
Abstract
Mesenchymal stem cells (MSCs) have emerged as important tools for cell therapy; therefore, identification of factors capable of governing their ex vivo expansion become essential. In this study we demonstrate that human adipose-derived stem cells (ASCs) express all components of the bone morphogenetic protein (BMP)/BMP receptor signaling pathway and respond to BMP4 inducing upregulated expression of its specific target genes Id1–Id4. Moreover, ASCs grown in a medium reduced in serum produce endogenous BMP4 that could affect autocrinely ASC growth. On the contrary, dorsomorphin, an inhibitor of BMP signaling pathway, decreases cell numbers yielded from ASC cultures in correlation with increased apoptosis and decreased cycling cells. Therefore, BMP4 emerges as a possible factor for ex vivo expanding human ASCs. Our results demonstrate that, as other morphogens, BMP4 effects on human MSCs are dose dependent. High doses significantly increased apoptosis and drastically reduced cell proliferation, whereas low doses of BMP4 (0.01–0.1 ng/mL) significantly increase culture cell content, reduce the number of apoptotic cells, and increase that of cycling cells. Further, treatment of human ASCs with low doses of BMP4 does not modify expression of Nanog and Oct4, two transcription factors involved in self-renewal and pluripotency of stem cells or avoid their osteogenic or osteoblastic differentiation capacities when cultured in adequate inducing media, as shown by the induction of specific gene expression (CEBP, PPARγ, and RUNX2). Our results therefore support BMP4 as a promising factor for expanding human adipose tissue-derived MSCs maintaining their properties of stemness and multipotency.
Introduction
M
However, a critical problem of MSCs is their limited ex vivo growth rate that makes mandatory their expansion in vitro to obtain a sufficient number of cells for cell therapy. Currently, MSCs destined for clinical therapy are expanded before transplantation in the culture medium supplemented with 10%–20% fetal bovine serum (FBS), increasing the potential risk of transferring animal proteins to human cells. To drastically reduce the animal-derived serum concentration or to use reagents of human origin become, therefore, critical issues for improving the therapeutical applications of MSCs. It is important to identify growth factors capable of inducing MSC expansion maintaining their properties of survival and multilineage differentiation capacities to an extensive therapeutic application of MSCs. In the last years, several factors, especially different members of transforming growth factor β superfamily, have been used with the proposal of increase in vitro the number of MSCs. Remarkably, MSCs survive at low density in the absence of serum [6], a phenomenon that has been recently associated with bone morphogenetic protein (BMP) signaling [7].
BMPs, members of transforming growth factor β superfamily, were initially recognized as bone inductive proteins but later demonstrated to be involved in the regulation of cell proliferation, differentiation, and survival/death during embryonic development, but also in adult tissue remodeling and regeneration [8,9]. BMPs interact with 2 types of receptors: type I (Alk2, Alk3–BMPRIA−, and Alk6–BMPRIB−) and type II (BMPRII). Different combinations of type II receptor with any one of type I receptors may determine specificity and result in different effects [9]. Although other signaling pathways in BMP signal transduction are known, canonical BMP pathway is mediated by phosphorylation of R-Smad (Smad 1, Smad 5, and Smad 8). Two phosphorylated R-Smads form a heterotrimer with the common Smad 4 (co-Smad) that translocate into the nucleus and, together with other transcription factors, trigger target gene expression [10].
Effects of BMPs on MSCs are, however, controversial. There are more than 20 BMPs; some exhibit a distinct function on MSCs, while others have overlapping effects depending on the specificity of their interactions with distinct types of receptors and tissues in which they are differentially expressed [11]. In the present study, we demonstrate expression of different components of the BMP2/4 signaling pathway and show that this canonical pathway is active in ASCs. Further, our results demonstrate that effects of BMP4 on proliferation and survival of ASCs are dose dependent and that low doses of exogenous BMP4 maintain expression of transcription factors (Nanog and Oct 4) key to ensure the stemness and self-renewal properties of undifferentiated ASCs without affecting their pluripotency. BMP4 may be therefore a good candidate for using in ex vivo extensive production of human MSCs for therapeutic application.
Material and Methods
Cell culture
ASCs were isolated as described before [12] from human adipose tissue from 6 healthy women (aged 30–35 years) with a body mass index considered as normal according to parameters of World Health Organization (18.5–25 kg/m2), after informed consent according to the guidelines of the Ethics Committee of the Hospital La Paz (Madrid, Spain). Briefly, the cells were obtained by elective liposuction procedures (4 thigh, 1 hip, and 1 abdomen) and isolated in Medio Dulbbecco's modified Eagle's medium (DMEM) high glucose (Gibco, Invitrogen) with 100 U/mL penicillin/100 μg/mL streptomycin and 10% FBS. The cells were cryopreserved in passage 2 using DMEM, with 100 U/mL penicillin/100μg/mL streptomycin, 10% FBS, and 10% DMSO freezing overnight in isopropanol container at −80°C and after conserved in liquid nitrogen. For in vitro experiments frozen aliquots of ASCs were thawed and cultured in complete medium containing DMEM, 10% FBS, and antibiotics.
After initial passages, the cells were maintained and expanded in a reduced serum (2%) medium, MesenPRO-RS™ (Gibco-Invitrogen), containing 100 U/mL penicillin/100 μg/mL streptomycin and 1 mM L-glutamine in a 5% CO2-in-air incubator at 37°C. Once the cultures reached 80%–90% confluence, the cells were recovered by addition of 0.25% trypsin solution (Gibco-Invitrogen) and counted in a hemocytometer. Cell viability was assessed by trypan blue staining, and, then, the cells were replated at a density of 5 × 103 cells/cm2. The medium was replaced each 4–5 days. All experiments were performed with cells harvested between the 5th and 10th passage.
