
Abstract
Patient-specific human induced pluripotent stem (hiPS) cells not only provide a promising tool for cellular disease models in general, but also open up the opportunity to establish cell-type-specific systems for personalized medicine. One of the crucial prerequisites for these strategies, however, is a fast and efficient reprogramming strategy from easy accessible somatic cell populations. Keratinocytes from plucked human hair had been introduced as a superior cell source for reprogramming purposes compared with the widely used skin fibroblasts. The starting cell population is, however, limited and thereby further optimization in terms of time, efficiency, and quality is inevitable. Here we show that rat embryonic fibroblasts (REFs) should replace mouse embryonic fibroblasts as feeder cells in the reprogramming process. REFs enable a significantly more efficient reprogramming procedure as shown by colony number and total amount of SSEA4-positive cells. We successfully produced keratinocyte-derived hiPS (k-hiPS) cells from various donors. The arising k-hiPS cells display the hallmarks of pluripotency such as expression of stem cell markers and differentiation into all 3 germ layers. The increased reprogramming efficiency using REFs as a feeder layer occurred independent of the proliferation rate in the parental keratinocytes and acts, at least in part, in a non-cell autonomous way by secreting factors known to facilitate pluripotency such as Tgfb1, Inhba and Grem1. Hence, we provide an easy to use and highly efficient reprogramming system that could be very useful for a broad application to generate human iPS cells.
Introduction
Nevertheless, the generation of iPS cells is limited due to the low reprogramming efficiency [1] (<0.5%) and long duration [10,11] needed for reprogramming lasting several weeks in contrast to nuclear transfer or cell fusion [6]. Reprogramming efficiencies were tried to be improved by either co-expression of additional factors [10] that enhance reprogramming, for example, by chromatin modulation [12], application of chemical compounds [13], or the use of synthetic modified mRNA [14]. Particularly for the generation of patient-derived hiPS cells, a high reprogramming efficiency in a reasonable time schedule is required. Therefore, the generation of new reprogramming protocols and strategies dealing with these requirements is crucial.
hiPS cells can be generated and cultivated on mitotically inactivated feeder cells such as mouse embryonic fibroblasts (MEFs) [5] or human foreskin fibroblasts (HFFs) [15]. Feeder cells favor the process of reprogramming and the survival of the emerging stem cells via the supply of cell-to-cell contacts and the secretion of growth factors [16 –18]. Among these factors, Tgfb1, Inhba, Grem1, and Bmp4 have been reported as potential key candidates [19]. It has already been shown that these molecules are differentially secreted by MEFs after FGF2 stimulus and thereby facilitate human embryonic stem (ES) cell self-renewal [19]. This mechanism might also support the reprogramming process of somatic cells. Therefore, the type and quality of the feeder cells should have great impact on the efficiency of the reprogramming process [19]. Although human feeder cells, such as HFFs circumvent potential xeno-contaminations (eg, murine viruses and other xenobiotics), the reprogramming on HFFs remains rather inefficient and MEFs are currently the most widely used feeder cell for cellular reprogramming and hiPS cell culture [19].
We thus show that rat embryonic fibroblasts (REFs) can serve as a superior feeder cell layer compared with MEFs during reprogramming of human keratinocytes and HFFs. Arising iPS cell colonies were higher in absolute number when REFs were used instead of MEFs as feeder cell layers. This observation was further supported by an increase in absolute numbers of SSEA4 or NANOG-positive cells in REF-based reprogramming cultures. REF-based hiPS cells showed all major hallmarks of pluripotency. In addition, we found that potential candidate genes, described to be favoring stem cell survival [19], were upregulated (Tgfb1, Inhba, and Grem1) or downregulated (Bmp4) in REFs compared with MEFs, thereby pointing toward a partly non-cell autonomous mechanism. Together, we provide a very efficient reprogramming protocol for human keratinocytes, which we believe is useful for a broad application to generate human iPS cells in a large-scale format.
Materials and Methods
Keratinocyte cultures from plucked human hair
Outgrowth of keratinocytes from plucked human hair was induced essentially as previously described [7]. We used hair from 7 healthy volunteers (age between 24 to 45 and both male and female gender). Keratinocytes were consecutively cultured on 20 μg/mL collagen IV (Sigma-Aldrich, St. Louis, MO;
MEF and REF culture, HFFs, and FGF2 producing immortalized HFFs
CD-1 MEFs from embryonic day E12.5 or E14.5 (Stemcell Technologies, Vancouver, CA;
Human iPS cell culture
hiPS cells were kept in an incubator with 5% O2 on feeder cells in Knockout DMEM supplemented with 20% Knockout Serum Replacement (Invitrogen), 2 mM GlutaMAX, 100 μM nonessential amino acids, 1% antibiotic–antimycotic, 100 μM β-mercaptoethanol, 50 μg/mL ascorbic acid, and 10 ng/mL FGF-2 and passaged mechanically or by scratching after collagenase-IV treatment. For later passages hiPS cells were transferred to Matrigel-coated dishes (BD Biosciences, Franklin Lakes, NJ;
For cultivation in conditioned medium, irradiated CF-1 MEFs or REFs were kept in the medium described above but with 5 ng/mL FGF-2. The medium was collected for 5 days, sterile filtered, and stored at 4°C. One day after seeding cells on Matrigel in mTeSR1 the medium was changed to a 50:50 mixture of mTeSR1 and conditioned medium. From the following day on, the cells were kept in the conditioned medium supplemented with an additional 5 ng/mL FGF-2. Passaging was performed mechanically.
