
Abstract
Transplantation of purified pluripotent stem cell-derived cardiomyocytes into damaged myocardium might become a therapy to improve contractile function after myocardial infarction. However, engraftment remains problematic. Aim of this study was to investigate whether murine embryonic fibroblasts (MEFs) support the functional integration of purified embryonic stem cell-derived cardiomyocytes (ES-CMs). Neonatal murine ventricular tissue slices were subjected to oxygen and glucose deprivation to simulate irreversible ischemia. Vital tissue slices served as control. Vital and avital tissue slices were cultured with or without MEFs before coculturing with clusters of puromycin-selected ES-CMs. Integration of ES-CM clusters was assessed morphologically, motility by long-term microscopy, and functional integration by isometric force measurements. We observed a good morphological integration into vital but a poor integration into avital slices. Adding MEFs improved morphological integration into irreversibly damaged slices and enabled purified ES-CMs to migrate and to confer force. We conclude that noncardiomyocytes like MEFs support morphological integration and force transmission of purified ES-CMs by enabling adhesion and migration.
Introduction
P
Currently, cardiac progenitor cells [4], and cardiomyocytes derived from ES cells (ES-CMs) [5] or from induced pluripotent cells generated by reprogramming of somatic human cells [6,7] or even from fibroblasts [8] represent the most promising cell source (CMs) for cardiac regenerative medicine. However, transplantation of these CMs seems to be more difficult than using fetal CMs [9,10]. One explanation might be that the CMs used in the proof-of-principle studies mentioned above used dissociated myocardium, that is, a mixture of cardiac cells including primarily CMs but also other cell types. In contrast, CMs derived from pluripotent stem cells have to be highly purified to avoid tumor formation [11,12].
Current knowledge on the mechanisms of cell integration and processes of physiological reconstitution as well as mechanical and electrical coupling after transplantation into host tissue is still fragmentary [13]. The reported beneficial effects of transplanted cells might be caused by numerous factors that are not related to the functional integration of contractile cells. For example, transplanted cells are known to produce paracrine factors that enhance survival of host CMs after infarction [14 –16]. Paracrine factors may also enhance angiogenesis, improve the extracellular matrix (ECM), or modulate the immune/inflammatory response [9,17].
Frequently, in vivo infarct models are too complex to allow meaningful conclusions especially on mechanisms of integration. Therefore, we used a recently developed in vitro tissue culture model to mimic cardiac cell therapy [18]. Viable ventricular tissue slices from murine neonatal hearts were damaged irreversibly by oxygen and glucose deprivation (OGD) to simulate severe ischemic injury [19,20]. Highly purified, puromycin-selected ES-CMs without tumor-forming progenitors were employed for in vitro transplantation [11].
There is cumulating evidence reporting beneficial effects of cell transplantation strategies combining CMs with other cell types like, for example, hematopoietic stem cell-derived cells [21], physiological constituents of myocardial tissue like endothelial cells [22] or fibroblasts [11]. Fibroblasts are found throughout cardiac tissue and account for up to two-thirds of the cells in the adult heart [23,24]. They are embedded within the ECM of the connective tissue and are, to a large extent, responsible for its synthesis. In diseased hearts, fibroblasts have central and dynamic roles in modulating cardiac function. The ECM and growth factors secreted from fibroblasts promote CM hypertrophy, leading to myocardial remodeling [25,26].
Previously, Kolossov et al. observed in an in vivo infarct model that purified ES-CMs showed an improved engraftment after transplantation when injected into injured myocardium together with an equal number of syngeneic fibroblasts [11] but did not provide information on underlying mechanisms. Previous own observations suggested that the nonpurified human ES-CMs [20] integrated better than purified murine ES-CMs. Most recently, Pfannkuche et al. [27] showed that purified ES-CMs alone were not capable to populate a 3D extracellular collagen type I matrix or to attach quantitatively to any of the numerous 2D biomaterials tested when cocultured without murine embryonic fibroblasts (MEFs). Although this study stressed the role of fibroblasts for integration of CMs into biomaterials, information on integrative properties into true myocardial tissue or even ischemically damaged myocardial tissue as well as data on the functional relevance of these observations are still lacking.