Differentiation assays
The procedures were adapted from the protocols previously established [13]. ASCs were plated at a cell density of 5,000 cells/cm2 in 6-well tissue culture plates and grown for 6 days in MesenPRO-RS as control or MesenPRO-RS supplied with 0.01 ng/mL BMP4. Then, they were moved to the conditionating medium consisted of either minimum essential medium alpha (Gibco-Invitrogen) for osteogenic differentiation or DMEM (Gibco-Invitrogen) for adipogenic differentiation, supplemented with 5% FBS and 100 U/mL penicillin/100ug/mL streptomycin, by 1 day, and then the cells were replaced with the differentiation medium.
The adipogenic medium contained 1:1 DMEM, F-12 Nutrient mixture (Ham) (Gibco-Invitrogen), and 3% FBS, supplemented with 1 μM dexametasone, 60 μM Indometacine, 0.5 mM 3-isobutyl-1-methylxantine (IBMX), and 100 U/mL penicillin/100 μg/mL streptomycin.
The osteogenic medium included minimum essential medium alpha supplemented with 10% FBS, 10 nM dexametasone, 10 mM β-glycerophosphate, and 60 μM ascorbic acid with 100 U/mL penicillin/100 μg/mL streptomycin. All reagents used in the differentiation media were purchased from Sigma.
After 5 days of differentiation the cells were collected to extract RNA and to perform quantitative polymerase chain reactions (PCRs).
BMP4 and dorsomorphin treatment
ASCs were detached, counted, and cultured during 6 days in MesenPRO-RS™ medium supplemented with different concentrations of human recombinant BMP4 (ranging 0.01–100 ng/mL; R&D Systems). For treatment with dorsomorphin, an inhibitor of the canonical BMP signaling pathway (Calbiochem), cells were cultured in MesenPRO-RS™ medium for 24 h and then the medium was supplemented with the inhibitor (5–10 μM) for 5 days more. After treatment, human adipose tissue-derived MSCs were harvested and used for differentiation assays or processed for proliferation assays, viability assays, and PCR analysis.
PCR analysis
RNA was isolated using Absolutely RNA Microprep kit (Stratagene Cloning Systems), including a DNase I digestion step, as recommended by the supplier, to avoid genomic DNA contamination. Total cDNA was synthesized by High Capacity cDNA Reverse Transcription Kit (Applied Biosystems), according to the supplier's instructions, and then used as a target in the PCR amplifications. Standard PCR was carried out as previously described [14]. Real-time PCR was performed with the following TaqMan assays: BMPRIA (Hs01034909_g1), BMPRIB (Hs00176144_m1), BAMBI (Hs00180818_m1), Id1 (Hs00357821_g1), Id2 (Hs00747379_m1), Id3 (Hs00171409_m1), Id4 (Hs00155465_m1), NANOG (Hs2387400_g1), OCT4 (Hs3005111_g1), CEBP (Hs269972_g1), PPARγ (Hs1115513_m1), and Runx2 (Hs01047976_m1); all of these were obtained from Applied Biosystems. All PCR reactions were set in duplicates using the TaqMan Gene Expression Master Mix (Applied Biosystems) according to the manufacturer's instructions. Amplifications, detections, and analyses were performed in a 7.900HT Fast Real-time PCR System (Centro de Genómica, Complutense University). Gene expression was normalized with respect to relative expression of GAPDH (Hs002550_m1) and GNB2L1 (Hs 00272002_m1) housekeeping genes and was calculated based on the CT values, using the ΔΔCt methods since efficiency of all assays used have 100% PCR efficiency.
Flow cytometry
The following mAbs conjugated with FITC, PE, PE-Cy5, or APC were used for flow cytometric analysis: CD44, CD106, CD29, CD45, CD34, and CD14 from BD Biosciences and Immunostep (Spain). Two- and three-color immunofluorescence stainings were performed by incubating the cells in PBS containing 1% FCS and 0.1% NaN3 in the presence of saturating amounts of fluorochrome-conjugated antibodies for 30 min at 4°C. Analyses were conducted in a FACSCalibur flow cytometer (BD Biosciences) from the Centro de Microscopía y Citometría, Complutense University of Madrid.
Apoptosis assays
For apoptosis assays, cells were stained with Annexin-V-FITC (Boehringer Mannheim) according to the supplier's instructions. Cells were analyzed on a FACScan (Centro de Microscopía y Citometría Complutense University) and gated according to forward scatter, side scatter, and their ability to exclude propidium iodide. Annexin V-positive and propidium iodide-negative cells were considered apoptotic cells.
Proliferation assays
Cultures were pulsed for 12 h with 10 μM BrdU. A specific kit from Boehringer Mannheim (BrdU Labeling and Detection kit III) was used to measure BrdU incorporation into newly synthesized DNA. Briefly, the labeling medium was removed, and cells were dried (2 h at 60°C), fixed in ethanol in HCl (0.5 M) for 30 min at −20°C, treated with nucleases (30 min at 37°C), and then incubated with peroxidase-conjugated Fab of mouse anti-BrdU Abs (30 min at 37°C). The peroxidase reaction was developed with ABTS substrate, and the sample absorbance was measured using an ELISA reader at 405 nm with a reference wavelength at 492 nm.
BMP4 measurements
The culture supernatants were carefully collected and the concentrations of BMP4 were determined by using a specific ELISA kit (R&D Systems) according to the manufacturer's instructions. Absorbance (optical density) was measured at 450 nm. Concentrations were obtained by interpolation from standard curves.
Statistical analysis
The Student t test was used for statistical analysis. Values of P ≤ 0.05 (*), P ≤ 0.01 (**), and P ≤ 0.001 (***) were considered to be statistically significant.
Results
Preliminary characterization of the ASC was performed after 6 days of culture in a medium with reduced serum (see Materials and Methods). For this, we carried out a flow cytometry study for testing expression of CD44, CD106, CD29, CD45, CD34, and CD14 cell markers on cells ranging from passage 5 to 10. The cells maintained the typical MSC phenotype [15] expressing CD44, CD106, and CD29 markers but not CD45, CD34, or CD14 (Fig. 1A), as recently reported in bone marrow MSC cultures [7]. We also evaluated the growth kinetics of ASCs along 10 days of culture, confirming a gradual growth of ASCs after the stabilization of cultures (days 0–4) (Fig. 1B).