Lentivirus generation
Lentivirus containing a polycistronic expression cassette encoding for Oct4, Sox2, Klf4, and c-Myc [20] was produced in 70% confluent 10 cm dishes with Lenti-X 293T cells (Clontech, Mountain View, CA;
Reprogramming of keratinocytes
For infection, up to 3×105 keratinocytes per well of a 6-well plate were infected with 5×107 proviral genome copies in EpiLife medium containing 8 μg/mL Polybrene (Sigma-Aldrich) at 2 sequent days. Another 24 h later cells were detached using TrypLE Express (Invitrogen) and distributed onto 3 wells of a 6-well plate with already attached irradiated feeder cells (3×105 cells per well irradiated with 30 Gy) in hiPS cell medium. Further on the cells were cultured in a 5% O2 incubator. The medium was changed daily until arising colonies were big enough for mechanical passaging at about 2–3 weeks after transduction. Colonies displaying a clear stem cell morphology were picked and transferred onto irradiated MEFs or Matrigel-coated plates for further passage.
Germ layer differentiation and teratoma formation
Several hiPS cell colonies were detached by dispase treatment and kept in a ultra-low attachment suspension culture flask (Corning, Corning, NY;
Immunocytochemistry and alkaline phosphatase staining
Immunofluorescence staining was performed according to [21,22] and the manufacturer's protocol using the StemLite Pluripotency Kit (Cell Signaling, Danvers, MA;
For alkaline phosphatase (AP) staining, cells were rinsed in phosphate-buffered saline (PBS) and fixed in 4% paraformaldehyde for 2 min. Cells were then stained in a buffer containing 0.3 mg/mL Nitro Blue Tetrazolium and 0.175 mg/mL 5-Bromo-4-chloro-3-indolyl phosphate p-toluidine salt for 15–30 min in the dark until a clear blue staining was visible. The staining reaction was stopped by washing with PBS.
Western blots
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis and western blots were carried out according to standard procedures [23,24]. The following antibodies were used: TGFB1 polyclonal rabbit antibody (1:400, PAB12738; Abnova, Taipei, Taiwan;
Quantitative one-step real-time reverse transcription-polymerase chain reaction
Quantitative one step real-time reverse transcription-polymerase chain reaction (qRT-PCR) was carried out according to [8,25] using the Rotor-Gene Q System (Qiagen, Hilden, Germany,
Gene expression microarray
Gene expression microarray was carried out using the Agilent Whole Human Genome Microarray Kit (4x44k microarray kit G4112F; Agilent Technologies, Santa Clara, CA;
Flow cytometry
