
Abstract
Mesenchymal stem cells (MSCs) are capable of regenerative and immunomodulatory functions in cell-based therapies in a variety of human diseases and injuries; however, their therapeutic efficacy and potential side effects remain major obstacles in clinical applications. We report here a 3D spheroid culture approach to optimize stem cell properties and therapeutic effects of human gingiva-derived mesenchymal stem cells (GMSCs) in mitigation of experimental oral mucositis. Under growth condition of ultra-low attachment, GMSCs spontaneously aggregated into 3D spheroids and exhibited distinct early stem cell phenotype characterized by elevated expression Stro-1 and CXC chemokine receptor 4 (CXCR-4) as well as OCT-4 and Nanog, 2 important transcriptional factors relevant to stem cell properties, and decreased expression of MSC-associated markers, including CD29, CD90, and CD105. Functionally, spheroid GMSCs are capable of enhanced multipotency and augmented secretion of several chemokines and cytokines relevant to cell migration, survival, and angiogenesis. More importantly, spheroid GMSCs expressed increased levels of reactive oxygen species, hypoxia-inducible factor (HIF)-1 and -2α, and manganese superoxide dismutase, which correlated with improved resistance to oxidative stress-induced apoptosis. Using an in vivo murine model of chemotherapy-induced oral mucositis, we demonstrated that spheroid-derived GMSCs possessed better therapeutic efficacy than their adherent cells in reversing body weight loss and promoting the regeneration of disrupted epithelial lining of the mucositic tongues. These findings suggest that 3D spheroid culture allows early stemness preservation and potentially precondition GMSCs for enhanced mitigation of oral mucositis.
Introduction
Oral mucositis is one of the most debilitating side effects of common cancer therapies such as chemotherapy and radiation therapy. It affects 10%–40% of patients receiving chemotherapy for solid tumors, 60%–100% of patients undergoing myeloablative regimen for hematopoietic stem cell transplant, and up to 100% of those undergoing high dose radiation therapy for head and neck cancers [11,12]. Oral mucositis is characterized by impaired regenerative capacity of the oral and alimentary epithelium, leading to atrophy, erythema, ulceration, and, eventually, the loss of mucosal barrier functions [13,14]. Clinically, oral mucositis is manifested as mouth, throat, and abdominal pain, bloating, nausea, vomiting, and severe diarrhea, along with a loss of appetite and increased risks of infection and bleeding at mucositis sites [11 –14]. These manifestations lead to a prolonged hospital stay, an increased need for antibiotics, narcotic analgesia, and parenteral nutrition, and, consequently, a huge burden in healthcare costs [13,14]. More importantly, mucositis threatens the efficacy of cancer treatment due to interruption of radio- or chemotherapy cycle and dose de-escalation, thus adversely affecting patient overall survival and quality of life [13,14]. However, most of current interventions for oral mucositis are only palliative, neither specific nor efficient at preventing or treating this complication. Even though recombinant human keratinocyte growth factor (Palifermin) is the only drug approved by FDA for the treatment of oral mucositis, its application has been greatly limited by the requirement of frequent injections, the high cost, the inconvenience, and the potential tumor-promoting effect [15 –17]. Therefore, more effective approaches for the treatment of oral mucositis are urgently needed.
Recently, we and several other groups have isolated a new population of mesenchymal stromal cells from human gingiva, namely, GMSCs, which exhibit similar stem cell-like properties and immunomodulatory abilities to human bone marrow MSCs [18 –21]. Using GMSCs as cell-based therapy, we demonstrated a reversal of colonic inflammation and restoration of body weight in colitis mice [18], and a rapid dermal closure in full-thickness excisional wounds [22]. In the present study, we seek to optimize stem cell properties of our regular adherent GMSCs by generating spheroid GMSCs using a low attachment culture condition and determine their therapeutic effects in ameliorating oral mucositis in chemotherapy-treated mice and the potential mechanisms.
Materials and Methods
Animals
Balb/c mice (male, 8–10 weeks old) were obtained from Jackson Laboratories (
Cell culture
The isolation of human bone marrow and GMSCs were described previously [18]. Gingival tissues were obtained as remnants of discarded tissues under the approved Institutional Review Board protocol at USC. MSCs were cultured either as adherent monolayers on regular dishes or as suspension culture in ultra-low attachment dishes at a density of 2×105/mL to allow 3D spheroid formation for up to 3 days. All cultures were maintained in complete alpha-minimum essential medium containing 10% fetal bovine serum (FBS), 100 U/mL penicillin/100 μg/mL streptomycin, 2 mM
Cell surface phenotype detection
Spheroid-derived MSCs and their adherent counterparts were resuspended at 2×106/mL in 100 μL phosphate-buffered saline (PBS) containing 2% FBS and incubated for 30 min at 4°C with specific primary antibodies for human CD45, CD29, CD73, CD90, CD105, CD146, CXC chemokine receptor 4 (CXCR-4; eBiosciences), Stro-1 and SSEA-4 (R&D System), or isotype-matched control IgGs, followed by fluorescein-conjugated secondary antibody. Cells were subjected to flow cytometric analysis using a flow cytometer [18].
