
Abstract
The enrichment of female germline stem cells (FGSCs) and the establishment of cell lines are influenced by the efficiency of cell purification. A previous study using mouse vasa homolog (MVH)-magnetic bead sorting for the isolation and purification of mouse FGSCs showed a relatively low efficiency. In this study, we tested 3 further proteins with the aim of improving the efficiency of FGSC purification. Immunofluorescence assays and magnetic sorting were performed using short-type pituitary gland and brain-cadherin (Stpb-c), CD9, and interferon-inducible transmembrane protein 3 (Iftm3, Fragilis), all of which are expressed in germ cells. Although all 3 proteins were expressed in FGSCs, CD9 was unsuitable because of its lack of germline specificity, and Stpb-c was also unsuitable because of the unavailability of an appropriate primary antibody. The efficiency of FGSC purification was remarkably enhanced using the germline-specific protein Fragilis, compared with that using MVH. This new method for the purification of FGSCs may have extensive applications in stem cell studies and clinical research.
Introduction
M
CD9 is a tetraspan protein that associates with several β1 integrins and plays an important role in sperm–egg binding and fusion [8]. In addition to being located on oocyte membranes, CD9 can also be detected in murine spermatogonial stem cells (SSCs) and embryonic stem cells [9 –11]. Short-type pituitary gland and brain-cadherin (Stpb-c) is a member of the cadherin family and contains transmembrane domains and 5 repeats of a cadherin motif in the extracellular domain [12]. This protein and its homeotic protein, long-type pituitary gland and brain-cadherin, are generated by alternative gene splicing. As an important signal molecule, Stpb-c plays a pivotal role in promoting the proliferation of SSCs in vivo and in vitro [13,14]. Fragilis has been reported to participate in PGC determination and is specifically expressed in germ cells [15]. It is composed of 137 amino acids with 2 transmembrane domains, and both the N- and C-terminal ends of Fragilis are located outside the cell [16]. Fragilis expression can be detected from the PGC determination stage through to germ cells in neonatal mice [16]. However, the expression of CD9, Stpb-c, and Fragilis in FGSCs and their potential applications in magnetic bead separation have not been reported.
In this study, we used immunofluorescence and magnetic bead-sorting assays to investigate CD9, Stpb-c, and Fragilis, with the aim of identifying a suitable marker to optimize the efficiency of FGSC purification. The results should improve our understanding of FGSCs, as well as aid the purification of FGSCs and the establishment of FGSC cell lines in vitro, with potential value for clinical medicine.
Materials and Methods
Animals
CD-1 mice were used in this study. All procedures for animals were approved by the Institutional Animal Care and Use Committee of Shanghai and were carried out in accordance with the National Research Council Guide for Care and Use of Laboratory Animals.
FGSC isolation and purification
Ovaries were collected from 5-day-old CD1 mice (Shanghai SLAC Laboratory Animal, China). Two-step enzyme digestion and FGSC sorting were performed as previously described [1], with minor modifications. Briefly, the duration of collagenase IV treatment was prolonged to 20 min, and the duration of trypsin digestion was reduced to 5–7 min. The cells obtained using the above method were incubated with polyclonal rabbit anti-mouse MVH (Abcam, ab13840), Fragilis primary antibody (Abcam, ab15592), or Stpb-c (Resgen) at room temperature for 30 min. The suspension was then centrifuged at 300 g min−1 for 5 min to collect the cell precipitate. After rinsing and resuspending in washing buffer [phosphate-buffered saline (PBS, pH 7.4) with 0.1% bovine serum albumin], sheep anti-IgG magnetic beads (Dynal Biotech) were added to the cell suspension and incubated at room temperature for 30 min. The mixture of cells and magnetic beads was placed on the magnetic bead separator for 2–3 min and the supernatant was aspirated. The fraction on the inner side of the eppendorf tube was rinsed twice with washing buffer and then resuspended in FGSC medium and cultured on mitotically inactivated SIM mouse embryo-derived thioguanine- and ouabain-resistant (STO) cell feeders (see later) in 24-well plates at 37°C under 5% CO2. The citrate elution step in the manufacturer's protocol was substituted by natural detachment; after culturing for 12–24 h, the magnetic beads spontaneously detached from the cells and were removed by the magnetic bead separator during the cell passage process. One microgram of primary antibody was added for every 105 gross cells, and correspondingly, 5×105 secondary antibody-coated magnetic beads were required.