Cells were detached by trypsin digestion and washed in buffer (2% FBS, 2 mM EDTA, and 20 mM Glucose in PBS). For intracellular staining, cells were fixed for 20 min in 2% PFA on ice and permeabilized with 0.5% saponin for 20 min on ice. Cells were resuspended in buffer containing primary antibody (SSEA4 antibody, 1:100, Cell Signaling, BrdU antibody, 1:200; BD Biosciences). After 30 min of shaking at 4°C the cells were washed twice, resuspended in buffer containing Alexa-647 secondary antibody (Invitrogen), and kept for 30 min at 4°C in the dark while shaking. Afterward the cells were again washed twice, resuspended in buffer, and filtered through a 100 μm filter. DAPI (5 μg/mL) was added just before analysis if unpermeabilized cells were used. Measurements were performed in an LSRII flow cytometer using the FACSDiva software (both BD Biosciences).
Karyotyping
Chromosome preparation of trypsinized keratinocyte-derived hiPS (k-hiPS) cells of passages 10–20 was performed according to standard procedures, and karyotyping was done after GTG-banding. A total of 38 metaphases were scored.
Bisulfite sequencing
Genomic DNA was isolated from different k-hiPSC lines using the QIAamp DNA Mini Kit (Qiagen). Three hundred nanograms was used as input for bisulfite conversion (EpiTect Bisulfite Kit; Qiagen). Fifty nanograms of converted DNA was used as a template for conventional nested PCRs amplifying regions of the OCT4 and NANOG promoters using primers described earlier [27]. Purified PCR products were TA-cloned into pCR2.1-TOPO (Invitrogen), sequenced, and analyzed as previously described [27].
Statistical analysis
Reprogramming efficiency was calculated by dividing the average count of AP-positive colonies per well by the initial number of cells plated, scaled to the fraction of cells replated in each well. All quantitative data are expressed as mean±standard error of the mean. Statistical comparisons were made by Dunnett's t-test or Student's t-test and considered significant if P values were <0.05 (*) and <0.01 (**). Calculations of significance were done with GraphPad Prism 5 (GraphPad Software Inc., La Jolla, CA;
Results
REFs as feeder cells during reprogramming of human keratinocytes
REFs were isolated and cultured similar to MEFs and showed a similar morphology (Fig. 1A, B) and growth behavior. To determine optimal conditions for the reprogramming on MEFs as a feeder layer, we initially tested MEFs of different origin (Supplementary Fig. S1; Supplementary Data are available online at
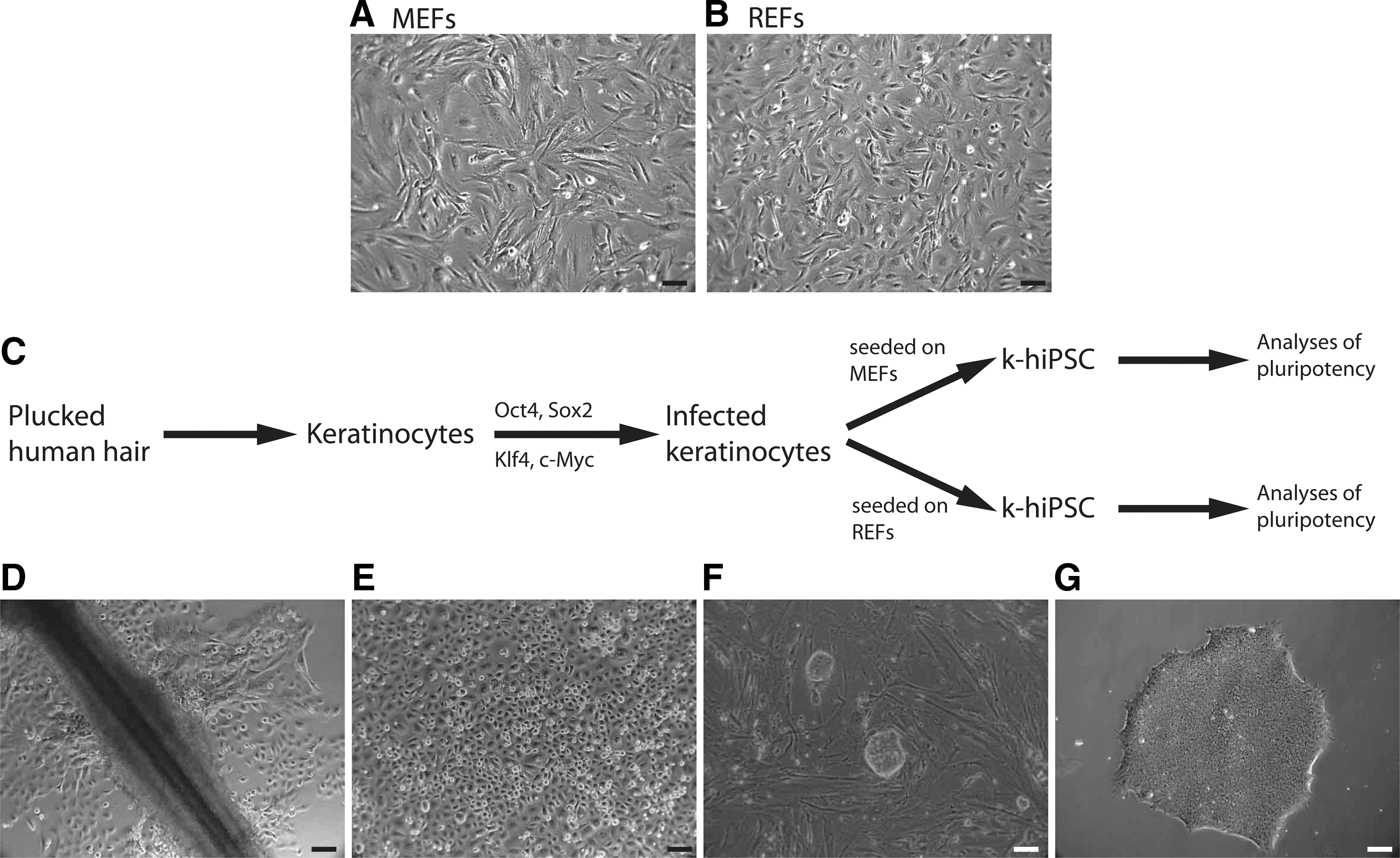
Human keratinocytes can be reprogrammed on MEFs and REFs using the same protocol.
REFs as a feeder layer improve reprogramming of keratinocytes
To directly compare the reprogramming efficiency of both feeder types, infected keratinocytes were seeded at identical cell numbers on either inactivated MEFs or REFs of the same passage. A few hours later, viable cells attached on both feeder cell types in comparable numbers. Seven to 8 days after infection, keratinocytes displayed strong alterations in cell morphology. They started to form tightly packed colonies that flattened during ongoing culture. Within 14–21 days colonies