The aim of this study was therefore to analyze the engraftment of purified murine ES-CMs into vital and avital cardiac tissue without fibroblasts and in the presence of MEFs with special respect to migration and confer of force in the above-described in vitro model of simulated myocardial ischemia.
Materials and Methods
Preparation of vital murine neonatal ventricular slices
Generation of ventricular tissue slices was performed as described previously [18]. Briefly, neonatal murine ventricles were embedded in 4% low-melting agarose and sectioned into 300-μm-thick slices along the short axis with a microtome (Leica VT1000S; Leica Microsystems,
Oxygen and glucose deprivation
Avital slice were generated by simulated hypoxia and ischemia, due to OGD for 20 h, as described previously [20]. These avital ventricular slices lost their contractile function and could be further used as a dead natural and soft cardiac matrix tissue for in vitro transplantation experiments and recording of contractile function mediated by ES-CMs. OGD was performed by transferring the vital slices into a custom-made steel hypoxia chamber that was placed in a temperature bath maintained at 37°C. The chamber was filled with Tyrode's solution composed as described for the slicing procedure (with Ca2+ 1.5 mmol/L) with the exception that
Cell culture of MEFs
MEFs were generated as described previously [28]. Briefly, embryos (mouse strain Him:OF1) at embryonic day 14.5 were sacrificed and cut into small pieces after removing of bones, arms, legs, tail, and all inner organs. The tissue pieces were trypsinized, centrifuged, resuspended, and seeded into cell culture dishes with the medium composed of IMDM supplemented with 15% FCS. The cells were incubated at 37°C in 5% CO2. The medium was changed every 2 days. Cells were passaged up to 4 times in the ratio 1:4 when they were confluent.
Cell culture of murine ES cells and harvesting of purified beating ES-CM clusters
Purified ES-CM clusters were generated from transgenic ES cells (clone αPIG44) expressing puromycin resistance and enhanced green fluorescent protein (GFP) cassettes under control of a cardiac-specific promoter α-myosin heavy chain [11]. Undifferentiated ES cells were grown on MEFs inactivated by mitomycin C treatment (SERVA Electrophoresis GmbH) in the propagation medium composed of IMDM supplemented with 17% FCS, 100 U/mL penicillin, 100 μg/mL streptomycin, 1% nonessential amino acids (all purchased from Invitrogen GmbH,
Preincubation of avital ventricular slices with MEFs
For coculturing, we used custom-made dishes that contained a small funnel-shaped cavity as described previously [29]. After placing the slices into the funnel-shaped cavity, 10 μL of a cell suspension containing 100,000–150,000 MEFs was suspended over the slice. The small volumes were designed to maximize interactions between MEFs and the slices. After 30 min 400 μL and after 2 h of incubation another 3 mL of fresh IMDM supplemented with 20% FCS were gently added. The medium was changed every day. After 4 days of incubation the slices were coated with MEFs.
In vitro transplantation with purified ES-CM clusters
Avital slices with or without preincubation with MEFs were cocultured with purified ES-CM clusters again using funnel-shaped dishes. ES-CM clusters (17 ± 1) were placed partially beneath the slices providing direct contact to the ventricular slice. Videos and fluorescent images were acquired on days 1–7 of coculturing.
Integration score
To describe the morphological integrations that were visible during coculture a score was defined. This so-called integration score used bright field as well as fluorescent low-magnification microscopy to unequivocally identify the GFP-positive ES-CM clusters (Zeiss Axiovert 10; objective 2.5-fold or 4-fold) and video imaging to judge structural integration. Verbal descriptions and schematic drawings for pattern recognition were used for classification (Fig. 1). Observation during cautious wiggling was required to differentiate between “0” and “+”. Intrarater and interrater reliability of this score were high with intrarater reliability (Spearman) of 0.95 ± 0.08 and interrater reliability (Cronbach's Alpha) of 0.82. If the morphological integration of individual ES-CM clusters differed within one preparation, the cluster exhibiting the best integration was used for scoring.