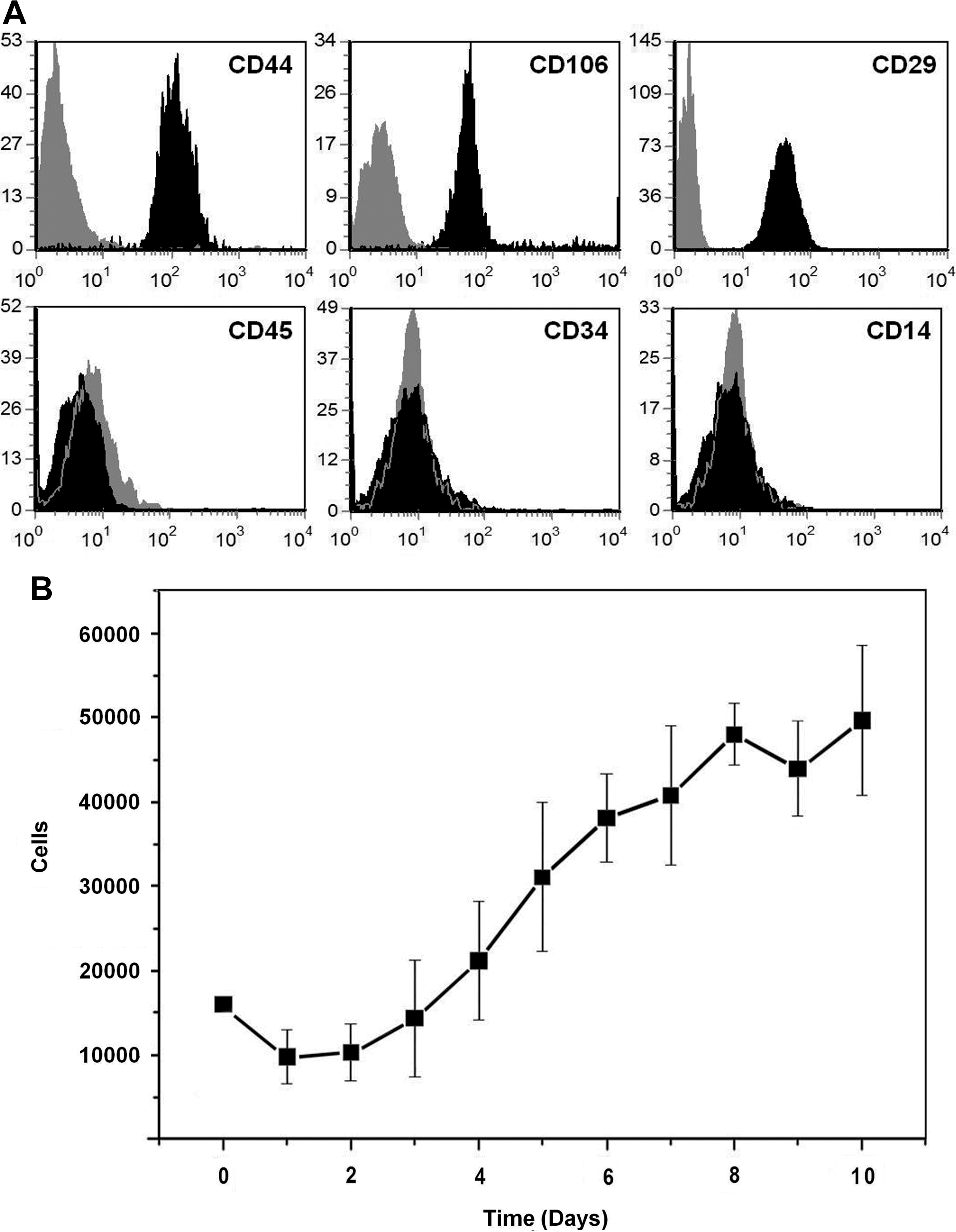
Adipose-derived stem cells cultured in the reduced serum medium maintain their typical phenotypical features and growth characteristics.
Because several authors have reported expression of BMP ligands [7,16 –20], receptors [7,19,21 –23], and some transducers (Smad 1) of the BMP signaling pathway [7] in MSCs derived from different sources, we analyzed by PCR the expression level of several BMP receptors and R- (Smad 1 and Smad 5) and co-Smads (Smad 4) involved in the canonical BMP signaling pathway [10] in human ASCs (hASCs) at passages 5–10. We found by quantitative PCR significantly higher expression of the type I receptor BMPR1A than BMPR1B or BAMBI, an inhibitory receptor of the BMPR type I [24] (Fig. 2A). Further, PCR analysis demonstrated expression of BMP transducers Smad 1, Smad 5, and Smad 4, indicating that hASCs contain the molecular machinery for transducing BMP-mediated signals (Fig. 2B). To confirm the suggested capacity of hASCs to transmit signals through BMP receptors, we analyzed the variations in the expression levels of inhibitors of differentiation (Id), transcription factors that are one of the main targets of the BMP signaling pathway [25], in hASCs after BMP4 supply (Fig. 2C). Treated cultures showed significant upregulated expression of Id1, Id2, Id3, and Id4 with respect to untreated, control ones as measured by quantitative PCR (Fig. 2C).