emerged that were big enough for mechanical passaging (Fig. 2A). On day 14, cultures were stained with AP (Fig. 2B). The total number of AP-positive colonies was about 3-fold higher on REFs compared with MEFs, leading to a reprogramming efficacy of ∼2.8% on REF feeder layers (Fig. 2C). To apply a more stringent assay for cellular reprogramming, namely, flow cytometry, we aimed to determine the amount of SSEA4-positive cells using either REFs or MEFs as feeder cell layer (Fig. 2D). On day 14 after infection the number of SSEA4-positive cells was about 2-fold higher in REF feeder layer-based cultures (Fig. 2E). There were no relevant differences in reprogramming performance between 4 REF preparations (Fig. 2C, E). In addition, REF-based reprogramming efficacies were independent of passage numbers (Supplementary Fig. S2). Reprogramming on REFs was superior to MEFs in all keratinocyte samples generated from 7 donors. In contrast, the use of FGF-2 producing immortalized HFFs that have been shown to be useful as feeder cells for higher hiPS cell passages [15] led to poor reprogramming efficiencies (below 0.001%, not shown).
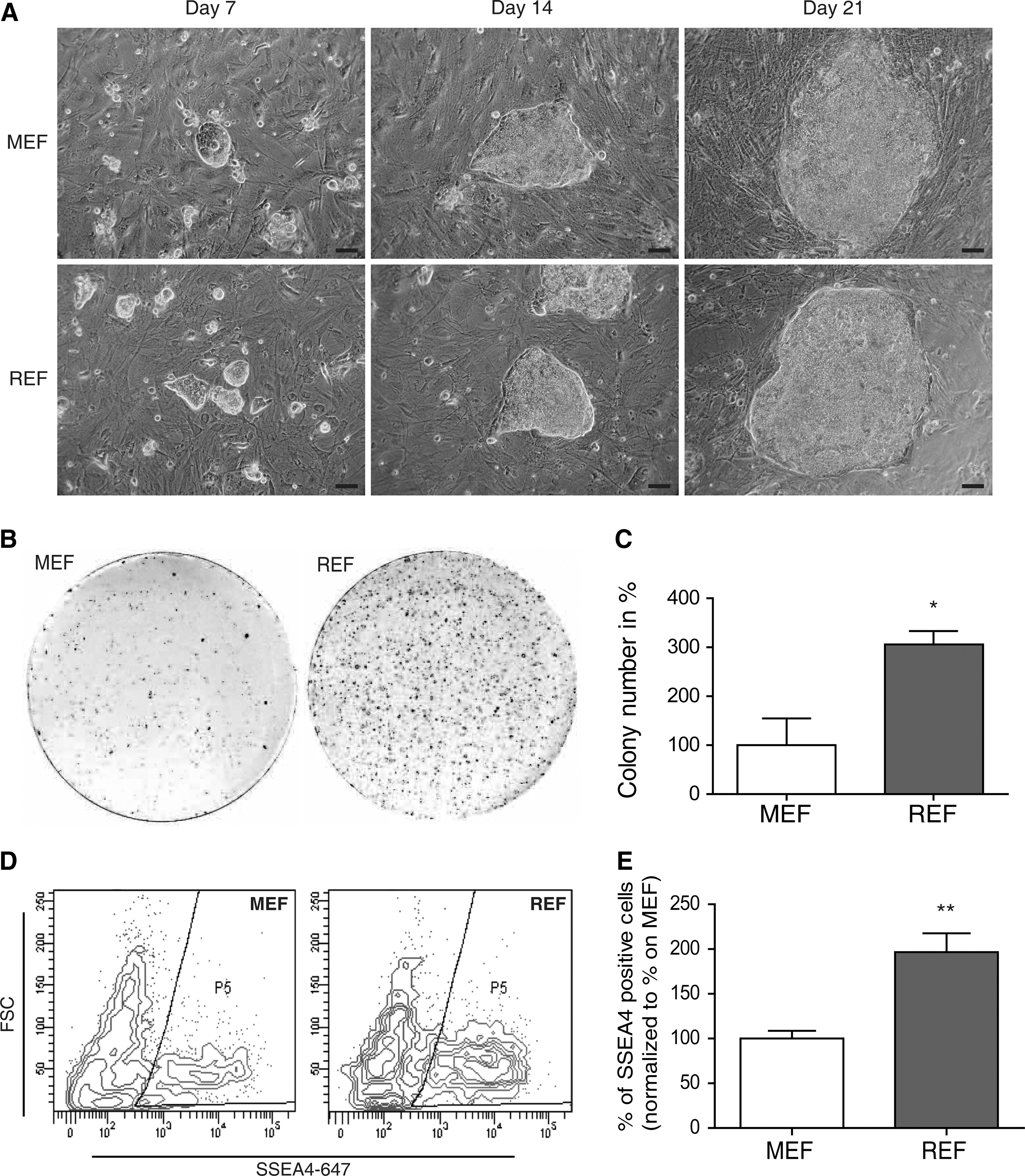
REFs have a significantly higher suitability as feeder cells for keratinocyte reprogramming compared with MEFs.
REF-based k-hiPS cells show all major pluripotency characteristics
Given the strongly enhanced reprogramming potency of REF feeder layers, we analyzed the quality of the generated k-hiPS cell clones according to established protocols (see Materials and Methods section). We generated numerous k-hiPS cell clones on REFs from different donors, which all harbored typical morphology and growth rates of human ES cells. All established clones stained positive for the stem cell markers OCT4, NANOG, SOX2, SSEA4, TRA-1-60, and TRA-1-81 in immunocytochemical analysis (Fig. 3A). In addition, all tested clones showed high endogenous expression levels for OCT4, SOX2, and NANOG, whereas KLF4 showed lower expression levels (also described in ref. [7]) when compared with keratinocytes (Fig. 3B). Global gene expression of k-hiPSC clustered close to hES cells and hiPS cells established elsewhere (Fig. 3C, D). Scatter plots analysis showed a high similarity between k-hiPSC and human ES cells, but not between k-hiPSC and keratinocytes (Fig. 3E). Thus, k-hiPSC are close to human embryonic stem cells at the global transcription level. Chromosome analysis of k-hiPSC showed that the majority (88%) has a normal chromosome set (Fig. 3F). As expected in fully reprogrammed hiPS cells, the promoter regions of the endogenous OCT4 and NANOG genes were fully demethylated (Fig. 3G) and expression of viral vector-encoded genes was silenced in our k-hiPS cell colonies (Fig. 3H).
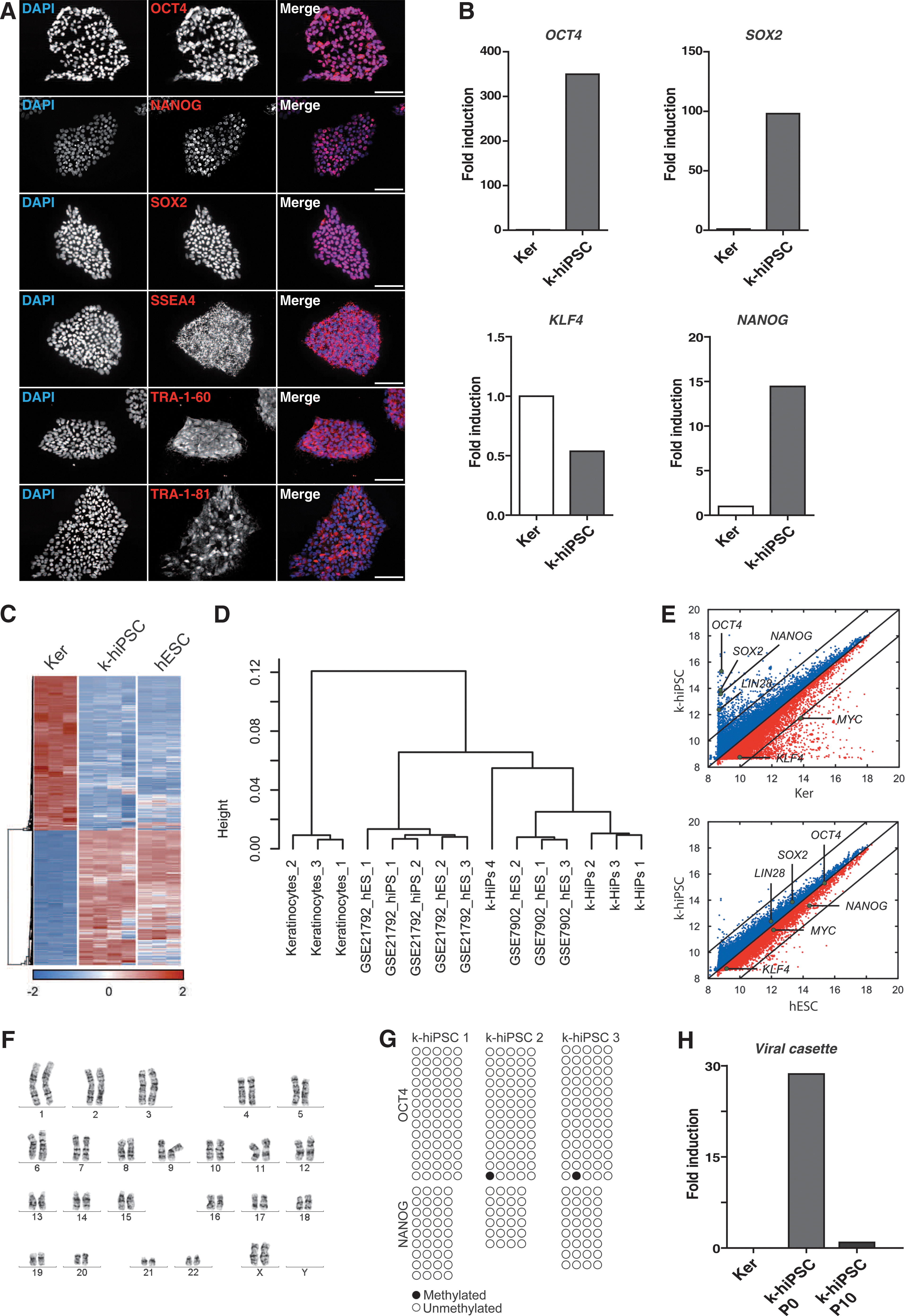
Stem cells produced on REFs as a feeder layer show major pluripotency characteristics.