Cell culture and growth
Cells sorted using MVH or Fragilis were cultured on mitotically inactivated STO cell feeders, using STO and FGSC culture medium, as previously described [1]. FGSCs were cultured on 96-well plates, and a growth curve was constructed based on the numbers of FGSCs at 24, 48, 72, 96, and 120 h. The initial concentrations of FGSCs sorted by Fragilis and MVH in the 96-well plates were 1.9×103 cells mL−1. Moreover, growth curves of Fragilis- and MVH-sorted cells were drawn as follows: cells isolated from 10 neonatal female mice were purified by Fragilis or MVH and aliquoted into three 96-well plates with initial concentrations of 4×103 cells mL−1 and 1.6×103 cell mL−1, respectively.
Immunofluorescence
Ovaries were collected from 5-, 10-, 20-, and 42-day-old mice or 5-day-old mice that were intraperitoneally injected with 5′-bromodeoxyuridine (BrdU; 50 mg/kg; Sigma) after 1 h for dual immunofluorescence. They were fixed in 4% neutral paraformaldehyde, processed through ascending concentrations of alcohol, and embedded in paraffin, using a standard protocol [17]. Sections were cut at 6 μm, dewaxed in xylene, rehydrated through descending concentrations of ethanol, cultured with 2 μM HCl at 37°C for 1 h, and rinsed 3 times with boric acid buffer (pH 8.5) for dual immunofluorescence. The sections were cultured with 0.25% trypsin at 37°C for 15 min and blocked with 10% goat serum at 37°C for 10 min. Polyclonal rabbit anti-mouse primary antibodies specific for MVH (Abcam), Stpb-c (Resgen), CD9 (BTC), and Fragilis (Abcam) were used at dilutions of 1:200, 1:150, 1:180, and 1:150, respectively. Mouse monoclonal anti-BrdU (1:200; Neomarker) was also used for dual immunofluorescence. Goat anti-rabbit or mouse secondary antibodies (Proteintech) labeled with fluorescein isothiocyanate or tetramethyl rhodamine isothiocyanate (TRITC) were diluted 1:150 or 200. Slides were mounted and analyzed using an Olympus fluorescence microscope.
For FGSC immunofluorescence, FGSCs were fixed in 4% paraformaldehyde for 15 min and rinsed twice with PBS (pH 7.4), followed by incubation with 3% H2O2 for 10 min, all performed at room temperature. After aspirating the H2O2 and rinsing twice with PBS, the samples were incubated with 10% goat serum at 37°C for 10 min. Primary polyclonal rabbit anti-Fragilis antibody (Abcam) or anti-Stpb-c antibody (Resgen) at 1:150 dilutions was added and the samples were incubated at 37°C for 1 h. After washing 3 times, samples were subsequently incubated with TRITC-labeled secondary antibodies at 1:150 dilution and with 4′-6-diamidino-2-phenylindole (100 ng/mL) for 30 and 15 min, respectively. After 2 washes, the cells were covered with PBS and visualized using a fluorescence microscope.
Reverse transcription–polymerase chain reaction
Total RNA from ovaries or FGSCs was extracted using Trizol reagent (Qiagen), according to the manufacturer's instructions. Approximately 2 μg of total RNA was used to synthesize cDNA with 20 U M-MLV reverse transcriptase, as previously described [1]. The primer sequences used are listed: Oct4 (NM_013633) F: AGCTGCTGAAGCAGAAGAGG, R: GGTTCTCATTGTTGTCGGCT; Dazl (NM_010021) F: GTGTGTCGAAGGGCTATGGAT, R: ACAGGCAGCTGATATCCAGTG; Nanog (AK010332) F: CAGGAGTTTGAGGGTAGCTC, R: CGGTTCATCATGGTACAGTC; Figla (NM_012013) F: CCAAAGAGCGTGAACGGATAA, R: TCTTCCAGAACACAGCCGAGT; glyceraldehyde 3-phosphate dehydrogenase (Gapdh) (NM_008084) F: GTCCCGTAGACAAAATGGTGA, R: TGCATTGCTGACAATCTTGAG.
The polymerase chain reaction (PCR) conditions for Oct4, Dazl, and Gapdh were 50 s at 94°C, 50 s at 54°C, and 30 s at 72°C, for 37 cycles. For Figla, the PCR conditions were 50 s at 94°C, 50 s at 57°C, and 20 s at 72°C, for 37 cycles. The PCR conditions for Nanog were 50 s at 94°C, 50 s at 50°C, and 30 s at 72°C, for 37 cycles.
Combined bisulfite restriction analysis
The methylation status of imprinting genes of FGSCs purified by Fragilis was determined by combined bisulfite restriction analysis. DNA of juvenile mouse ovaries or FGSCs was extracted with the DNeasy Tissue Kit, according to the manufacturer's protocol (Qiagen), and was treated by EZ DNA Methylation Kit (Zymo). Imprinted gene regions were amplified using specific primers (5′-TTAGTGGGGTATTTTTATTTGTATGG-3′ and 5′-AAATATCCTAAAAATACAAACTACACAA-3′ for Igf2r, 5′-GTAAAGTGATTGGTTTTGTATTTTTAAGTG-3′ and 5′-TTAATTACTCTCCTACAACTTTCCAAATT-3′ for Peg 10).