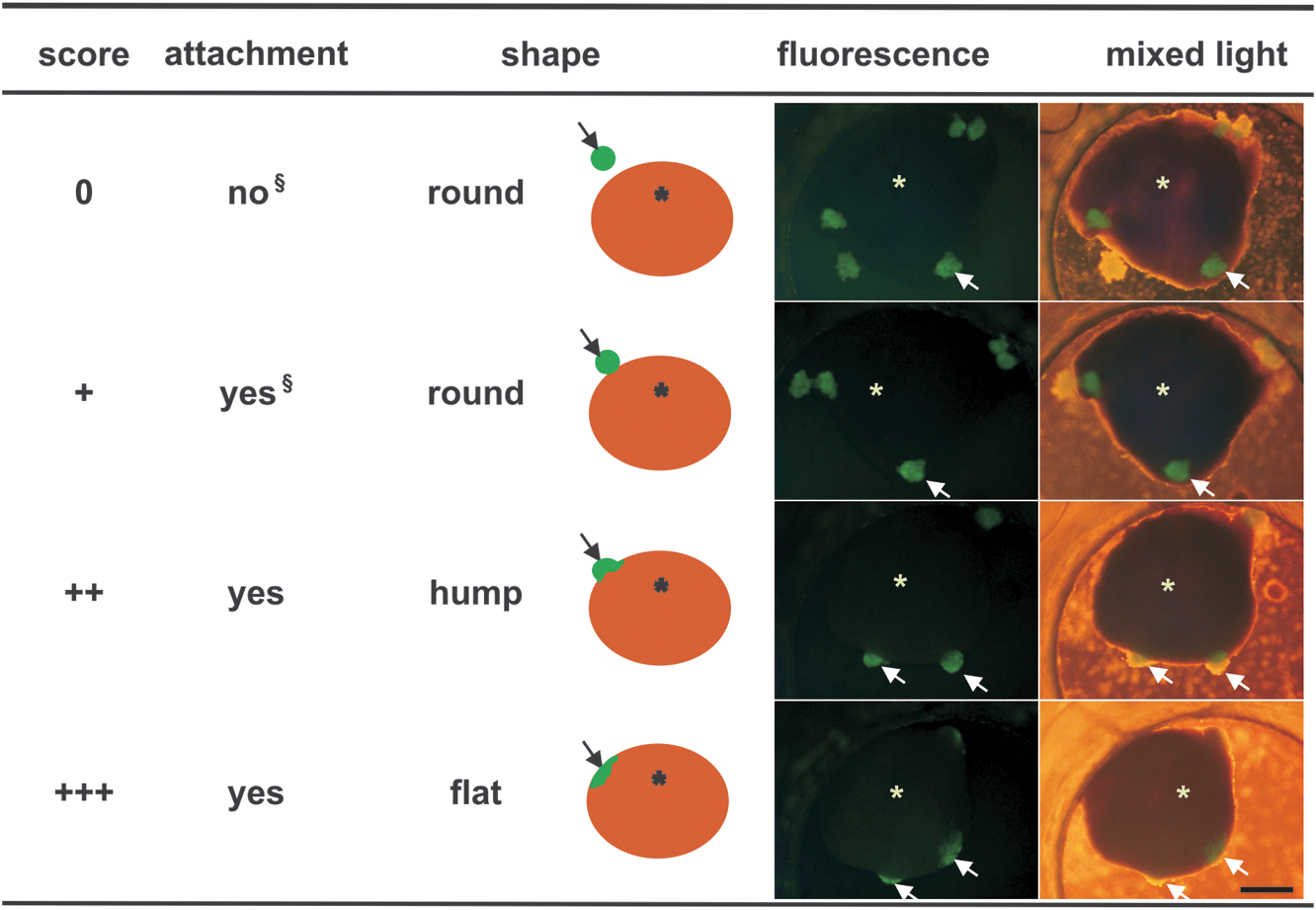
Integration score for transplanted ES-CM clusters. Criteria for the classification of integration. Arrows point to the fluorescent ES-CM cluster; asterisks and ocher circular disks mark the myocardial tissue slice. Observation during cautious wiggling is required to differentiate between “0” and “ + ”. Scale bar = 500 μm. ES-CM, embryonic stem cell-derived cardiomyocyte.