REF-based k-hiPS cells differentiate into all 3 germ layers in vitro and in vivo
A hallmark feature of pluripotent stem cells is their potency to differentiate into all 3 germ layers. To confirm this differentiation pattern on protein level, marker proteins were analyzed by immunostaining. REF-based, k-hiPS cell-derived EBs were capable to form mesoderm and subsequently striated muscle cells as shown by α-actinin staining. Neuronal differentiation potential was confirmed by the neuronal marker β3-tubulin (TUBB3). In addition, α-feto-protein staining (AFP) depicted multi-nucleated cells pointing to endodermal, early hepatic differentiation of the cultures, representing the endoderm (Fig. 4A). mRNA expression analysis via quantitative RT-PCR (qRT-PCR) showed upregulation of marker genes for neural/ectoderm differentiation (PAX6, TUBB3), endodermal/hepatic differentiation (AFP, FOXA2), and mesoderm/cardiac differentiation (MYH6, T) (Fig. 4B) during EB differentiation.
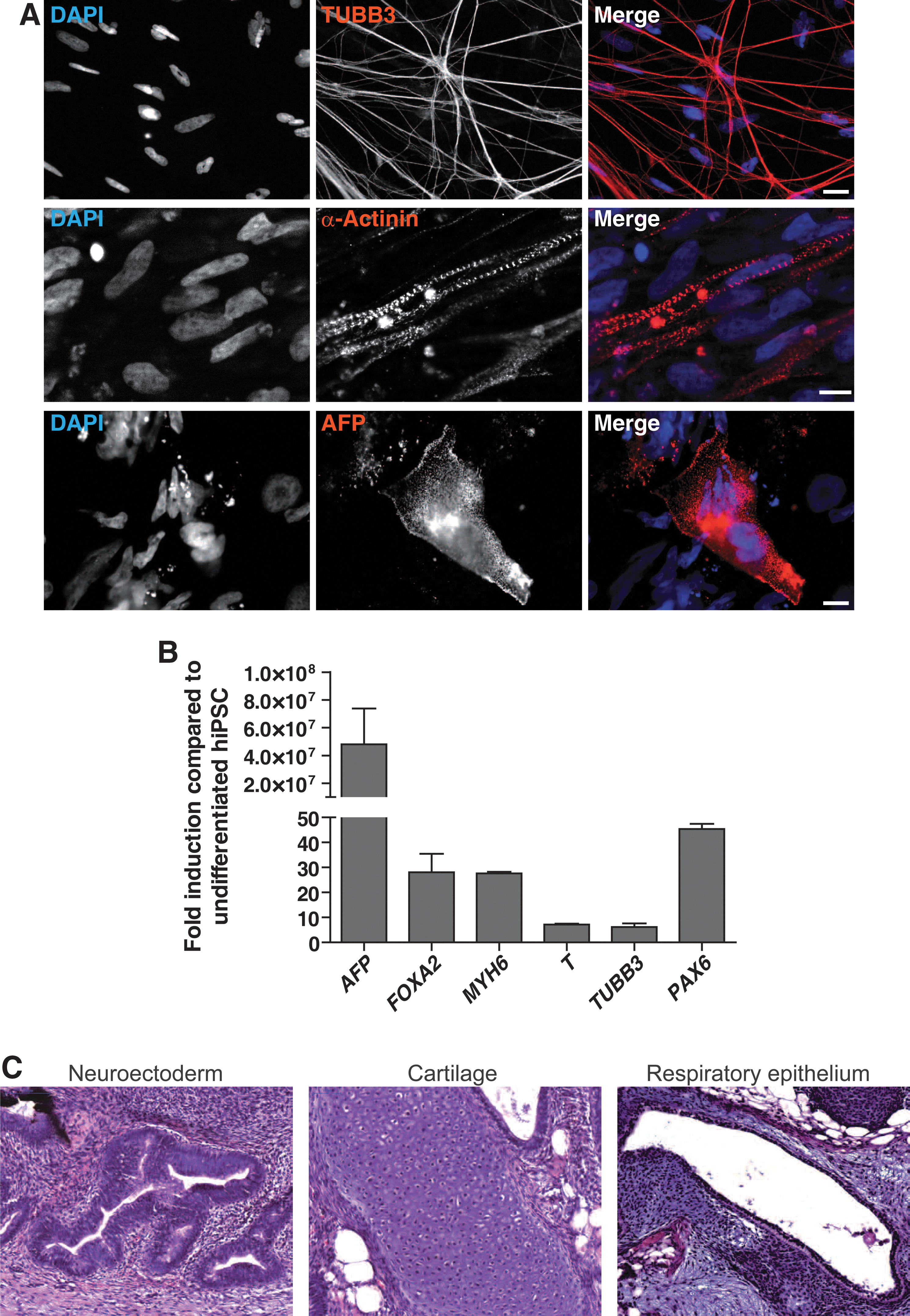
The stem cell colonies established on REFs are able to differentiate into cells of all 3 germ layers in vitro and in vivo. hiPS cell colonies were detached and kept in suspension for 10 days to form embryoid bodies. These were seeded on Matrigel-coated slides and stained after additional 14 days.