Cell viability assays
Spheroid-derived cells and their adherent counterparts were incubated at 37°C overnight in the presence or absence of 200 μM H2O2. Cells were collected and cell viability was determined by flow cytometry using Annexin V-FITC/propidium iodide (PI) apoptosis detection kit (BD Biosciences). Viable cells are FITC Annexin V and PI negative, and early apoptotic cells are FITC Annexin V positive and PI negative, whereas cells at end stage apoptosis and death are FITC Annexin V and PI positive.
Cell cycle analysis
Cells collected and fixed in 75% ethanol, washed, and resuspended in PBS containing 7 U/mL RNase A (Sigma). About 50 μg/mL PI (Sigma) was added and incubated at room temperature for 1 h. DNA content was measured with a flow cytometer.
Adipogenetic differentiation analysis
To induce adipogenetic differentiation, MSCs at 80%–90% confluence were cultured in adipocyte induction medium (complete culture medium containing 500 nM dexamethasone, 500 nM isobutylmethylxanthine, and 50 μM indomethacin) for 2–3 weeks with fresh medium changes every 3–4 days. The adipocytes were determined by Oil Red O staining [18].
Cytokine antibody array
Cytokine expression profiles in the supernatants were detected using RayBio Human Cytokine Antibody Array 3 (RayBiotech, Inc.;
Measurement of intracellular reactive oxygen species
Intracellular reactive oxygen species (ROS) was assessed using 2′,7′-dichlorofluorescein diacetate (DCFH-DA). In brief, spheroids or their adherent counterpart GMSCs were seeded in 8-well slide chamber and incubated with 10 μM DCFH-DA for 30 min at 37°C and washed with PBS. ROS generated in the cells caused oxidation of DCFH, yielding a fluorescent product (DCF), which can be detected using a fluorescence microscope.
Immunofluorescence studies
Cells were fixed with 4% paraformaldehyde; immunostained with specific primary antibodies for Oct-4, Nanog, super-oxidative dismutase (SOD) 2, hypoxia-inducible factor (HIF)-1α, HIF-2α, CXCR-4; and then incubated with FITC- and/or rhodamine-conjugated secondary antibodies (BD Biosciences). Nuclei were counterstained with 4′,6-diamidino-2-phenylindole (DAPI). Isotype-matched control antibodies (eBiosciences) were used as negative controls.
Western blot analysis
Cell lysates were separated on polyacrylamide–sodium dodecyl sulfate gel and electroblotted onto nitrocellulose membrane (BioRad;
Oral mucositis model
Oral mucositis was induced in Balb/c mice by intraperitoneal (i.p.) injection with 5-fluorouracil (5-FU; 50 mg/kg) for 3 consecutive days [23]. On day 4, mice of the treatment groups (n=5) were intravenously infused with either spheroid-derived GMSCs (1×106/mice) or their adherent counterparts (1×106/mice). Placebo group was treated with PBS. On day 7, tongue samples were collected for further analysis. Epithelial thickness and disruption was observed using standard hematoxylin and eosin (H&E) staining and optical coherence tomography scanning. To track cells homed to the injury sites, MSCs were prelabeled with CM-DiI (max absorbance at 570 nm) before systemic injection into mice. Tissue sections were prepared and observed under a fluorescence microscope. For the BrdU labeling, the mice were injected with BrdU (50 mg/kg) 1 h before sacrifice for tissue harvesting.
Histological and immunohistochemical studies
Paraffin-embedded sections of mice tongue samples were prepared for H&E or immunohistochemical staining using primary antibodies specific for mice proliferating cell nuclear antigen (PCNA), CK-14, BrdU, human vimentin, and E-cadherin as described previously [18]. Isotype-matched control antibodies (eBiosciences) were used as negative controls. For semi-quantification, positive signals in at least 5 random high-power fields (HPF) were observed, counted, and expressed as percentage of total DAPI-positive cells (mean±SD).
Statistical analysis
All data are expressed as mean±SEM from at least 3 independent experiments. Differences between experimental and control groups were analyzed by 2-tailed unpaired Student's t-test using SPSS. P values less than 0.05 were considered statistically significant.