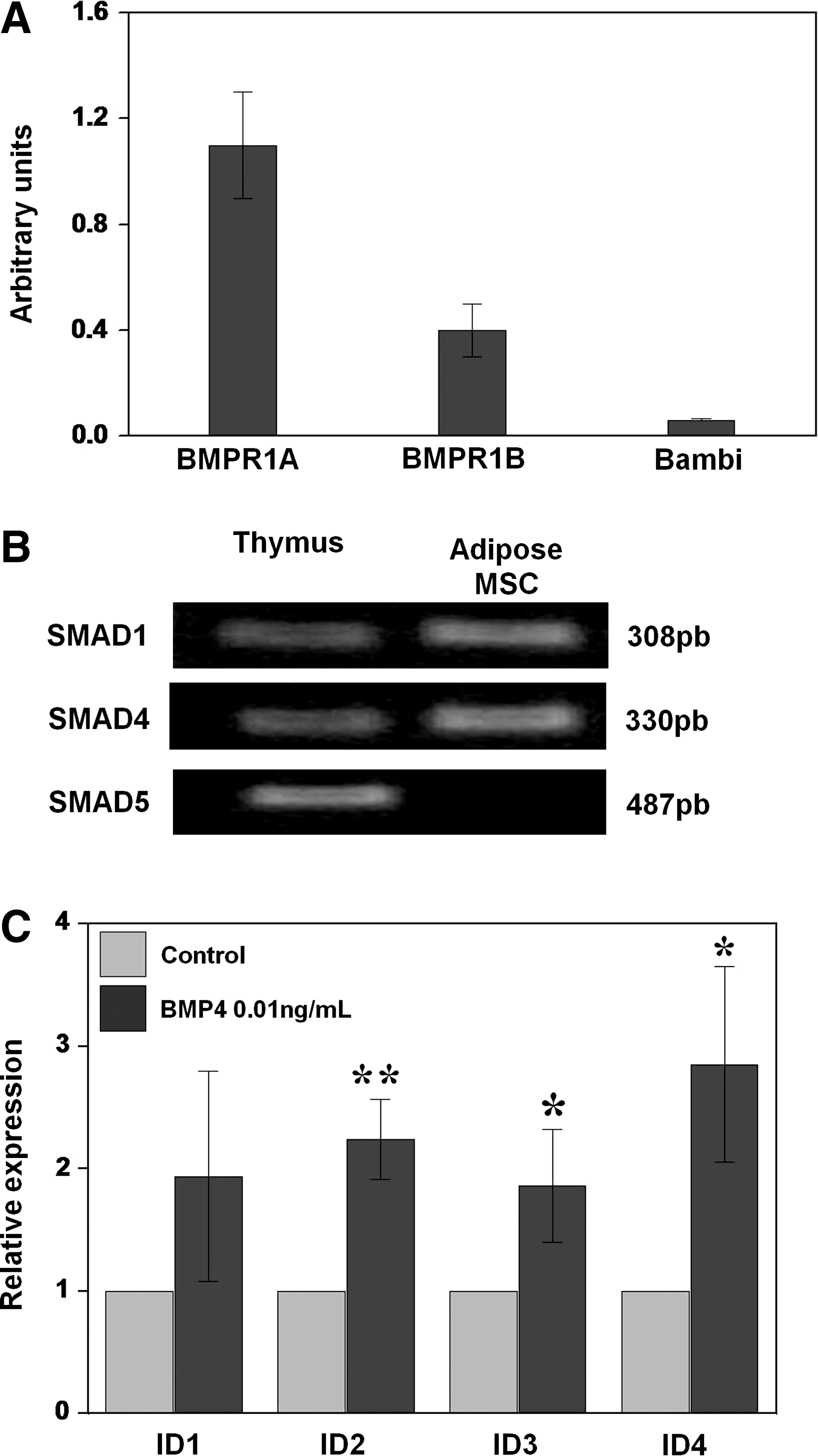
Functional expression of the BMP2/4 signaling pathway.
Because hESCs have a high levels of BMP activity in unconditioned culture medium [9], and BMP4 is expressed in several cell types closely related to MSCs (e.g., bone marrow stromal cells and perivascular cells) [26] and, moreover, Solmesky and colleagues [7] have pointed out recently that capacity of human bone marrow MSCs to grow in a serum-free medium is dependent on the endogenous production of BMPs, we measured by ELISA at different times of cultures over a period of 11 days the production of BMP4 in ASC culture supernatants. As shown in Fig. 3, hASCs, but not fibroblasts, grown in the medium with reduced serum produced BMP4, which reached highest values at day 3 of culture and later significantly decreased (days 6 and 11).
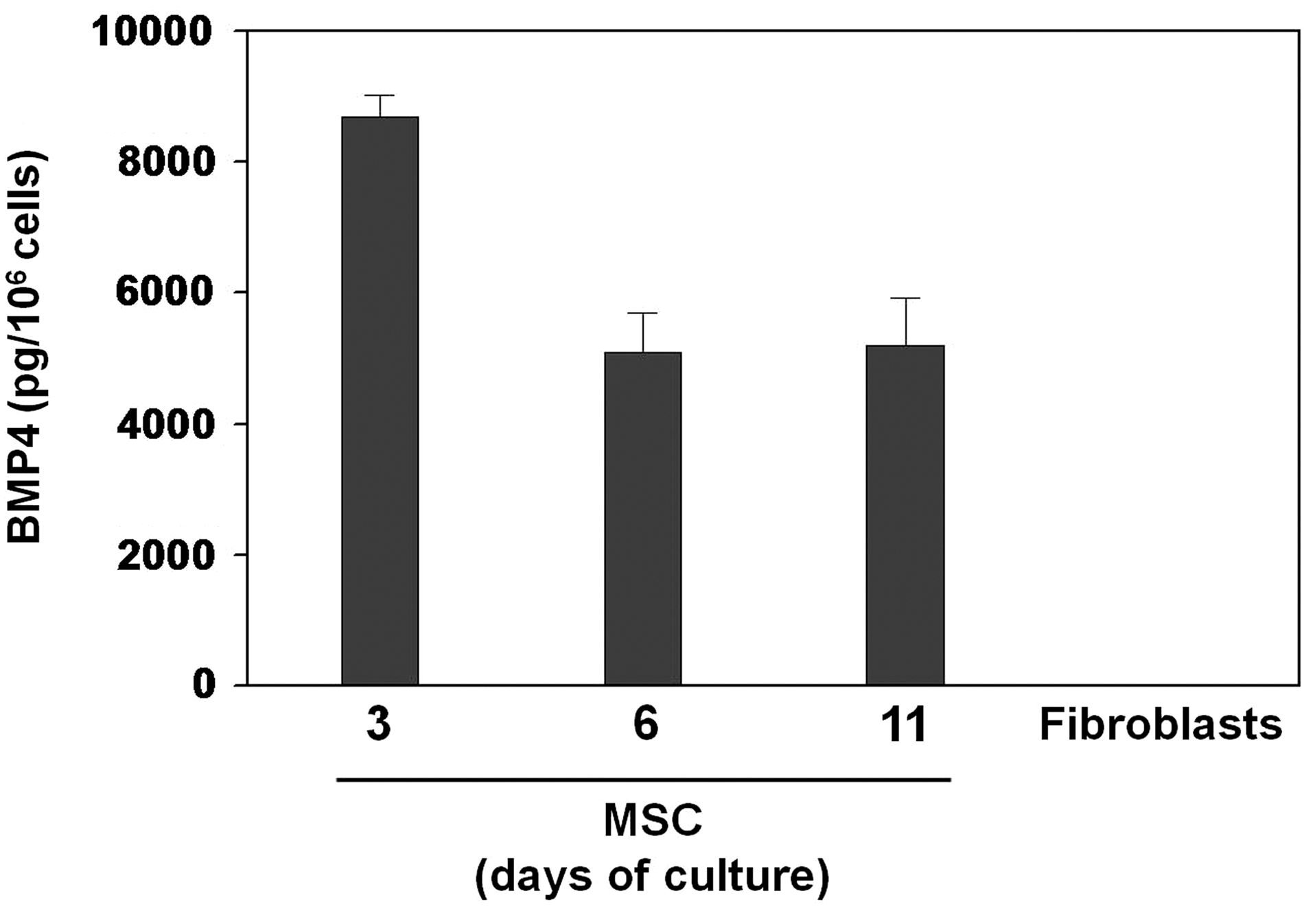
BMP4 production by adipose tissue-derived MSCs. Adipose-derived stem cells were cultured in reduced serum medium and the amount of BMP4 was assessed by ELISA at different culture time points in culture supernatants. No BMP4 was detected in supernatants from skin fibroblasts cultured in the same medium. Data represent the mean (±SD) of 3–4 independent experiments from 4 different ASC donors.
These results suggest that hASCs grow normally in cultures importantly reduced of serum and produce endogenous BMP4 that could autocrinely stimulate their growth, since hASCs express all components of BMP-dependent signaling pathway and respond to the morphogen upregulating expression of Id transcription factors.
