the fornix plus cingulum lead to substantially reduced cholinergic innervation [29] and produce lasting impairments of spatial learning and memory [30], all of which are among the earliest events in the pathogenesis of AD [47 –49]. Lesions of the fornix plus cingulum have therefore been widely used in experimental animal models of dementia [30,50,51].
Our previous studies have focused on processes underlying hippocampal regeneration in such animal models. Transection of the FiFx and overlying structures interrupts not only FiFx projections but also the CB commissural fibers, resulting in elimination of both afferent and efferent fibers, but predominantly affects cholinergic innervation of hippocampus and cortex [31]. We previously reported that transection of FiFx plus CB was followed by a period of local NSC proliferation, migration, and neuronal differentiation in the DG of the hippocampus [32]. The denervated hippocampus provides a rich environment for neuronal differentiation of either endogenous or exogenous NSCs [31,32], and the changes elicited by FiFx plus CB transection are likely to underlie hippocampal repair and regeneration following injury [36].
In addition to inducing changes in the local microenvironment of innervation-deprived target regions, damage to projecting fibers such as the FiFx induces physiological changes in the anatomical pathways involved, such as loss of mitochondrial membrane potential [52,53], calcium influx, and calpain activation [54,55]. In the present study, we report another change, neuroregenerative processes, taking place in the FiFx and adjoining tissue within 1.5 mm of the transection site. Following FiFx transection, large numbers of BrdU+ or nestin+ cells were detected; the majority were BrdU+/nestin+ and are therefore strong candidates for proliferative NSCs. In vitro, we successfully cultivated neurospheres from lesioned FiFx, but not from contralateral FiFx, and these most likely arise from BrdU+/nestin+ NSCs. Further, FiFx-derived neurospheres demonstrated pluripotency in that they generated cells positive for markers of neurons, astrocytes, and oligodendrocytes, supporting the contention that BrdU+/nestin+ cells in lesioned FiFx are NSCs. The fact that almost no BrdU+/nestin+ cells were observed in normal FiFx argues that NSC proliferation is induced by FiFx lesion, and this could argue that NSC proliferation is involved in repair and regeneration processes. Despite this, a small number of BrdU+/nestin+ cells were found in the contralateral FiFx, which may result from loss of nervous fiber connection between bilateral hippocampus through FiFx.
It is generally thought that the FiFx only contains projecting fibers and supporting astroglial cells and that neuronal cell bodies are absent. However, immunohistochemistry at day 7 following lesion revealed the presence of cells positive for DCX, a microtubule-associated protein expressed almost exclusively by developing neurons [39,40]. Five weeks after transection, we detected cells expressing BT3, a specific marker of newborn and mature neurons, in the lesioned FiFx. The sequential appearance of immature and mature neurons suggests that neurogenesis and maturation takes place in the lesioned FiFx and that these neuronal cells arise from induced proliferation of NSCs. Parallel results were obtained in CB and MBN, arguing that similar processes also take place in response to injury in these regions.
Neural regeneration has been reported in several brain regions following injury or disease [9,56]. It is generally assumed that NSC proliferation and differentiation are intended to restore lost neurons and/or compensate for decreased transmitter levels. However, the possibility that such regenerative processes might extend to cholinergic projections has not been previously addressed. The present study, in conjunction with our previously reports, argues that NSC proliferation and differentiation take place not only in hippocampus and MBN but also in projecting regions including the FiFx and CB.
The best studied and validated reservoirs for neural stem and progenitor cells in the adult mammalian brain are the SVZ of the lateral ventricles and the SGZ of the DG [57 –60]. Under normal conditions, the postnatal SVZ contributes progenitors to the rostral migratory stream to support ongoing olfactory neurogenesis, whereas the SGZ of the DG provides new granule neurons throughout life [58,61]. However, neurogenesis in these 2 principal areas is known to be stimulated by brain insults such as stroke or trauma. CNS injury in adult rodents can stimulate the proliferation of neuronal stem/progenitor cells located in the SVZ and the SGZ, and the resulting newborn cells have been reported to migrate into damaged brain regions where they express mature neuronal markers. This lends weight to the contention that damaged cells can be replaced from endogenous NSC pools.
In the present study, it is therefore possible that FiFx injury promoted the local proliferation and differentiation of neural precursors migrating from the adjacent SGZ pools into the FiFx. The tracking of SGZ cells using lentivirus argues that NSC regeneration in the injured FiFx was not the result of migration from SGZ pools. Radial glial cells expressing vimentin, GFAP, and nestin provide a second potential source for proliferative NSCs in the FiFx. During development, radial glial cells contribute to neurogenesis and neuronal migration and differentiate into astrocytes at the end of the developmental period [62]. It was recently demonstrated that radial glial cells remaining in the adult brain can function as primary progenitors or NSCs and can generate new neurons throughout adulthood [63,64]. Injury could therefore induce the proliferation and differentiation of these glial cells in the adult. The emerging evidences demonstrated that the astroglial cells can generate neurons not only during development but also throughout adult life and potentially even after brain lesion [37,65,66]. Imura et al. [38] have reported that there are 2 types of astrocytes in the adult brain—neurogenic GFAP-expressing astrocytes that are restricted to the forebrain, and nonneurogenic GFAP-expressing astrocytes that are widely distributed throughout the brain. We surmise that the cholinergic system could contain neurogenic astrocytes and that injury stimulates the proliferation and differentiation of these cells. Given current interest in cell-based therapies for the treatment of degenerative conditions affecting the cholinergic system, such as AD, it will be crucially important to determine the respective contributions of NSCs, neurogenic astrocytes, and other potentially neurogenic cell types in repair and regeneration processes taking place in response to brain injury.
Footnotes
Acknowledgments
This study was supported by grants from the Natural Science Foundation of China (No. 30670648), Nature Foundation of Jiangsu (No. BK2006057), College Nature Foundation of Jiangsu (No. 04KJB180111), and Undergraduate Innovative Research Foundation of Jiangsu.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.