Results
Formation of 3D spheroid GMSCs in suspension culture condition
Recently, several studies have reported that aggregation of MSCs into 3D spheroids enhances MSC biological properties and therapeutic potentials [7
–10]. Herein, we showed that under condition of ultra-low attachment GMSCs spontaneously aggregated into small colonies as a suspension of 3D spheroids of 20–100 μm diameter (Fig. 1A). Similar to previous studies [7], GMSCs dissociated from spheroids were relatively small, nearly half in size, and more homogeneous as compared with cells derived from adherent monolayer culture as determined by microscopy (Fig. 1A) and flow cytometric analysis (Supplementary Fig. S1; Supplementary Data are available online at
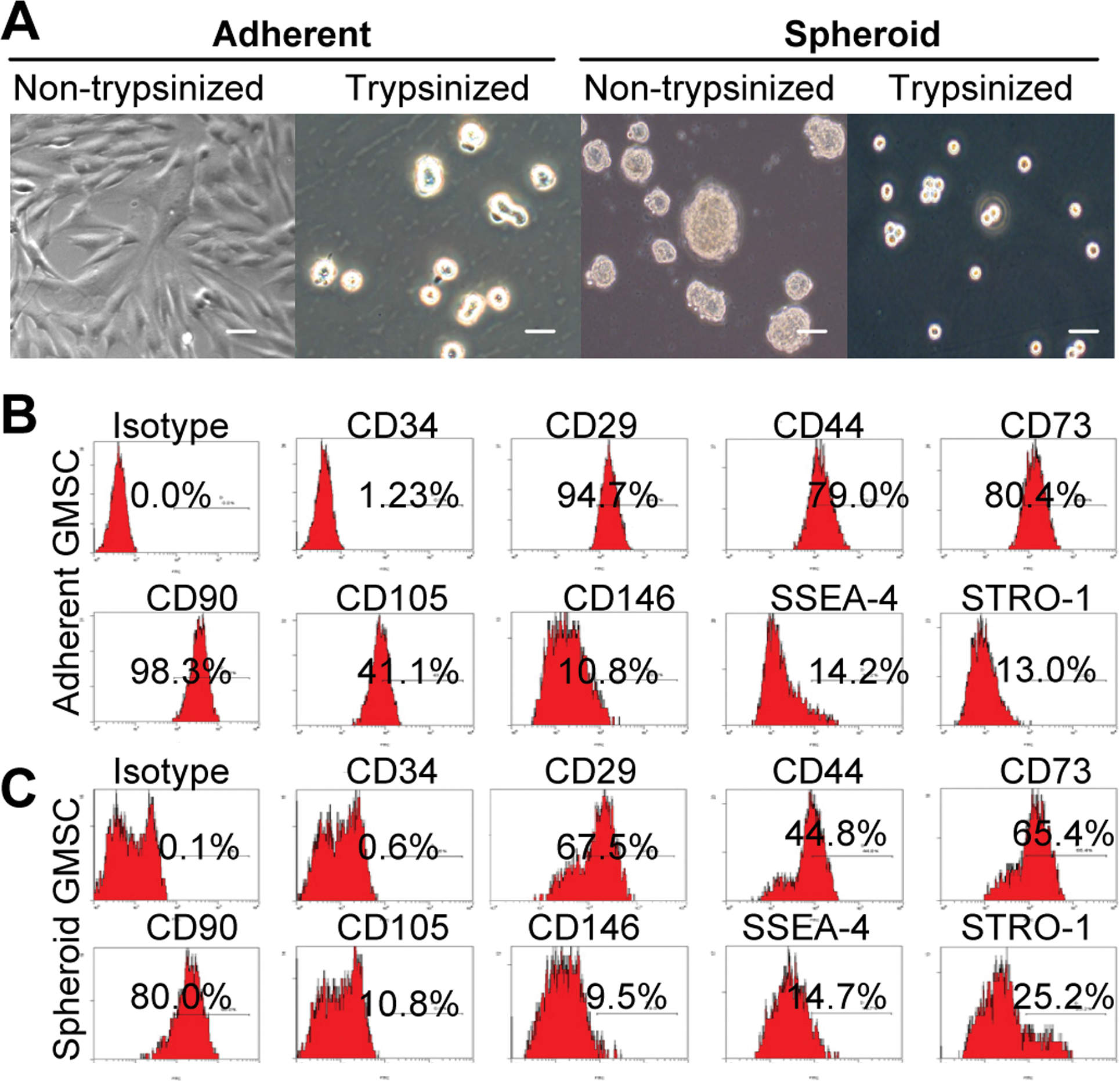
Characterization of cell morphology and surface phenotype of GMSCs grown in 3D spheroid cultures.
Enhanced expression of stem cell-related genes by spheroid GMSCs
We then examined expression of Oct-4 and Nanog, 2 important transcriptional factors in the regulation of stemness and self-renewal of a various types of stem cells [24,25]. Immunofluorescence studies indicated that expression of both Oct-4 and Nanog was significantly increased in spheroid GMSCs in comparison to their adherent counterparts (Fig. 2A). Expression of both transcriptional factors was not uniform among cells in the spheroid colonies. Increased expression of Oct-4 and Nanog in spheroid GMSCs was further confirmed by western blot analysis (Fig. 2B). These results further support the notion that 3D spheroid culture can promote the stemness of GMSCs by upregulating the expression of stem cell-related transcriptional factors.