PCR products were purified by The E.Z.N.A.™ Gel Extraction Kit (Omega) and digested by restriction endonuclease AciI (NEB) and HhaI (Takara) at 37°C for 1 h, respectively.
Results
Expression of MVH, Stpb-c, CD9, and Fragilis in mouse ovaries
We investigated the expression of MVH, Stpb-c, CD9, and Fragilis in mouse ovaries from 5-, 10-, 20-, and 42-day-old female mice using immunofluorescence assays. MVH was exclusively expressed in all types of germ cells, with an intense signal in each age group (Fig. 1A–D). Stpb-c showed a similar expression pattern to that of MVH and was also detected in germ cells of all stages, from early germ cells in the cortex through to oocytes in the antral follicles (Fig. 1E–H).
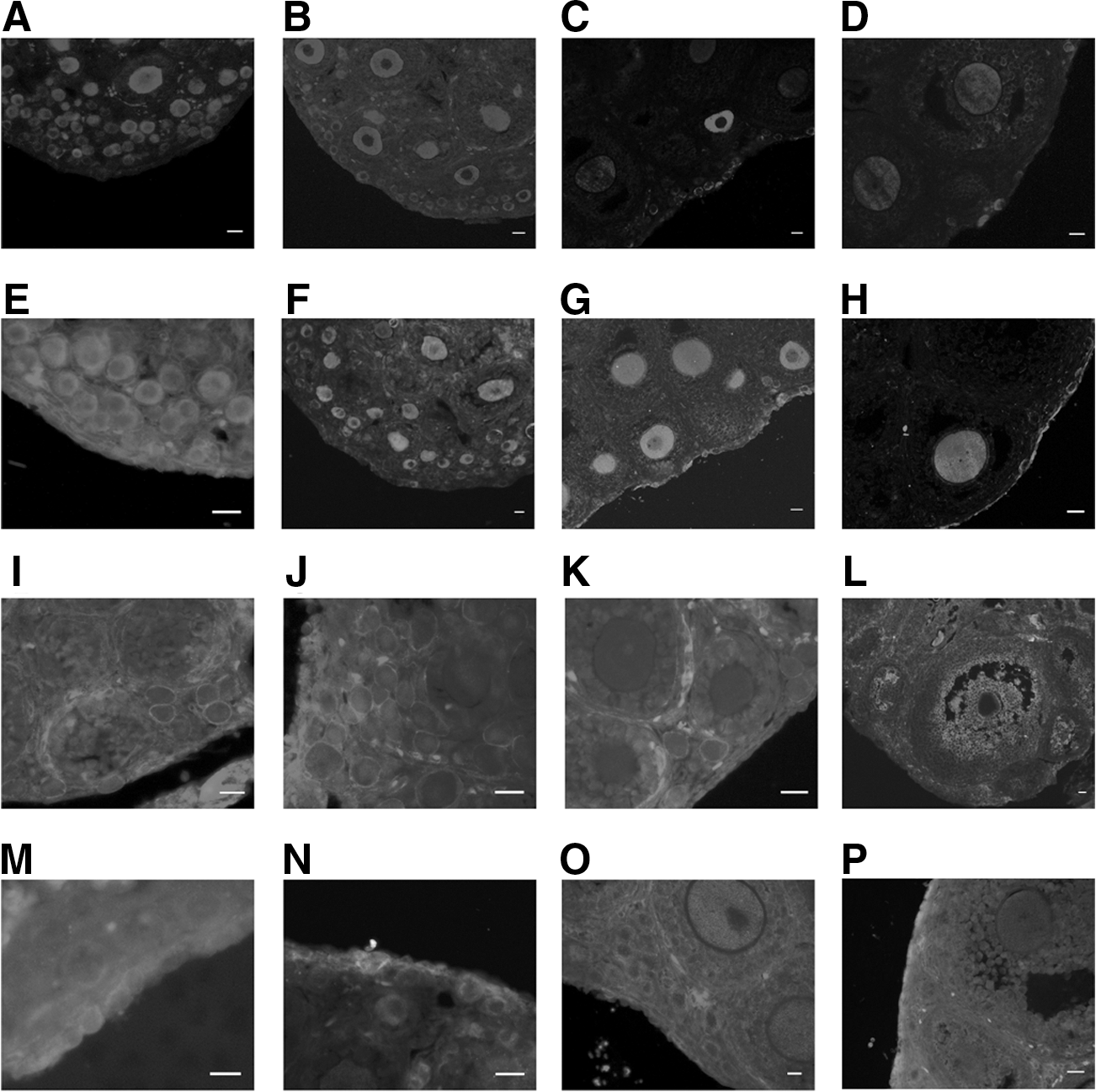
Expression of MVH, Stpb-c, CD9, and Fragilis in mouse ovaries.