The in vivo potential of REF-based k-hiPS cells was assessed by teratoma formation in SCID mice, which contained tissues originating from all 3 germ layers (Fig. 4C).
REFs favor reprogramming and stem cell self-renewal non-cell autonomously
Proliferation rates have been shown to influence reprogramming efficiencies [10,29]. Although under debate, it has been proposed that proliferation rate is the major factor inducing stochastic events that lead to reprogramming [10,29,30]. To analyze whether differences in keratinocyte or k-hiPS cell proliferation due to different feeder layers could explain increased reprogramming efficiencies, proliferation rates were analyzed at early time points after virus infection of keratinocytes. BrdU labeling for 6 h revealed no differences in proliferation rates when being reprogrammed on MEFs or REFs (Fig. 5A). Another explanation for increased keratinocyte reprogramming on REFs could be a non-cell autonomous action such as secretion of specific factors enhancing hiPS formation. To that end culture medium was conditioned on REFs for 24 h and then used for reprogramming keratinocytes on MEFs. REF-conditioned medium enhanced the reprogramming on MEFs and led to an intermediate reprogramming efficiency between those on MEFs and REFs with fresh medium (Fig. 5B). In case of a non-cell autonomous action, REFs should be also able to enhance reprogramming of other cell types than keratinocytes. In this respect, we compared hiPS formation from HFF on MEF or REF feeder layers. In fact, REF-facilitated reprogramming was also superior when HFFs were used as parental cell type (Fig. 5C). Finally, we assessed the capacity of REFs to facilitate stem cell culture in the undifferentiated stage. Both REFs themselves and their conditioned medium enabled k-hiPS cells to be cultured for several passages without loss of pluripotency as assessed by morphology and OCT4 and NANOG protein expression (Fig. 5D, E; Supplementary Figs. S3 and S4).
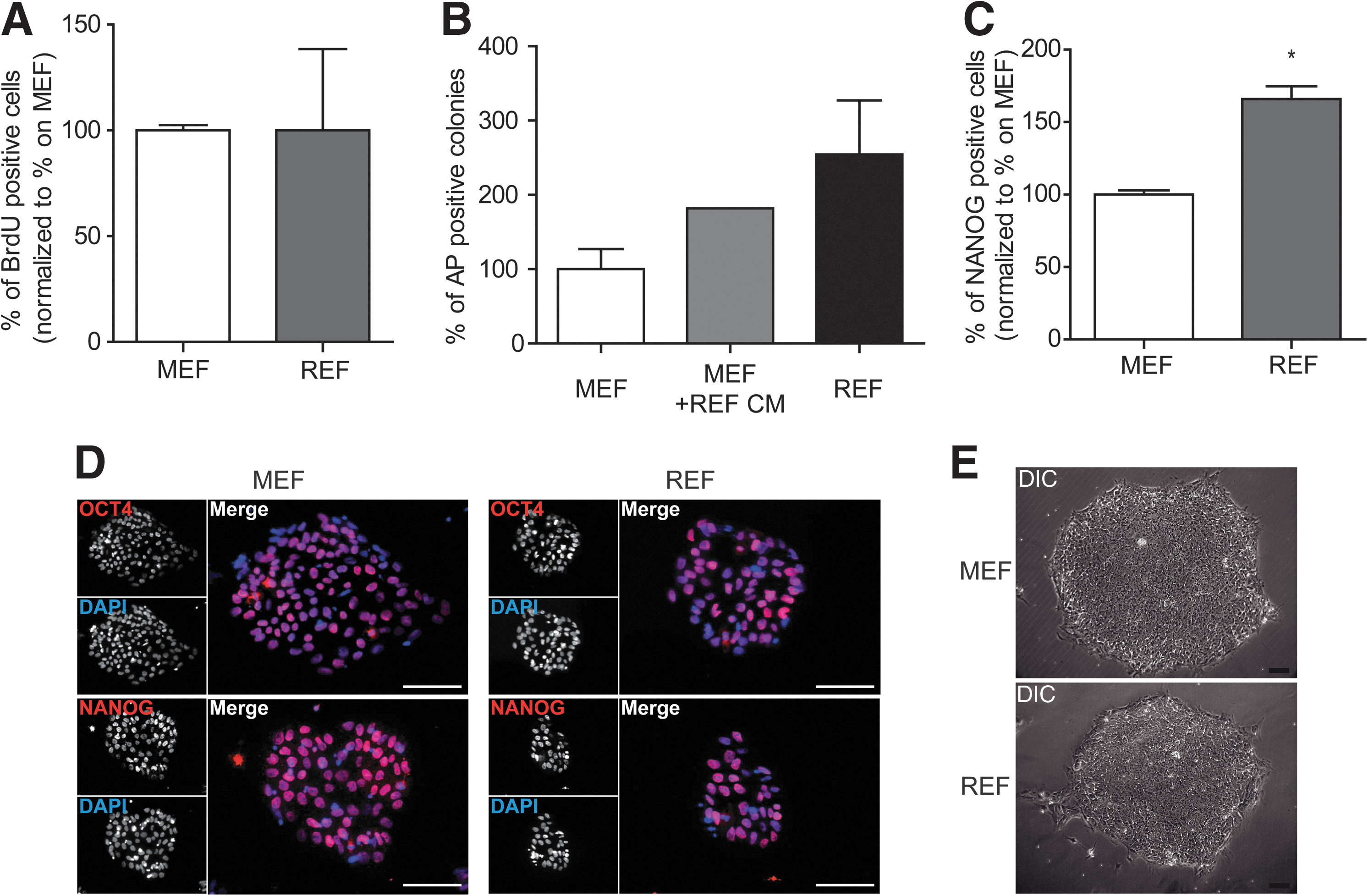
REFs favor reprogramming but not proliferation in a non-cell autonomous way.