Time-lapse microscopy and velocity analysis of migratory activity
ES-CM migration in the absence or presence of MEFs was observed using time-lapse microscopy. Petri dishes (35 mm tissue culture grade) were coated with fibronectin (100 μg/mL; Millipore) for at least 30 min at 37°C with slight swirling. Then, the liquid was removed and the dishes were dried for 30 min. ES-CM clusters were selected for at least 4 days in the presence of puromycin. The clusters were trypsinized for 10–15 min and were washed twice with PBS. The cell suspension was filtrated (40 μm Cell Strainer; BD Biosciences) to remove nondissociated clusters. Cell number was evaluated with an electronic cell counter (Casy 1; Innovatis AG,
Histology and immunohistochemistry
For immunostaining, preparations were fixed for 20 min in 99.8% methanol at −20°C or 120 min in 4% paraformaldehyde at room temperature, washed with PBS, and incubated in 18% sucrose overnight before embedding in Tissue Tek OCT (Sakura Finetek Japan Co., Ltd.,
Isometric force measurements
After coculturing for 7–14 days, preparations consisted of the irreversibly ischemically damaged, noncontractile myocardial tissue, MEFs, and ES-CM clusters. They were mounted on an isometric force transducer (Scientific Instruments). Since slicing the hearts orthogonal to the long axis resulted in a tissue ring; the cavity of the left ventricle provided a preformed hole that eased mounting of the preparations onto the tips of 2 adjacent steel needles. This setup is very similar to the one used for force measurements of vascular rings. Contractions were recorded from spontaneously beating as well as from electrically stimulated preparations. Field stimulation was performed by silver electrodes (0.5–8 Hz, 5–15 V, stimulus pulse duration: 5 ms) connected to a custom-made stimulator. The preparation was maintained at 37°C and immersed in a dish filled with IMDM. Electrical stimuli and analog signals from the force transducer (KG7A; range, 0–5 mN; resolution, 0.2 μN; resonance frequency, 250–300 Hz) were amplified with a bridge amplifier (BAM7C; Scientific Instruments); analog signals were transferred to an A/D board and recorded as well as analyzed using the software DasyLab Version 7.0 (National Instruments).
Statistical analysis
All data are expressed as means ± SEM until otherwise stated. Force analysis was performed using paired t-tests. One-way analysis of variance was used for comparison of each group. Statistics were calculated using SPSS 11.0 (SPSS, Inc.). P values of <0.05 were considered significant.