The transmembrane proteins CD9 and Fragilis were detected from the early germ cell stage. As a crucial protein in gamete fusion, CD9 was mainly expressed in oocytes, and the signals in early germ cells were relatively weak (Fig. 1I–K). However, CD9 expression was also detected in the granulosa cells surrounding the oocytes (Fig. 1L). The expression level of Fragilis in early germ cells was high (Fig. 1M–P), whereas the signal in the oocytes of adult ovaries was much weaker. Moreover, the expression of Fragilis in the early stage of germ cells in the cortex was obviously stronger than that in oocytes (Fig. 1P). All 4 proteins were expressed from early germ cells through to mature oocytes, indicating their expression throughout the germline; MVH and Stpb-c were mainly localized to the cytoplasm, whereas CD9 and Fragilis were restricted to the membranes. Based on the comparative analysis of the expression of these proteins, we chose Stpb-c and Fragilis for subsequent FGSC sorting because of their specific expression in germ cells.
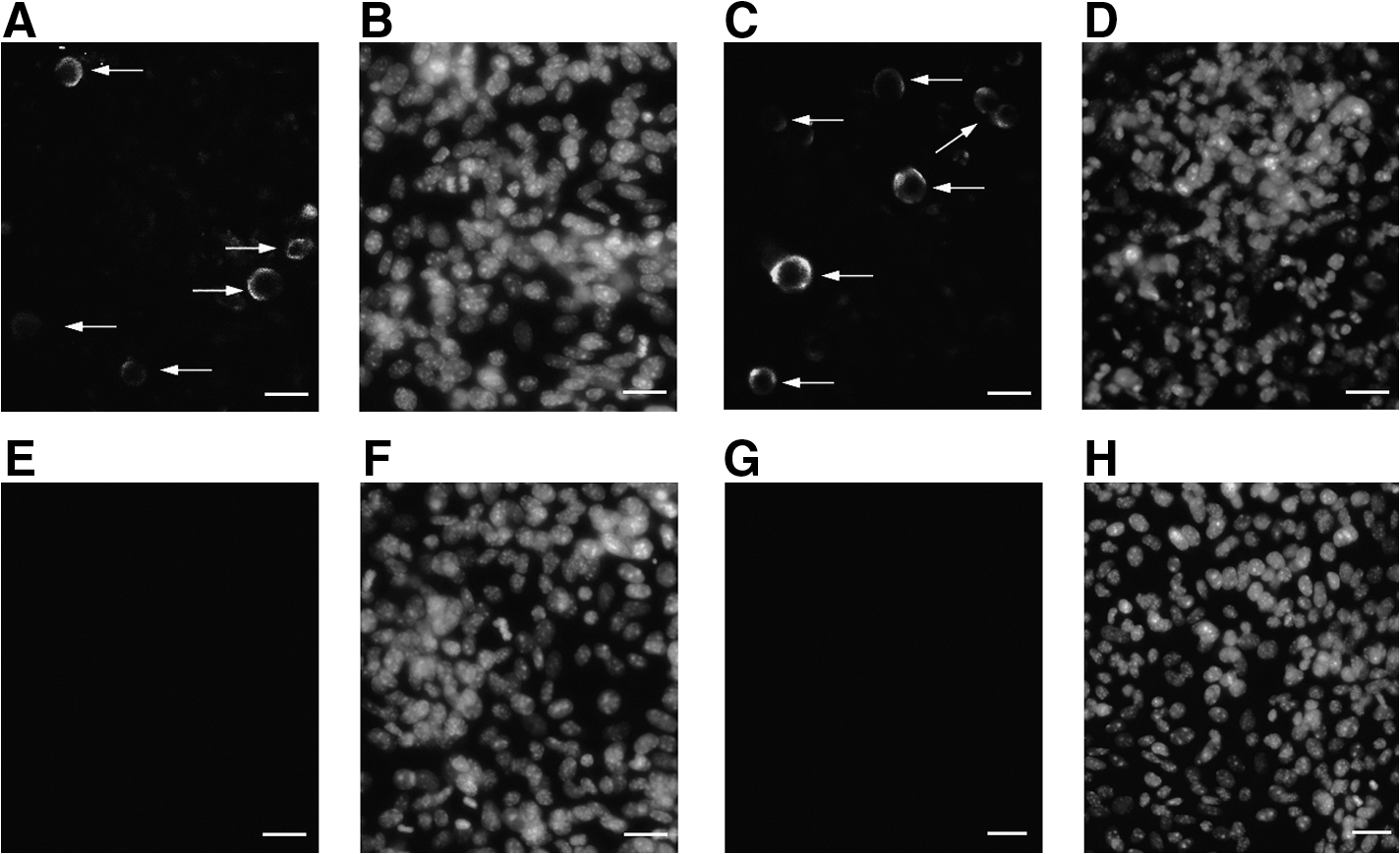
FGSC localization in vivo using dual immunofluorescence
Dual immunofluorescence assays were used to confirm the expression of Stpb-c and Fragilis in FGSCs in vivo. The typical characteristic of FGSCs is the simultaneous expression of the features of both stem cells and germline cells, with the implied capacities for self-renewal, differentiation, and production of the next generation. We therefore searched for cells positive for BrdU and Stpb-c/Fragilis in ovaries to localize the FGSCs. Some BrdU+/Fragilis+ and BrdU+/Stpb-c+ cells were observed in the cortex of 5-day-old ovaries (Fig. 2). Based on the results of our previous research, BrdU+/MVH+ cells at the edge of the ovary cortex are FGSCs. Because Stpb-c and Fragilis, like MVH, are also expressed from the early stage of germ cells, these BrdU+ early germ cells expressing Stpb-c or Fragilis were also implicated as FGSCs.