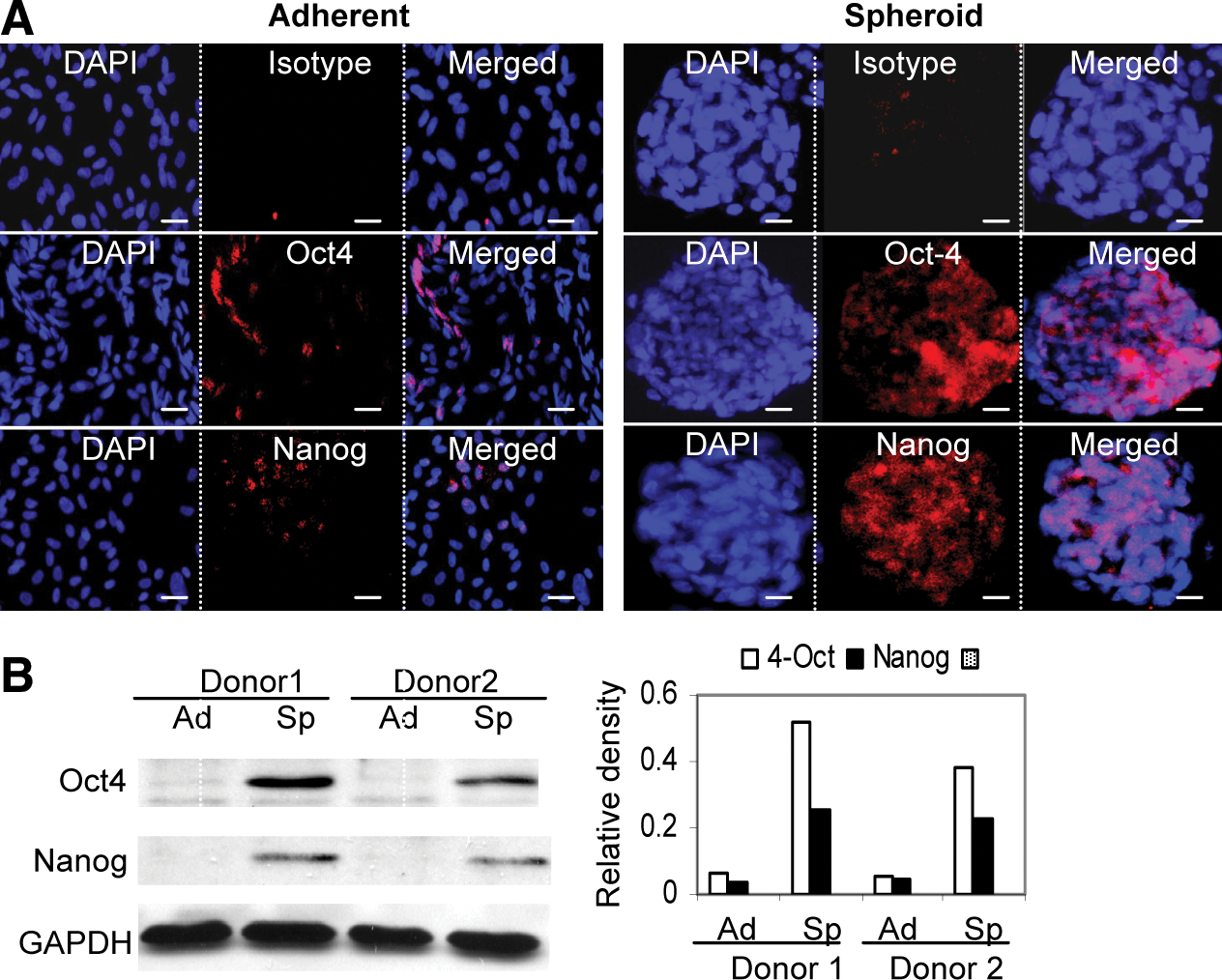
Spheroid cultures promote Oct-4 and Nanog expression in GMSCs.
Spheroid GMSCs are capable of enhanced stem-like functions
First, we studied cell cycle distribution of spheroid GMSC culture using flow cytometric analysis. As shown in Fig. 3A, spheroid culture significantly decreased the percentage of GMSCs in S-phase, with >90% of cells arrested at G0/1-phase as compared with their adherent counterparts. We next tested the multipotent differentiation of using in vitro studies. Our results showed that spheroid GMSCs displayed increased capacities to differentiate into adipocytes, suggesting an enhanced multipotency in comparison with their adherent counterparts (Fig. 3B). Meanwhile, we examined expression of CXCR-4, a receptor involved in the homing or recruitment of MSCs [6,7]. Fluorescence microscopy and flow cytometry analyses demonstrated a marked increase in expression of CXCR-4 in spheroid GMSCs as compared with cells from the adherent monolayers (Fig. 3C). MSCs exert their biological functions through the secretion of a plethora of functional soluble factors [3,5]. Therefore, antibody arrays were performed to compare the cytokine and chemokine profiles of spheroid and adherent GMSCs. Our results showed that, in comparison to the adherent GMSCs, spheroid GMSCs secrete an elevated level of a variety of cytokines and chemokines, including monocyte chemotactic protein (MCP)-2, MCP-3, RANTES, M-CSF, EGF, VEGF, SDF-1, and angiogenin (Fig. 3D), which play important role in promoting cell migration, proliferation, survival, and angiogenesis. Taken together, these results suggest that spheroid culture led to improved cellular functions of GMSCs, which might contribute to an enhanced stem-like functions and therapeutic efficacy for several clinical applications.