We then tested the effects of BMP4 on the biology of hASCs. Dorsomorphin, a drug that inhibits the phosphorylation of R-Smads, induced through BMP the type I receptors [27]; the cell numbers yielded by ASC cultures decreased in a dose-dependent manner after 6 days of treatment. Although the drug was effective at the 2 doses tested, the highest effects were observed with 10 μM dorsomorphin (Fig. 4A). To understand the origin of this decreased growth of ASCs when BMP signaling was impeded, we evaluated rates of cell survival/death and proliferation in hASC cultures supplied with distinct doses of dorsomorphin (Fig. 4B, C). hASC viability was assessed after 6 days of culture by annexin V and propidium iodide staining. A significant increase of the proportion of annexin V-positive cells within the propidium iodine-negative cell population (apoptotic cells) occurred in the dorsomorphin-treated cultures (Fig. 4C). On the other hand, the percentage of cycling hASCs (BrdU incorporating cells) significantly decreased after the treatment with dorsomorphin (Fig. 4B). These effects were recovered when cultures treated with dorsomorphin received 0.01 ng/mL BMP4 (Fig. 4B).
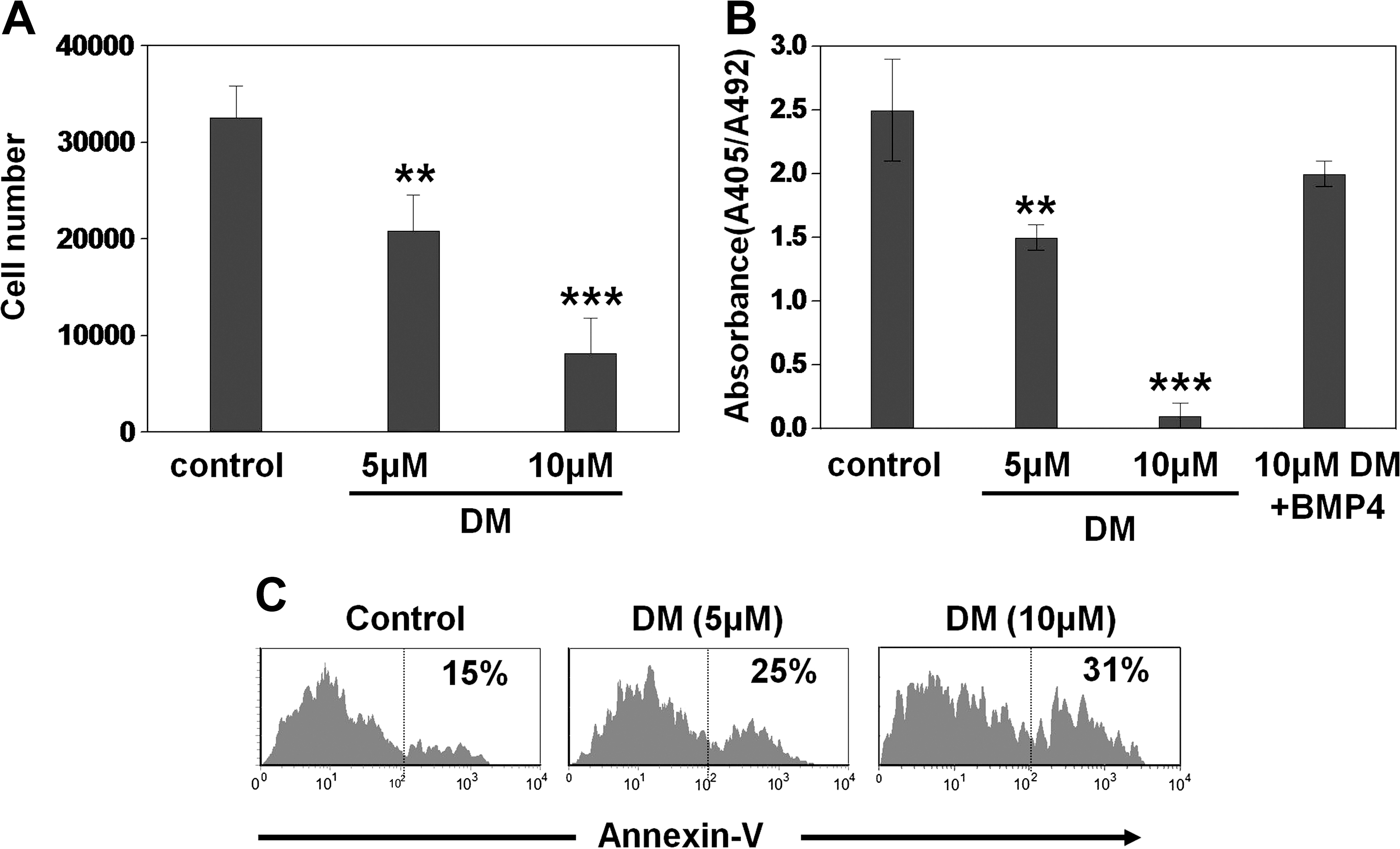
Canonical BMP signaling pathway regulates the proliferation and viability of adipose-derived stem cells. Cells were seeded and cultured for 5 days in MesenPRO-RS™ medium alone (control) or containing different doses of dorsomorphin.