Dual immunofluorescence analysis of 5-day ovaries.
Fragilis expression in long-term–cultured FGSCs
Stpb-c and Fragilis expression in vivo was identified using dual immunofluorescence assays. To further determine the expression of these proteins in vitro, we tested their expression in FGSCs from our previously established FGSC line [1]. The FGSC line expressed both Stpb-c and Fragilis (Fig. 3). Overall, these results confirmed Stpb-c and Fragilis expression in FGSCs in vivo and in vitro and confirmed that these proteins were suitable for FGSC sorting.
Expression of Stpb-c and Fragilis in FGSC line.
Magnetic bead purification of FGSCs by MVH, Stpb-c, and Fragilis
Having verified that MVH, Stpb-c, and Fragilis were expressed in FGSCs in vivo and in vitro, we investigated the efficiencies of cell sorting using these proteins. Our previous study using the bioinformatics tool TMPRED predicted that MVH possesses 2 transmembrane domains and reported that about 300 germ cells could be collected from 20 ovaries from 5-day-old mice by MVH-targeted immunomagnetic bead sorting. The sorted product comprised FGSCs and oocytes [1]. The efficiency of isolation and purification of MVH-magnetic bead sorting was improved by minor modifications; prolongation of collagenase IV digestion, reduced trypsin treatment, and optimization of the proportions of cells, primary antibody, and magnetic beads resulted in the collection of 538±42 germ cells from 20 ovaries (values given as mean±standard error of the mean, based on the results of 5 experiments) (Fig. 4G–I). In contrast, MVH sorting using Stpb-c failed to collect any cells, either germ cells or somatic cells, and cells attached to magnetic beads were not detected