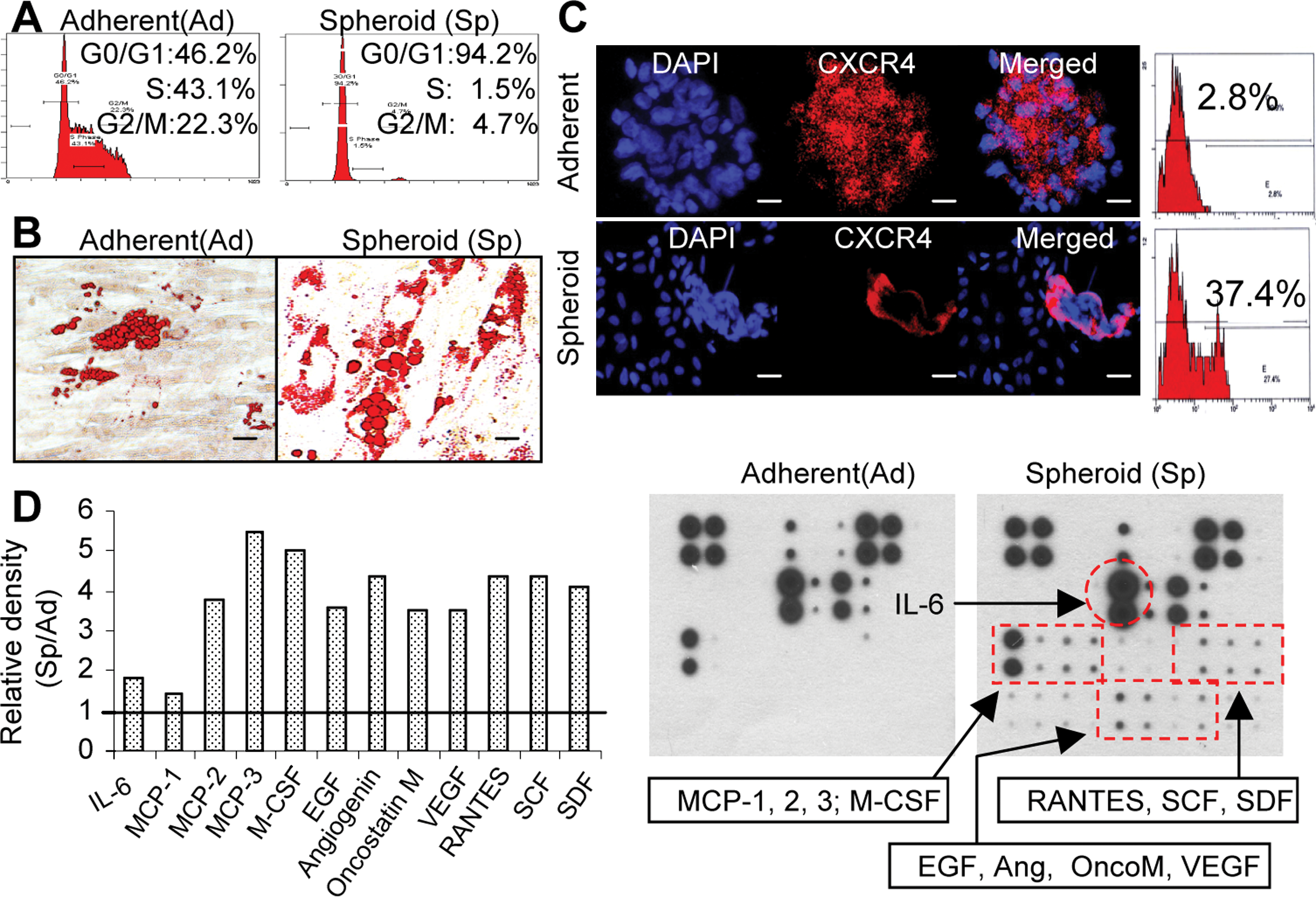
Three-dimensional spheroid culture improves cell plasticity of GMSCs.
Spheroid GMSCs are more resistant to oxidative stress
The nature of the 3D spheroid structure potentially limits cells in the inner core of adequate oxygen exchange. We postulated that a hypoxic microenvironment may be present within the spheroid structure. Our studies demonstrate an increase in the production of ROS within the GMSC spheroids as determined by DCFH-DA staining (Supplementary Fig. S2A). Meanwhile, immunofluorescence studies demonstrated that spheroid GMSCs expressed a much higher level of both HIF-1α and HIF-2α, 2 important hypoxia-responsive transcriptional factors, as compared with the adherent counterparts (Supplementary Fig. S2B, S2C). These results suggest the existence of an oxidative microenvironment within GMSC spheroids.
Manganese SOD (SOD2) is an antioxidant enzyme, which plays a key role in cellular resistance to oxidative stress via scavenging mitochondrial ROS (mtROS) [26]. We explored whether GMSCs expressed SOD2 in the presence of a potential oxidative and hypoxic microenvironment within the spheroid. Immunofluorescence studies demonstrated that expression of SOD2 was dramatically increased in spheroid GMSCs as compared with their adherent counterparts (Fig. 4A). Increased expression of SOD2 by spheroid GMSCs was further confirmed by western blot analysis (Fig. 4B). Next, we investigated the potential enhanced resistance of spheroid GMSCs to oxidative stress. To this end, both cells were exposed to exogenous H2O2 for 24 h, and apoptotic cells were tracked using annexin V-FITC. Interestingly, GMSCs prepared from 3-day spheroid culture without exposure to H2O2 contained a relatively high number of early and late apoptotic cells as compared with their adherent counterparts (P<0.05); however, after exposure to H2O2 for 24 h, the number of total apoptotic cells in spheroid derived GMSCs decreased to a lower level than that in monolayer-cultured cells (P<0.05) (Fig. 4C). Concomitantly, caspase-3 activities were suppressed in spheroid GMSCs as compared with those in adherent cells after exposure to H2O2 for 24 h (Fig. 4D). Taken together, these results suggest that spheroid culture conditioned GMSCs to hypoxic and oxidative stresses, mediated by the upregulation of hypoxia-adaptive gene expression (such as HIFs, VEGF, and SOD2), promotion of apoptosis resistance and survival ability, and enhanced secretion of chemokines and angiogenic factors.