Results
Integration score of transplanted ES-CM clusters
Integration was judged by the integration score as described before. After 7 days of coculture, in vitro transplanted ES-CM clusters attached, and eventually integrated completely into the vital slices (Fig. 2A and Supplementary Video S1; Supplementary Data are available online at
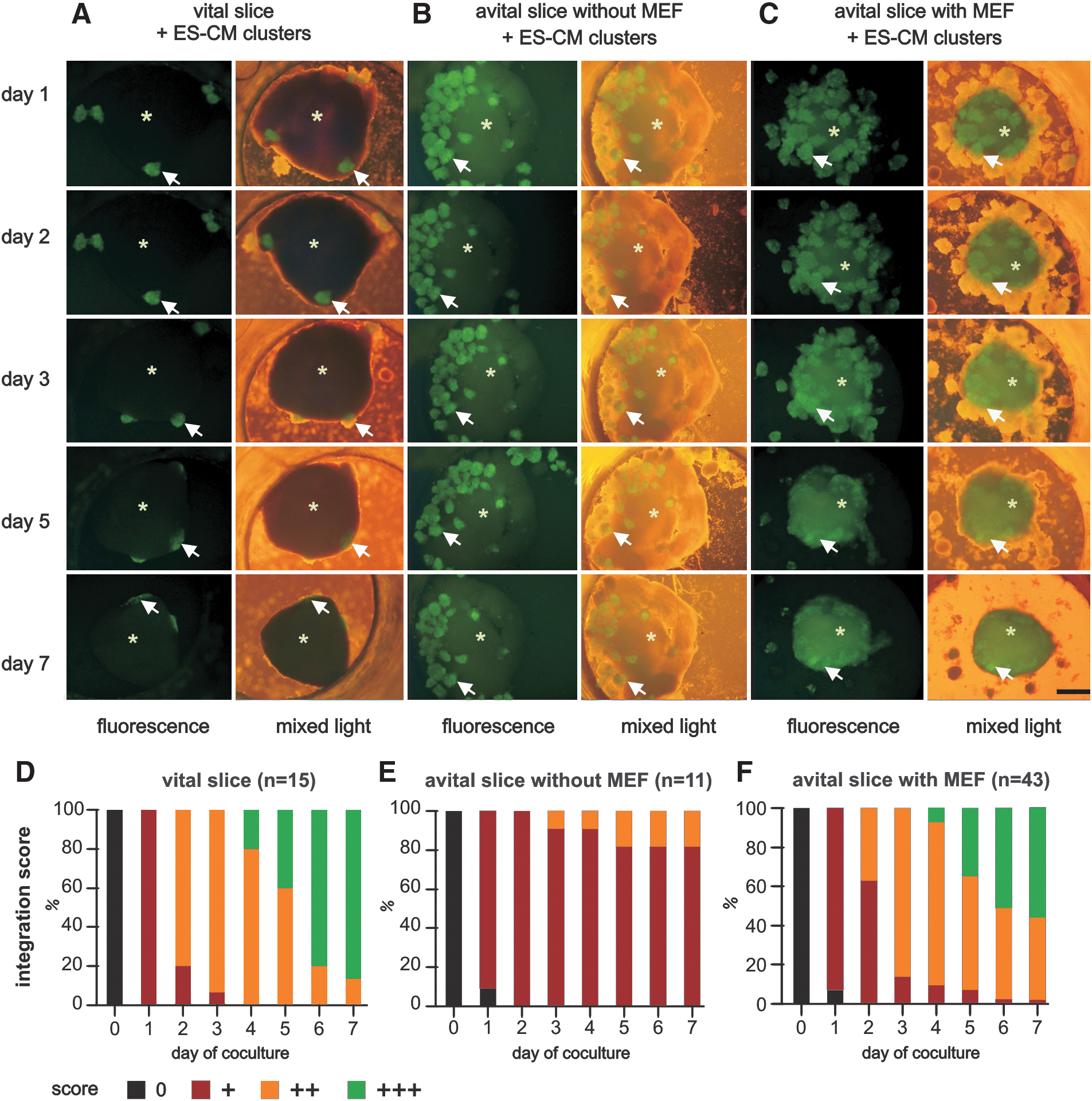
Sequential fluorescence microscopy and integration score for transplanted ES-CM cluster. Integration of GFP-positive ES-CM clusters (arrows) into a vital myocardial tissue slice
Morphological integration
Immunostaining was performed to elucidate the mechanisms causing the improved integration of ES-CM clusters into avital slices after preincubation with MEFs. Vital slices had well-preserved bright nuclei, indicating that the slices were morphologically intact. The vimentin-positive cells were detected foremost on the edge of the vital slices (Fig. 3A).

Immunohistochemistry and high-resolution electron microscopy.
Avital slices without any cocultured cells revealed no vimentin staining, demonstrating the absence of fibroblasts. The nuclei were blurred and fragmented or not detectable in the myocardial tissue subjected to 20 h of ischemia demonstrating cellular disintegration (Fig. 3B). Successful integration of MEFs into the avital tissue after preincubation was shown by positive vimentin staining (Fig. 3C). The colocalization of GFP-positive ES-CM clusters and vimentin-positive MEFs indicated a successful integration (Fig. 3D). Transplanted ES-CMs were located at the edge of the tissue and had clear cross-striations. Vimentin-positive fibroblasts surrounded the adjacent ES-CMs (Fig. 3E–H).