under the microscope (data not shown). However, cell sorting using Fragilis resulted in the collection of 1,120±38 cells from 20 ovaries of 5-day-old mice (based on the results of 7 experiments) (Fig. 4A–C). These results suggest that immunomagnetic bead sorting using Fragilis greatly improved the efficiency of FGSC purification and almost doubled the output compared with that using MVH (P<0.001). Stpb-c was excluded because of the failure to sort targeted cells.
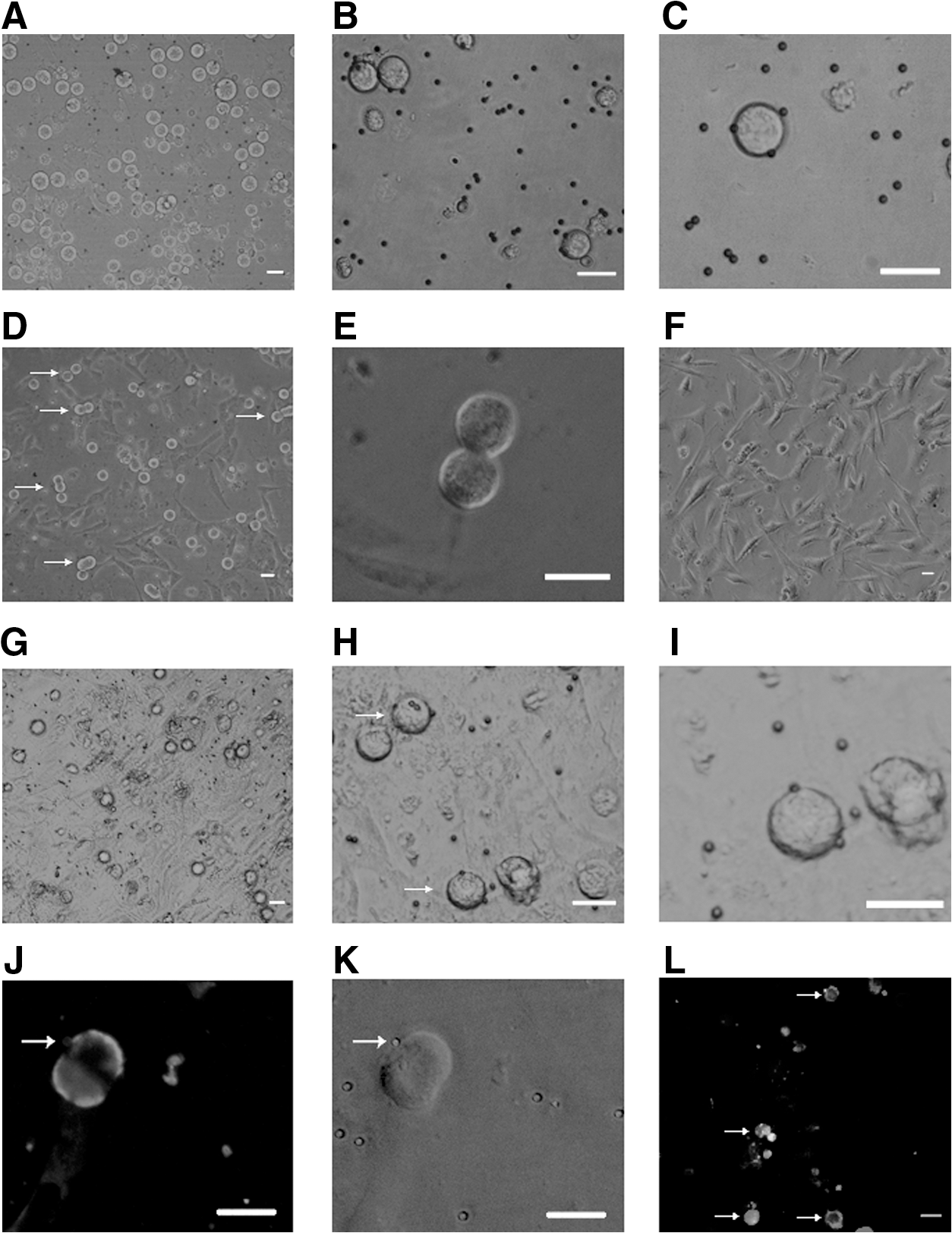
Germ cells purified by Fragilis and MVH sorting.
Characterization of purified cells
To confirm that the cells sorted by Fragilis were germ cells, we determined the expression of MVH by immunofluorescence assays at 12 h after purification. The result showed that all cells sorted were positive for MVH (Fig. 5A, B), whereas the negative fraction was mainly composed of fibroblasts (Fig. 5C) and did not express MVH (Fig. 5D). Under the reported culture conditions, a cell population with a diameter of 12–20 μm started to proliferate within 24 h (Fig. 4D, E) and expressed Fragilis (Fig. 4J, K). To further confirm that the cells collected by Fragilis sorting included dividing FGSCs, we identified the cells by BrdU/MVH dual immunofluorescence and checked the expression of Oct4, Dazl, Nanog, Figla, and Gapdh by reverse transcription–PCR, after culture for 15 days. BrdU+/MVH+ (Fig. 4L), Oct4+ /Dazl+ , and Nanog− /Figla− cells sorted by Fragilis were confirmed as FGSCs (Fig. 5E). Further, we determined the methylation status of maternally imprinted genes Igf2r and Peg 10 of Fragilis-purified cells after cultured in vitro for 3 weeks and found that they manifested a partially methylated pattern (Fig. 5F).