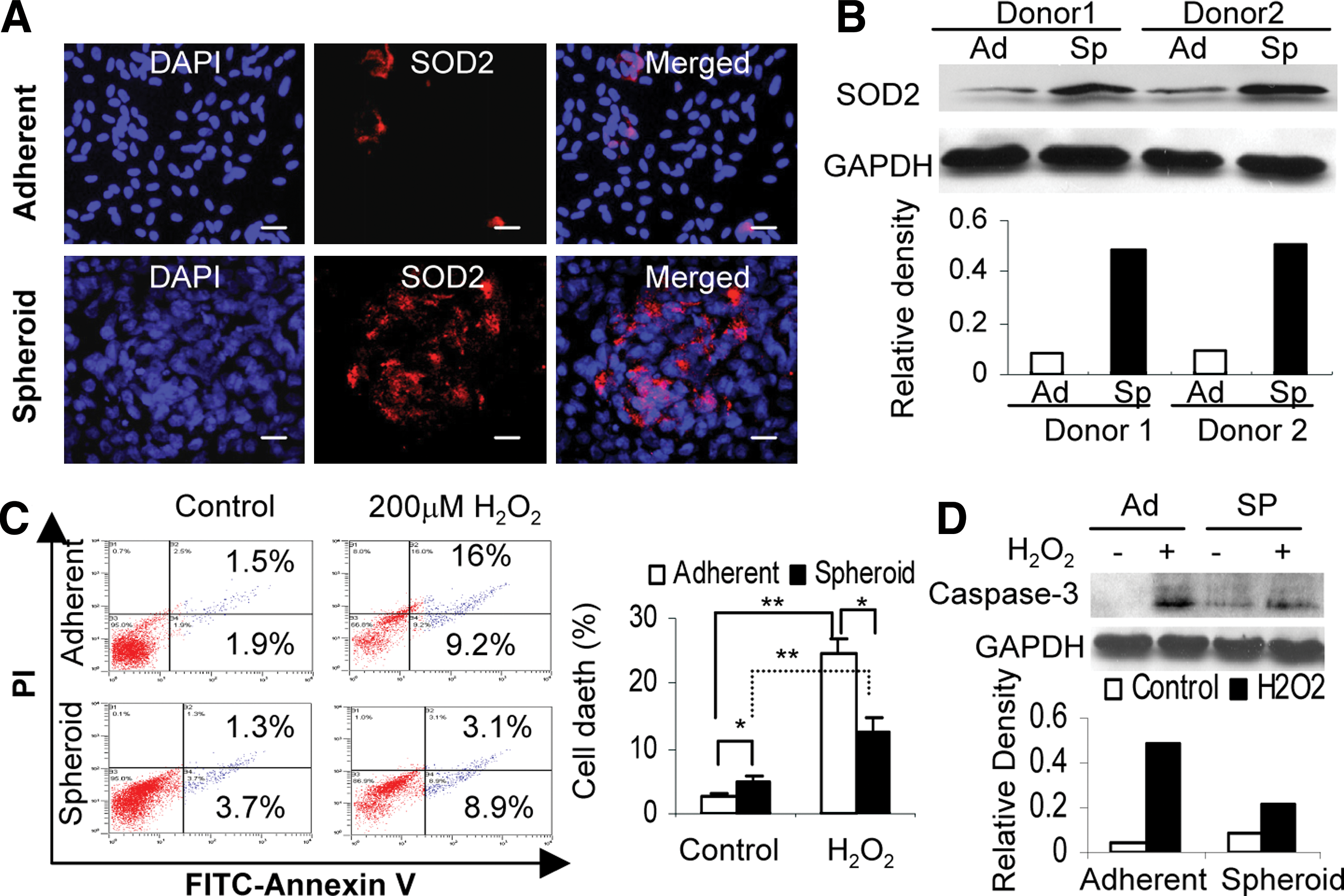
Spheroid GMSCs exhibit increased resistance to oxidative stress-induced apoptosis.
Spheroid GMSCs are capable of enhanced therapeutic effects on chemotherapy-induced oral mucositis
Given the findings that spheroid cultures were capable of optimizing multiple functions, we postulated that spheroid-derived GMSCs potentially possess improved therapeutic efficacy on wound healing or repair of injured tissues. Here, we employed a model of compromised wound healing using chemotherapy-induced oral mucositis model in mice. To this end, oral mucositis was induced by i.p. administration of 5-FU for 3 consecutive days. On day 4, the treatment groups (n=5) were intravenously injected with either spheroid GMSCs (1×106/mice) or their adherent counterpart (1×106/mice), whereas mice treated with PBS were used as disease control. Clinically, we observed a continued body weight loss in the disease group without cell treatment, which reached maximum on day 8 and started to recover on day 9 (Fig. 5A). Using optical coherence tomography coupled with multiphoton microscopy (MPM), we could delineate microscopic surface irregularities and loss of papillary structures on the dorsal surface of mucositic tongues (Fig. 6C). Interestingly, nearly all mice undergoing GMSC treatment demonstrated a restoration of body weight loss, but a much better effect was observed after treatment with spheroid GMSCs as compared with adherent cells (Fig. 5A, B). Meanwhile, MPM imaging delineated regeneration of topographical structures along with reorganization of filiform papilla in the tongue after spheroid GMSC treatment (Fig. 6C).