Ultrastructurally, we observed nonspecific adhesion contacts but no desmosomes or fasciae adhaerentes or other highly organized intercellular structures between the ES-CMs and MEFs (Fig. 3I–K). The transplanted ES-CMs exhibited an immature cardiac phenotype with poorly organized contractile elements.
Migratory activity of ES-CMs
ES-CM migration in the absence or presence of MEFs was observed using time-lapse microscopy. The images were captured with a camera every 5 min. For determination of velocity of ES-CMs in the presence of MEFs, 14 cells with migratory activity were chosen. The start and end positions of the cell were defined when the cells were at the peak of migratory activity. Purified GFP-positive ES-CMs without MEFs showed no motility at all (Fig. 4A–C), whereas purified GFP-positive ES-CMs with MEFs demonstrated a high motility with a velocity of 10.3 ± 1.4 μm/h (Fig. 4A–D). The motility of the ES-CMs was caused by a piggyback style movement of cardiac myocytes on fibroblasts. This is displayed in an exemplary sequence of a time-lapse video, where the movement of ES-CMs is indicated and the attachment to MEFs is evident (Fig. 4D). Moving pattern of the ES-CM–MEF complex seems to be random with unpredictable changes of direction.

Migratory activity of ES-CMs in the presence or absence of MEFs.
Force measurements
Since contractility is the pivotal quality of CMs, we analyzed whether ES-CMs are able to confer force of contraction in our in vitro transplantation model [20] at a defined developmental age (Fig. 5). Myocardial tissue subjected to 20 h of simulated ischemia by OGD without ES-CMs did not develop any force of contraction, even when stimulated with higher than physiological electrical impulses (20 V, 100 ms, n = 5) (Fig. 5 B). Cocultured preparations with ES-CMs without preincubation with MEFs were not able to confer force of contraction, because of limited integration into the ventricular slices (n = 5) (Fig. 5C). Cocultured preparations with ES-CMs after preincubation of the avital myocardial tissue with MEFs (Fig. 5D) developed and conferred force of contraction during spontaneous beating with a frequency of 1.7 ± 0.2 Hz with an amplitude of 1.8 ± 0.30 μN (n = 8). During electrical field stimulation with a frequency of 4 Hz, twitch amplitude increased to 3.2 ± 0.8 μN (n = 8, P < 0.05).

Force measurement. Representative force recordings of a vital myocardial slice without any coculturing
Discussion
Clusters of pure ES-CMs attach and integrate into vital myocardial tissue. In contrast, they are not able to do this or to confer force to myocardial tissue that has been subjected to irreversible injury by OGD before. Attachment, integration, and force transmission can be reconstituted by the presence of embryonic fibroblasts. Previously, Kolossov et al. observed in an in vivo infarct model that purified ES-CMs showed an improved engraftment after transplantation when injected into injured myocardium together with an equal number of syngeneic MEFs [11]. Therefore, we tested the functional relevance of MEFs as an enhancer of integration of purified ES-CMs.
Cardiac myocytes account for ∼3/4 of the normal adult myocardium, but for only 1/3 of cell numbers. The majority of the remaining myocardial cells are fibroblasts [24,30]. It is thought that cardiac fibroblasts originate from the proepicardium via epithelial-to-mesenchymal transformation [31 –33] and that the myocardial walls are later invaded by these epicardial-derived cells [34]. Cardiac fibroblasts are highly specialized cells that secret proteins of the ECM like different collagens, laminin, fibronectin, and perlecan. Synthesis and deposition of ECM proteins account for the elastic properties of the heart [35 –37] and play an important permissive role in cell survival and viability [38]. Fibronectin, collagens, and heparin-binding EGF-like growth factor secreted by murine embryonic cardiac fibroblasts enhance proliferation of cardiac progenitors [39]. Fibroblasts provide ECM support for developing CMs, but also possibly critical growth factors and cytokines, as reported for the in vitro differentiation of cardiac progenitor cells into CMs [4].