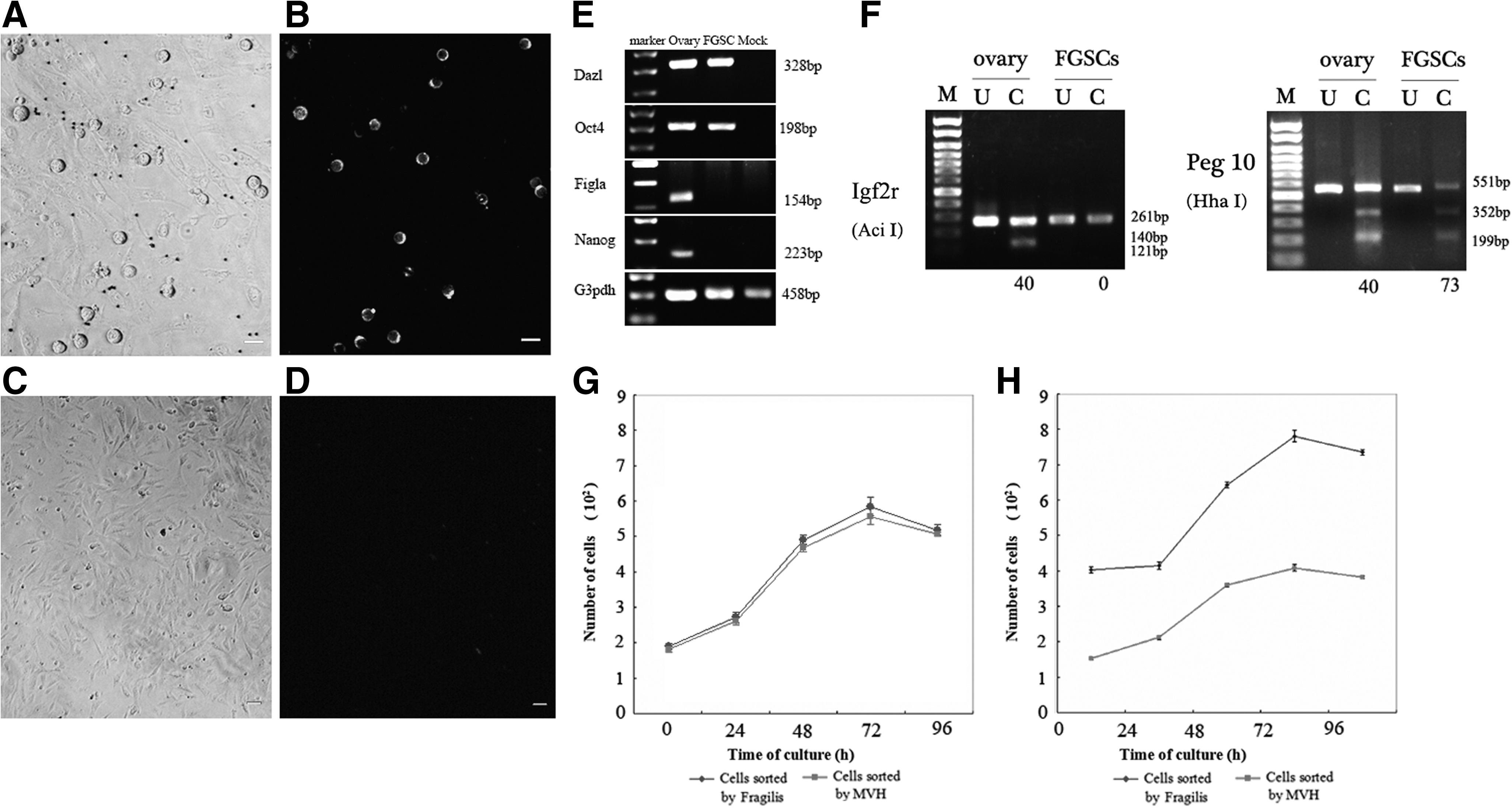
Characterization of Fragilis-sorted cells.
Then, FGSCs purified by Fragilis or MVH were passaged in 96-well plates with equivalent cell densities to produce growth curves. Cells sorted using the 2 different antigens shared identical characteristics. Based on the growth curves, we speculated that the cell cycles were 4–6 days and that the length of each phase in the 2 groups was similar (Fig. 5G). This suggests that cells sorted by Fragilis share identical growth characteristics with cells sorted by MVH. In addition, we compared the growth rates of the cells sorted by Fragilis or MVH from 10 mice. Fragilis-sorted cells showed a steeper log phase than those sorted by MVH, resulting from a higher initial seeding concentration (Fig. 5H). This suggests that Fragilis is a more efficient protein than MVH for the purification and enrichment of FGSCs and so facilitates the establishment of FGSC lines.
Discussion
We previously purified FGSCs by magnetic bead-based sorting using MVH; however, the purification efficiency was relatively low, suggesting that other germline-expressed proteins might provide more effective markers for FGSC sorting. Suitable markers for cell sorting should contain transmembrane domains and should be specifically expressed in the target cells. CD9, Stpb-c, and Fragilis have all been reported as germline markers [10,13,16] and were thus selected as potential candidates for FGSC sorting.
However, although CD9 was detected in mouse ovaries at different stages, this protein was expressed not only in the membranes of germ cells, but also in several types of somatic cells, including the granulosa cells surrounding the oocytes. This was consistent with a report of Takao et al. in 1999 [18]. FGSC purification using CD9 was therefore ruled out on the basis of the lack of germline specificity.