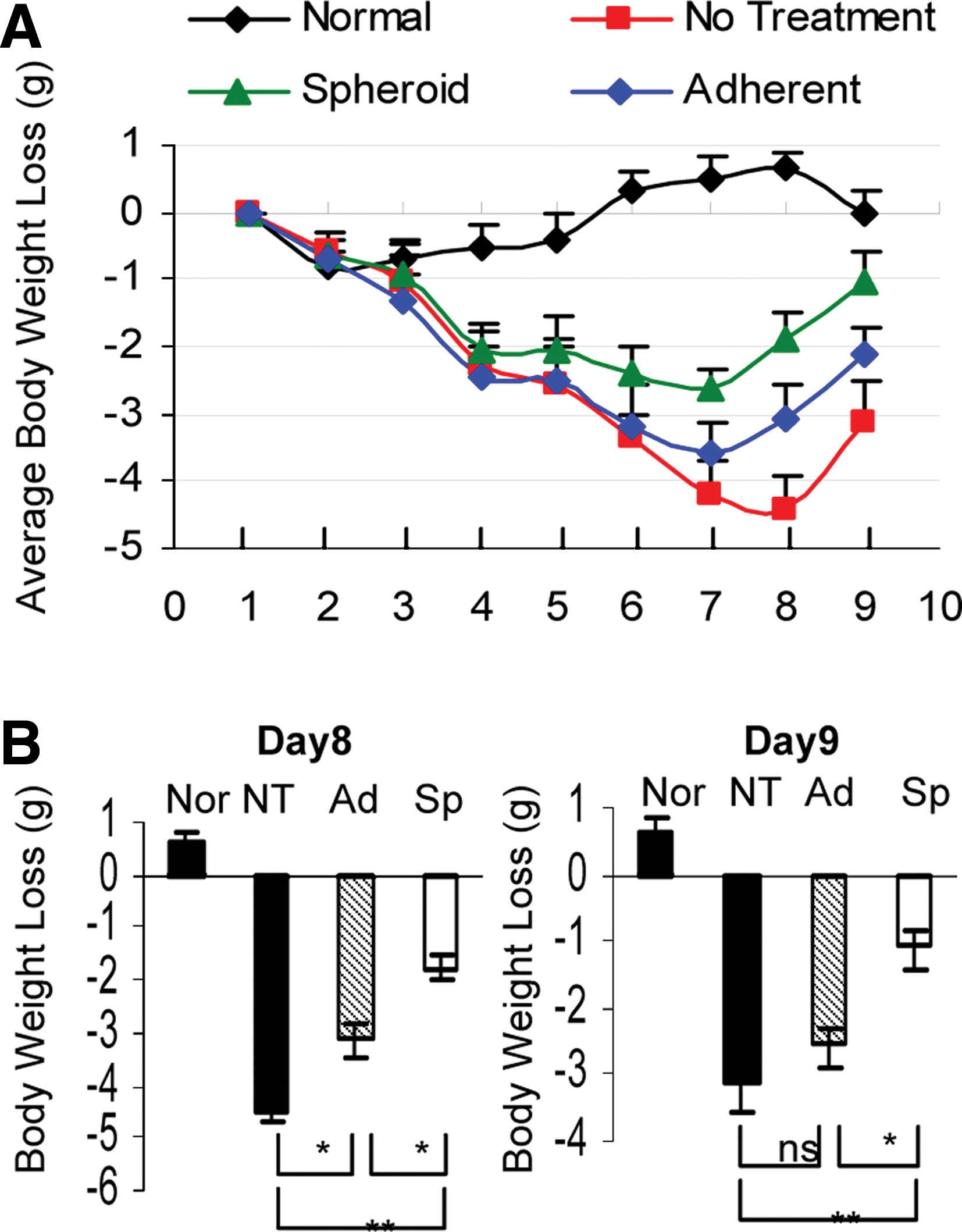
Spheroid GMSCs show enhanced therapeutic efficacy for reversing body weight loss caused by chemotherapy-induced oral mucositis. Oral mucositis was induced in Balb/c mice as described in detail in the Materials and Methods section. On day 4 after induction of oral mucositis, mice of the treatment group (n=5) were intravenously infused with either spheroid-derived GMSCs (Sp)(1×106/mice) or their adherent counterparts (Ad) (1×106/mice). Normal mice (Nor) without administration of any drug and cell treatments and mice with oral mucositis but with no cell treatment (NT) but receiving placebo treatment with phosphate-buffered saline were served as controls.