However, ECM proteins without vital fibroblasts improved the success of transplantation in vivo only marginally [2,38] and biomaterials simulating the ECM were shown to be unable to support integration [27]. Similarly, the ischemically damaged myocardium we used in our study should have a largely preserved ECM resembling the composition in vivo very closely. However, even this matrix was insufficient to support integration of cardiac myocytes. These observations stress the importance of factors that are not provided by an acellular ECM.
Our results provide evidence that functional integration into avital myocardial tissue and cellular engraftment deserves factors that most likely can only be provided by vital cells, for example, MEFs. Indeed, there is an emerging role of nonmyocytes for heart development significantly exceeding the synthesis of ECM proteins. The role of cardiac fibroblasts for growth and expansion of the ventricular chambers during heart development has just been recognized [37,39]. Possible mechanisms include secretion of fibronectin, collagen, and heparin-binding EGF-like growth factor, which collaboratively promote CM proliferation through b1 integrin signaling [39]. Irrespective of the mechanism, the presence of vital cellular elements seems to be of crucial importance for enhancing the retention and survival of transplanted ES-CMs far beyond producing ECM proteins.
Cardiac fibroblasts are known to interact with CMs [40] and to be capable of coupling electrically to the surrounding tissue via connexins [41]. In addition to these paracrine, ECM-mediated, and connexin-mediated effects, there seems to be a component supporting migration. Our observation of a piggyback style movement of cardiac myocytes on fibroblasts is another example for a quality that can only be achieved by vital interacting cells. It seems to be the prerequisite for migration of CMs that seemingly do not possess this ability at this stage of differentiation. In our preparations with avital tissue slices MEFs surrounded the integrated ES-CM clusters. Cell therapies need to replace injured cells and therefore transplanted cells have to migrate into injured tissue. To assess their migratory behavior we cultured ES-CMs as pure populations and observed their motility by long-term microscopy but could not show any motility of pure ES-CMs cultured on fibronectin, probably one of the most potent ECM components [27]. Obviously, purified ES-CMs are highly specialized cells that alone lack the ability to migrate. In contrast, when cocultured with MEFs they were highly motile [33,42].
Since, eventually, the goal of cardiac cellular replacement strategies is to improve the contractile function of the heart, we tested for mechanical integration and demonstrated that purified ES-CMs were only able to confer force of contraction when the slices had been preincubated with MEFs. Cardiac fibroblasts are known to couple mechanically via ECM receptors like integrins [43]. Ultrastructurally, we observed nonspecific adhesion contacts between purified ES-CMs and MEFs and no recognizable specific adhesion contacts such as desmosomes and fasciae adhaerentes. These nonspecific adhesion contacts between ES-CMs and MEFs and the ECM secreted by MEFs coupled ES-CMs effectively to the avital tissue. In conclusion, purified ES-CMs do not integrate into avital myocardial tissue and thus cannot transfer force of contraction. Vital cells like MEFs are necessary to enable migration of CMs and thus cellular engraftment, adhesion, and confer of force to noncontractile tissue.
Future studies should aim at defining the optimal relation of CMs to nonmyocytes and to characterize the properties of embryonic fibroblasts. In addition, from a translational point of view, other quite promising and well-accessible cells sources (eg, mesenchymal or endothelial stem cells) should be tested as an alternative to fibroblasts to improve cardiac cell replacement therapy.
Footnotes
Acknowledgments
This work was supported by the Koeln Fortune Program/Faculty of Medicine, University of Cologne: grant 157/2003 (F.P.), grant 154/2007 (T.H.), and BMBF grant No. 01GN0947 (M.K.). The authors thank Annette Köster, Moritz Haustein, Rebecca Dieterich, Maria Grotgans, Cornelia Böttinger, Martin Rubach, and the technical workshop for excellent support, and Suzanne Wood for the secretarial assistance.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.