REFs express distinct factors facilitating reprogramming and self-renewal
Having established several lines of evidence pointing toward a non-cell autonomous mechanism of REF-facilitated reprogramming, we aimed to identify at least some of these factors being present in REFs when compared with MEFs. It has been shown that TGFbeta signaling cascade is crucial for human pluripotent stem cell self-renewal, whereas BMP-signaling needs to be repressed to prevent differentiation [19]. qRT-PCR analysis of REFs growing in hiPS cell medium showed a significantly higher expression of key members of the TGFbeta pathway, Tgfb1, Inhba, and Grem1 compared with MEFs (Fig. 6A–C), whereas Bmp4, which favors differentiation, is downregulated (Fig. 6D). These observations could also be confirmed on protein level via western blot analysis of Tgfb1, Inhba, Grem1, and Bmp4 (Fig. 6E).
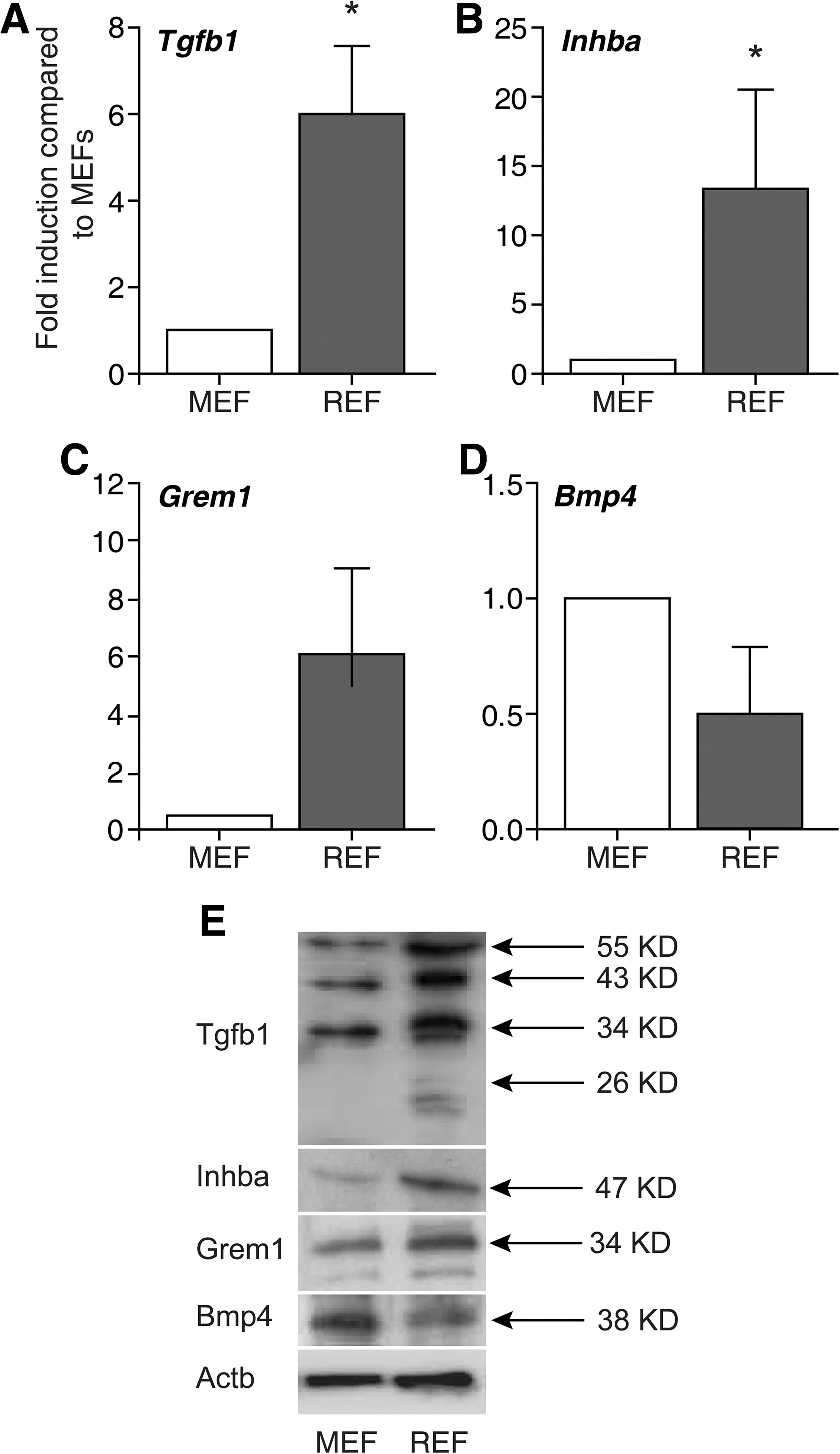
Analyses of potential candidate genes mediating the improvement of reprogramming by REF feeder layers by qRT-PCR and western blot. qRT-PCR show higher mRNA levels of the candidate factors for high reprogramming efficiency
Discussion
One of the most promising applications for hiPS cells is the acquisition of patient-specific stem cells and their use as a patient-specific disease model and readout system for pathophysiological and treatment research [31 –33]. Moreover, this novel technology provides individual cell systems for personalized medicine either by autologous cell replacement approaches or by person- and cell type-specific testing. The optimal tissue and cell type for reprogramming strategies is, however, still a matter of debate. Moreover, epigenetic memory preventing certain iPS cell lines to differentiate into a particular lineage has to be taken into account [34,35]. Most laboratories are currently using skin biopsy-derived fibroblasts that are highly proliferative and can be passaged over time. Nevertheless, reprogramming efficiencies remain low and the process is rather time-consuming and costly. Additionally, skin biopsies are more invasive compared with sampling plucked hair. Indeed, keratinocyte reprogramming has been reported to be 100-fold more efficient with reprogramming efficiencies of around 1% and 2-fold faster than fibroblast reprogramming despite using MEFs instead of our REF-based reprogramming system [7]. One of the reasons for the high reprogramming efficiencies in keratinocytes may be associated with transcriptional and epigenetic states favorable to reprogramming due to high endogenous levels of KLF4 and C-MYC in keratinocytes [7]. Similarly, a high reprogramming efficiency has been reported for neural stem cells, even when using only KLF4 and OCT4 as reprogramming factors [36]. Nevertheless, neural stem cells are doubtless not a broadly available cell source allowing the generation of patient-specific iPS cell libraries. Another, more noninvasive method favoring reprogramming is the usage of peripheral blood [37], but purification of the desired cell type is rather costly and difficult [38]. Cells that are easy to obtain are keratinocytes from the outer root sheath of human hair. This cell type is initially highly proliferative and can be propagated for several passages. To further improve reprogramming, we combined several known enhancements of the protocol to make it even more efficient such as the use of a polycistronic reprogramming construct [20,28], the use of low oxygen atmosphere [39], and the addition of ascorbic acid [40] or valproic acid [41]. However, most of these studies used feeder cell layers as a crucial factor for the survival and support of cells during the reprogramming process with MEFs being the most commonly used feeder cell type. Still, feeder cells display various and differential disadvantages. Indeed, MEF feeder cell lines are not only differing among preparations, but their quality also depends on age and strain of mice [19,42]. We identified REFs not only as an alternate and more stable feeder cell source but rather as an enhancer of human keratinocyte reprogramming. This has been shown by a significant increase in the number of reprogrammed cells and number of arising hiPS cell colonies on REF feeder layers with an estimated reprogramming efficacy of up to 2.8%. However, it is very difficult to compare reprogramming efficiency numbers between scientific groups, since there are a variety of different reprogramming protocols in use, including different vectors, virus amounts, feeder treatment, and, finally, experiment evaluation. To check if the reprogramming favoring effect of REFs is limited to keratinocytes, we reprogrammed HFFs and found that the efficiency was enhanced by more than 50% on REFs compared with MEFs. This indicates that the enhancement is not specific for the cell type being reprogrammed but for the used feeder cell layer. These data underline recent findings indicating that the environment of the cells being reprogrammed is crucial for their fate [43]. Therefore, REFs seem to produce a more suitable environment especially for early events in reprogramming.