Accordingly, these results demonstrate that canonical BMP signaling pathway regulates the proliferation and viability of hASCs governing therefore the numbers of cells yielded from cultures. In this respect, we expected that BMP4 treatment could improve the cell production of hASCs. High doses of BMP4 significantly increased the cell death (Fig. 5C) and drastically reduced the proportions of cycling cells (Fig. 5B), causing together a significant decrease in the cell numbers (Fig. 5A). On the contrary, low doses of BMP4 (range 0.01–0.1 ng/mL) significantly increased the proportions of cycling hASCs and decreased those of apoptotic cells (range 0.01–10 ng/mL) (Fig. 5B, C), improving the cell production of culture from day 6 onward (Fig. 5A).

Low-doses of rhBMP4 enhance the proliferation and survival of adipose-derived stem cells. hASCs were cultured in triplicate for several days in reduced serum medium alone (MesenPRO-RS™) (control) or supplemented with different doses of rhBMP4.
Several reports in distinct experimental models support a role for BMP not only in the survival of MSCs but also in their lineage commitment, although the results are controversial [28,29]. On this respect, it was important to clarify whether low doses of BMP4, which we have previously shown to induce increased cell proliferation and survival of hASCs, were able to alter their lineage commitment or their pluripotent properties, as inhibition of stem cell differentiation can lead to accumulation of stem cells [30 –32]. We first examined the effects of low doses of BMP4 (0.01 ng/mL) on expression of Nanog and Oct 4 transcription factors, known to be involved in the pluripotency and self-renewal of undifferentiated stem cells [33]. As shown in Fig. 6, the analysis by quantitative PCR of hASC cultures maintained for 6 days in the presence or not of BMP4 did not show significant differences in