Our previous study reported that Stpb-c was expressed in germ cells and Sertoli cells in rodent testes [19]. The current immunofluorescence assays demonstrated that Stpb-c in mouse ovaries was only expressed in the germ cells, and not in granulosa cells or stromal cells, thus identifying Stpb-c as a germline marker in female mice. As a member of the cadherin family, Stpb-c has 1 transmembrane domain, and the N-terminus lies on the extracellular side of the membrane. However, the sequence of Stpb-c that lies outside the membrane is identical to that of its family member, long-type pituitary gland and brain-cadherin [12]. We therefore chose a 20-amino acid intracellular sequence (GHRGTSNKEDHQCPAS) located close to the membrane for antibody synthesis, to avoid binding to the long-type pituitary gland and brain-cadherin. Because the recognition domain of the Stpb-c antibody was buried on the cytoplasmic side of the membrane, the Stpb-c primary antibody failed to bind to FGSCs, and no targeted cells were collected by the magnetic beads. Thus, the female germline-specific antigen Stpb-c was not suitable for magnetic bead sorting.
Stpb-c has a similar expression pattern to MVH, but purification of FGSCs using Stpb-c failed, because the epitope was hidden in the cytoplasm. The bioinformatics tool TMPRED identified the presence of 2 putative transmembrane domains in MVH, from residues 327–350 and from residues 494–514, with both the N-terminus and C-terminus located outside the membrane. Moreover, the MVH primary antibody from Abcam (ab13840) is based on residues 700 to the C-terminus of MVH and can thus bind to the putative C-terminus domain outside the membrane. This was supported by the results of the present study; 538±42 germ cells were collected from 20 ovaries by MVH-based sorting, but no targeted cells were obtained using Stpb-c. These results indicate that proteins with putative transmembrane domains and available corresponding primary antibodies can be used for targeted cell sorting by magnetic beads, even if the bulk of the protein is located within the cytoplasm.
Fragilis was initially reported as a marker of PGCs, although fragilis knockout mice were shown to have normal germline systems [20]. The function of this germline-specific gene therefore needs further study, although it is expressed from the early germline determination stage [16]. As a cell surface protein expressed in germline cells, Fragilis has 2 transmembrane domains, and both the N-terminus and the C-terminus are located outside the membrane. The primary antibody (Abcam) recognizes residues 1–50 of the Fragilis antigen, which was confirmed by bioinformatic analysis (
Sorting of FGSCs using MVH was less efficient than that using Fragilis. MVH is an RNA-binding protein that is mainly expressed in the cytoplasm, and it is possible that only some MVH molecules bind with the membrane at any given time; the purification efficiency of MVH would thus be limited by the finite binding sites. Fragilis, in contrast, is a cell surface protein, which gives it an advantage over MVH in terms of FGSC sorting; increased number of binding sites for antibodies resulted in an almost 2-fold increase in purification efficiency. Based on our experiments, Fragilis-sorted FGSCs shared identical molecular characteristics at mRNA level and imprinted pattern at epigenetic level with FGSCs sorted by MVH.
The density of FGSCs is a pivotal factor in FGSC maintenance; however, the initial density of FGSCs is directly related to the efficiency of their purification. Comparison of the growth curves between Fragilis- and MVH-sorted FGSCs showed the former had a steeper growth velocity because of the higher initial cell density. The cell number in the 24- or 96-well plate at the start phase directly influences the enrichment and growth of FGSCs, and Fragilis sorting had the advantage of a higher efficiency of FGSC purification. This new method will thus help to reduce human influences on the process of FGSC purification, facilitate the establishment of FGSC lines, and promote further research on FGSCs.
The following important aspects have to be considered in FGSC sorting by magnetic beads: (a) incomplete 2-step digestion will produce viscous collagens that can attach to the primary antibody, resulting in poor sorting; (b) the proportions of cells after 2-step enzyme digestion, to primary antibody and to magnetic beads, should be optimized to improve sorting; (c) a slight vortex aids binding during culture with primary antibody or magnetic beads; (d) an additional rinse with PBS in the final step can increase the yield of pure germ cells; (e) omission of the citrate elution step (pH 2–3) can reduce cell death during the sorting process, because the acidic conditions are harmful to the cells.
In summary, we analyzed the utilities of several germ cell proteins for the sorting and purification of FGSCs. The efficiencies of these processes were greatly improved by the use of the cell surface protein Fragilis. This new sorting technique will facilitate the maintenance of FGSCs and the establishment of FGSC lines and will also provide the basis for further studies and applications of FGSCs.
Footnotes
Acknowledgments
This work was supported by the Major Research Plan of the National Natural Science Foundation of China (90919020 to J.W.), Program of Shanghai Subject Chief Scientist (10XD1402200 to J.W.), National Basic Research Program of China (2010CB945001, 2011CB965104), Ministry of Agriculture of the People's Republic of China (2009ZX08006-010B), and the Specialized Research Fund for the Doctoral Program of Higher Education (SRFDP) in China (20090073110032 to J.W.).
Author Disclosure Statement
No competing financial interests exist.