To narrow down side effects in our culture system, we tested commercially produced MEFs to compare the REF-based system with standard MEF strains. The REF-based system was superior to all tested MEF batches and strains. Indeed, every REF preparation was successfully suitable for efficient reprogramming of keratinocytes pointing to increased reproducibility of REFs due to being less biased by culture conditions. REFs also maintained their reprogramming enhancing abilities over several passages. Although we did not test different reprogramming methods such as plasmid or RNA transfection, it is very likely that the use of REFs also eases reprogramming in other settings. Using REFs as feeder cells we successfully reprogrammed keratinocytes from several donors. The established k-hiPS cell lines showed the expected stem cell morphology and growth behavior, expressed all common stem cell markers, exhibited a gene expression profile very similar to hiPS and hES cell lines from other labs, and showed a mostly normal karyotype. Additionally, they had a silenced reprogramming cassette, demethylated promoters of typical stem cell marker genes, and could differentiate into various cell types in vitro and in vivo, comparable to stem cells obtained on MEFs. Further effort has been invested to delineate the mechanism behind the optimized reprogramming efficiency of REFs when compared with MEFs. Although under debate, it has been proposed that proliferation rate is the major factor inducing stochastic events that lead to reprogramming [10,29,30]. Nevertheless, REF-based reprogramming does not affect proliferation rates of the cells to be reprogrammed. Moreover, there seems to be a non-cell autonomous mechanism facilitating the reprogramming of keratinocytes by secreted factors. This has been shown by conditioned medium experiments and reprogramming of different cell types on REFs. We found that several genes, linked to self-renewal of hiPS cells [19], were expressed higher in REFs compared with MEFs, namely, Tgfb1, Inhba, and Grem1. Tgfb1 and Inhba code for secreted factors from the TGFβ pathway that enhance self-renewal through activation of SMAD2/3, whereas Grem1 inhibits Bmp4 signaling. On the other side, Bmp4 was downregulated in REFs compared with MEFs. This self-renewal favoring expression profile of REFs could well be the reason for their high suitability as feeder cells in the critical stages of reprogramming but also for long-term culture of hiPS cells. Future studies are warranted to finally confirm the responsible factors and/or the respective signaling pathways, allowing the generation of efficient reprogramming protocols without the use of feeder cell layers or—at least—without the use of xenobiotic material.
In summary, we provide a feeder cell system that is superior to the commonly used cell types employed for the support of human iPS cell reprogramming in terms of reprogramming efficiency. This system was generated to further facilitate the reprogramming of human plucked hair-derived keratinocytes. This unique complementation of a non-invasive to obtain cell source that harbors an endogenously high reprogramming potential together with a superior and more robust feeder system provides a pivotal prerequisite for reliable and fast generation of patient-specific k-hiPS cells to be applied in personalized medicine.
Footnotes
Acknowledgments
A.K. is supported by a fellowship provided by the Medical Faculty of Ulm University (Bausteinprogramm, L.SBR.0011). Main part of this study was funded by the Deutsche Forschungsgemeinschaft (DFG) (A.K. SFB 518 S.L., T.M.B. SFB 497/B8 and S.L., T.M.B. BO1718/4-1). The authors would like to thank Sabine Seltenheim, Renate Zienecker, Martina Bleidißel, and Boris Burr for their technical assistance.
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.