expression of the 2 studied genes. We further analyzed whether the pretreatment with low doses of BMP4 modified the capacity of hASCs to differentiate into the adipose or osteogenic lineage. To this purpose, hASCs were cultured for 6 days in the presence of 0.01 ng/mL BMP4, and transferred to media specifically inducing either adipose or osteoblast differentiation, and expression of genes specific of the adipose (CEBP and PPARγ) or osteogenic lineage (RUNX2) was analyzed (Fig. 7A). In these experimental conditions, our quantitative PCR analysis demonstrated increased expression of the specific genes studied in those cultures provided with the respective inducing medium (Fig. 7A). Further, MSCs that were cultured in the presence of low doses of BMP4 for 6 days and then transferred to either an osteoblast- or an adipose-inducing medium for 6 days more maintained their capacity to induce expression of specific genes involved in adipogenesis (CEBP and PPARγ) or osteogenesis (RUNX2) as control hASCs, which had not been pretreated with BMP4 (Fig. 7B). These data support that BMP4 pretreatment does not affect hASC properties to undergo specific differentiation.
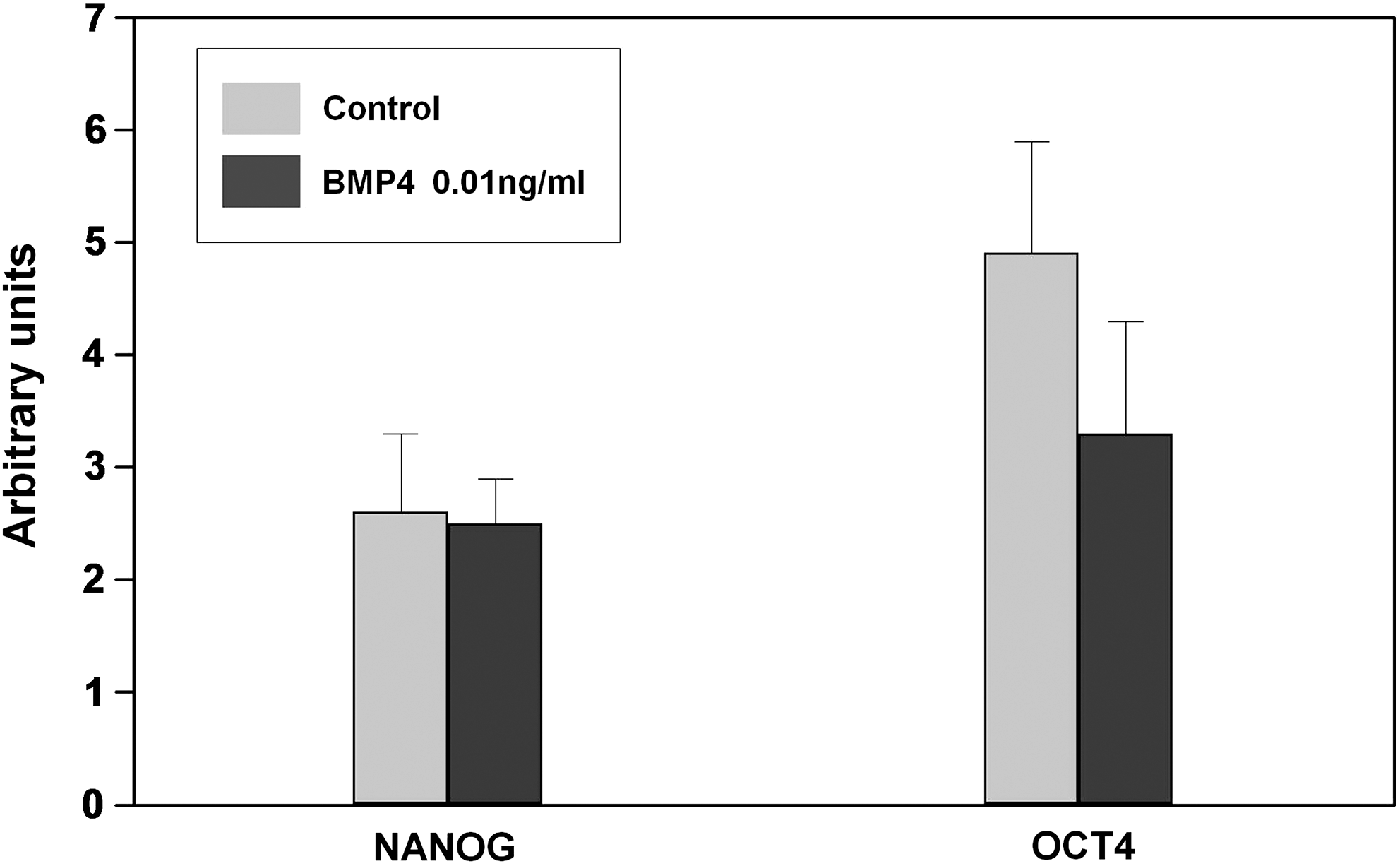
Adipose-derived stem cells treated with low doses of BMP4 maintain expression of transcription factors (Nanog and Oct4) essential to the pluripotency and self-renewal properties of undifferentiated stem cells. hASCs cultured with MesenPRO-RS™ medium alone (control) or supplemented with low doses of rhBMP4 (0.01 ng/mL) were collected after 6 days, and expression of Nanog and Oct-4 was analyzed by quantitative PCR. No significant statistical differences were observed between control and treated cultures.
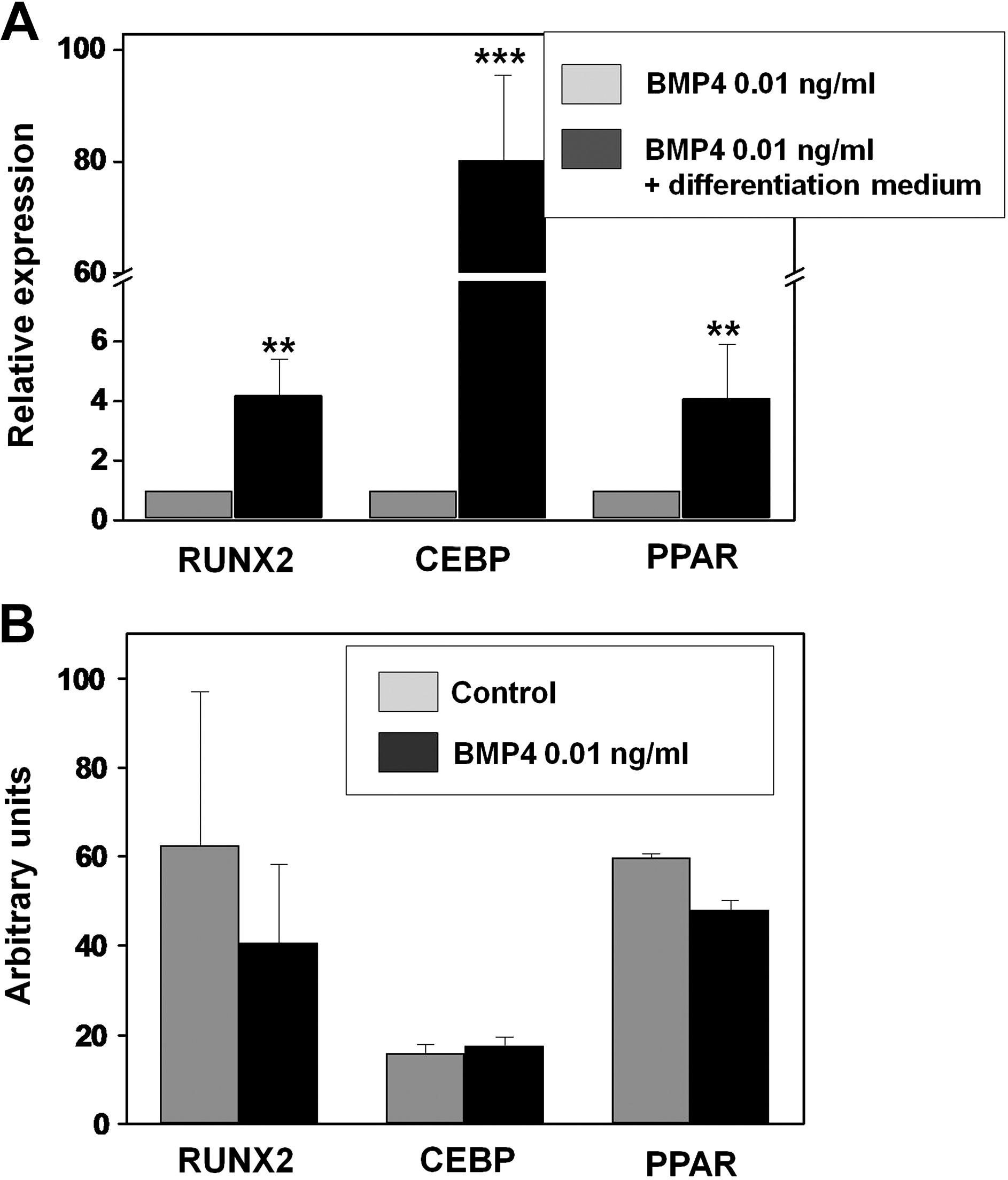
Adipose-derived stem cells pretreated with low doses of BMP4 maintain their ability to induce adipose and osteogenic gene expression.
Taken together, these results demonstrate the capacity of low doses of BMP4 to induce increased cell proliferation and survival of hASCs as well as to maintain their stemness and multipotent developmental properties.
Discussion
In the present study we have demonstrated that hASCs express all molecular machinery associated with the canonical BMP signaling pathway and respond to low doses of BMP4 increasing their cell proliferation and survival and maintaining their stemness and lineage multipotency properties.
Our results confirm that phenotype and growth rate of MSCs do not undergo variations when the cells are cultured in a medium with reduced amounts of serum, as other authors had observed [7]. The culture of MSCs in a medium devoid of serum or containing very few components is a necessary condition to understand the role played by BMPs in the biology of MSCs, one of the aims of our study. The medium MesenPRO-RS™ used in the present study is a culture medium containing very few components that does not include BMP4, as shown by the lack of this morphogen in the supernatants of fibroblast cultures (Fig. 3). On the contrary, normal serum contains BMP4 ranging from 3 to 5 ng/mL [34]. Thus, in these culture conditions, the presence of BMP4 in the culture supernatants confirms that hASCs endogenously produce BMP4 (Fig. 3). Previously, Halvorsen et al. [19] had reported expression of BMP2, BMP4, Alk3, Alk6, and BMPR-II in hASCs cultured in the medium with serum during 6 or 18 days. That expression was maintained even when the cells were cultured under adipogenic or osteogenic conditions.
On the other hand, our results demonstrate not only that hASCs express BMP receptors, specific (Smad 1 and Smad 5) and common transducers (Smad 4) of BMP signaling pathway, but also that stimulation of hASCs with low doses of BMP4 results in upregulated expression of transcription factors Id 1–4, targets of this pathway [25]. Other authors had described expression of BMP ligands [7,16 –20], receptors [7,19,21 –23], and Smad 1 [7] in hMSCs, but this is the first evidence, to our information, that demonstrate the activation of transcription factors specific of BMP pathway after BMP4 treatment of hASCs.
This evidence is further supported by the observed effects of different doses of BMP4 on the growth, survival, and proliferation of cultured hASCs. We first found that dorsomorphin, a drug that blocks the BMP signaling, significantly diminished the numbers of hASCs yielded in treated cultures as compared to those untreated. Further, our results indicate that dorsomorphin-dependent decreased numbers of hASCs are related to the effects of BMPs on cell proliferation and survival. Thus, dorsomorphin treatment produces significant decreased numbers of cycling hASCs and increased apoptosis within this cell population. Other data suggest a role for BMPs in the survival of different stem cells [19,26,35,36]. Self-renewing activity of hematopoietic stem cells is compromised in BMP4 hypomorph mice that express decreased amount of BMP4 in the hematopoietic niche [26]. Recently, Solmesky and colleagues [7] used Noggin, a specific BMP antagonist, to demonstrate that decreased endogenous production of BMP reduces the human bone marrow MSC survival by inducing caspase-dependent apoptosis. The reduced cell viability induced by Noggin is concentration dependent in agreement with our results using different doses of dorsomorphin. On the other hand, BMP4 treatment provokes remarkably opposite effects on survival and proliferation of hASCs depending on the used doses: whereas low doses of BMP4 stimulate proliferation and survival of hASCs, resulting in increased numbers of cells in the culture, high ones decrease the numbers of proliferating cells and increase the apoptotic hASCs. Other authors have indirectly reported similar results. Thus, decreased MSC survival induced by Noggin treatment is improved in the presence of BMP4, but the cell survival in presence of BMP4 alone is similar to that found in cultures supplied with Noggin [7], suggesting that increased levels of BMP result in cell death. Further, high doses of BMP2 (100 ng/mL) reduce the MSC growth [29].
We finally analyzed whether the effects of BMP4 on proliferation and survival of hASCs affected their properties of self-renewal and multipotency. By using quantitative PCR to evaluate expression of different transcription factors that provide to stem cells their properties, Nanog and Oct 4, or are indicative for their commitment to adipose (CEBP and PPARγ) or osteogenic lineage (RUNX2), we demonstrated that hASC cultures supplied with low doses of BMP4 maintained their stemness, as shown by expression of both Nanog and Oct 4, as well as their capacity to differentiate into either adipose cells or osteoblasts when adequate inductive culture medium is supplied. Expression of embryonic stem cell markers has been previously reported in adult human MSCs from bone marrow [37], and BMP4, together with other factors, has been pointed out to be important for the maintenance of murine embryonic stem cell pluripotency [9]. About the BMP-dependent commitment of MSCs to different cell lineages, results are really controversial. A short exposure of MSCs to BMP2 seems to be sufficient to trigger osteogenesis [38], but other results report that BMP2, BMP4, and BMP7 promote preadipocyte differentiation [39 –41]. Recently [42], it has been described that BMP2, BMP4, BMP6, BMP7, and BMP9 effectively induce both adipogenic and osteogenic differentiation of MSCs in vitro and in vivo, but the specific commitment to one or another cell lineage is mutually exclusive.
In summary, our results support that endogenous production of BMP is an important factor for governing biology of hASCs. In addition, low doses of BMP4 could be used to expand and to extend the survival of hASCs maintaining at the same time their stemness properties and the multipotent capacities to differentiate to different mesoderm-derived cell lineages. These are important results to improve the cell numbers and survival of MSCs necessary to be used in cell therapy, since the greater ability of the exogenous cells to survive the greater the therapeutic response.
Footnotes
Acknowledgments
This work was supported by grants S-BIO/0204/2006 from Regional Government of Madrid; BFU2006-00651, BFU2007-65520, and BFU2009-10315 from the Ministry of Science and Innovation; RD06/0010/0003 from the Health Institute Carlos III; and GR58/08-910552 from the Complutense University and Regional Government of Madrid.
Author Disclosure Statement
No competing financial interests exist.