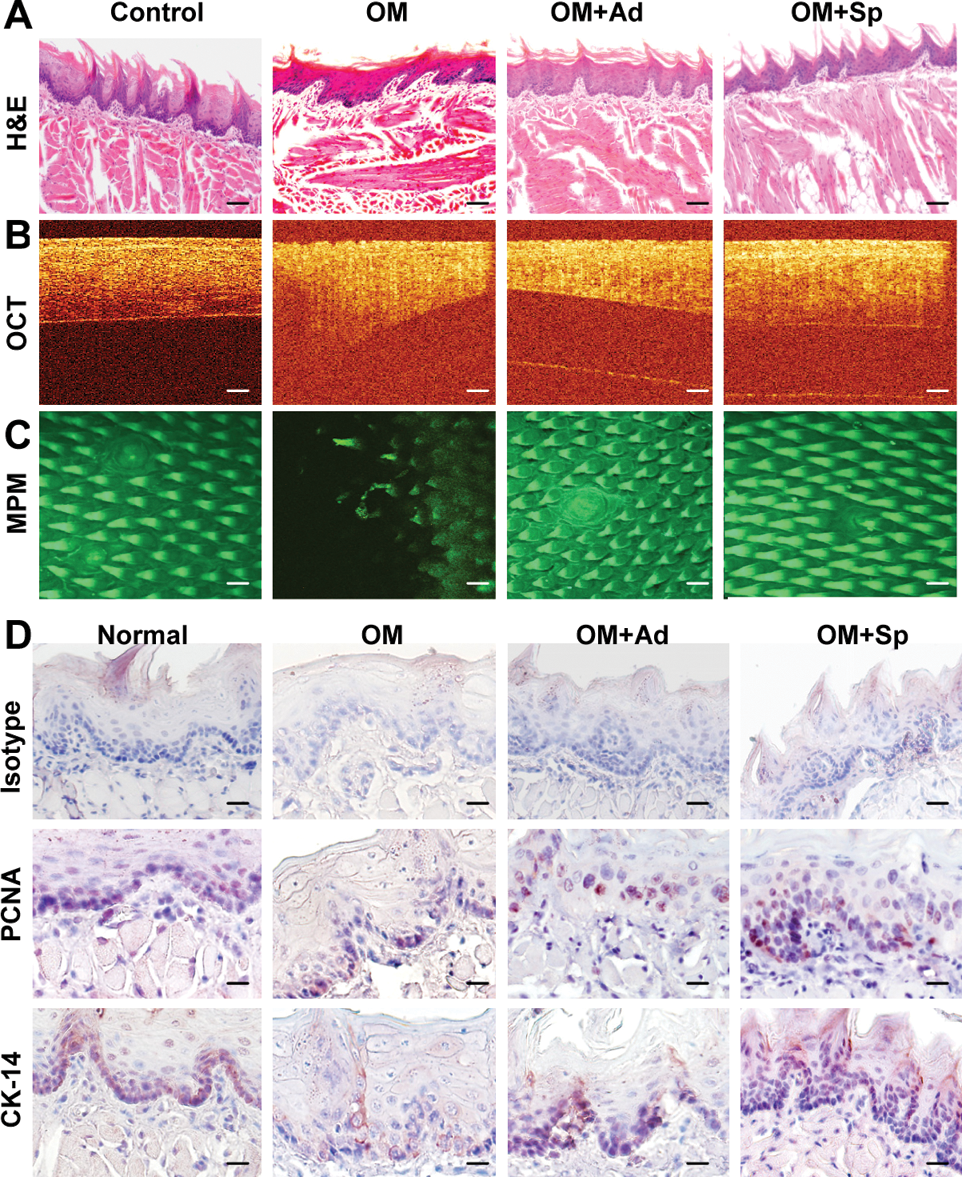
Spheroid GMSCs enhanced the recovery or regeneration of disrupted epithelial layers of mucositic tongues. On day 9, mice were sacrificed and tongue samples were harvested for further analysis.
Histologically, H&E staining and OCT imaging showed that untreated mucositic tongues displayed a complete atrophy of epithelial surface, including loss of filliform papillae and ulceration of the mucosal lining, thinning, and disruption of the epithelium layer along with the loss of the basal membrane (Fig. 6A, B). In addition, immunostaining with specific antibodies for CK14, a marker for basal layer cells, and PCNA showed a significantly decreased number of proliferating cells along with a disruptive basal cell layer in untreated mucositic tongues (Fig. 6D). Of special interest, treatment with GMSC decreased the severity and incidence of ulceration and restored the papillae structure, the lining, and thickness of the epithelial layer as compared with those of untreated disease group (Fig. 6A, B). Immunohistochemical studies demonstrated that treatment with spheroid GMSCs led to faster regeneration of the basal layer and greater restoration of the epithelial layer compared with treatment with adherent GMSCs (Fig. 6D).
Similar to previous studies, we have also shown that administration of 5-FU lead to a reduction in intestinal crypt length and obliteration of the crypts, blunting and fusion of intestinal villi, and suppressed cell proliferation in the treated jejunum as determined by in vivo BrdU labeling and immunostainings (Supplementary Fig. S3A). Interestingly, we observed that treatment with GMSCs also promoted proliferation of mucosal epithelial cells and the regeneration of damaged crypts (Supplementary Fig. S3A, S3B). All together, these results indicate that spheroid-derived GMSCs showed improved therapeutic efficacy in reversing chemotherapy-induced oral mucositis as compared with their adherent counterparts.
Spheroid GMSCs displayed increased homing to mucositis sites and underwent mesenchymal–epithelial transdifferentiation
Since spheroid-derived GMSCs expressed higher level of CXCR-4 than their adherent cultures (Fig. 3C), they might be capable of increased homing or stem cell grafting at the inflammatory mucositic sites. To confirm this, spheroid and adherent GMSCs were prelabeled with CM-DiI (570 nm) and then systemically injected into mice. We observed increased homing of spheroid GMSCs to the mucositic tongue as compared with adherent cells. More interestingly, the majority of homed adherent GMSCs were located in the subepithelial layer of tongue, whereas the homed spheroid-derived GMSCs were distributed in both epithelial and subepithelial layers (Fig. 7A), suggesting that spheroid GMSCs potentially underwent mesenchymal–epithelial transition (MET) and transdifferentiated into epithelial cells (Fig. 7A). To validate these findings, we performed immunostaining with antibodies specific for human vimentin, one of the specific markers for mesenchymal cells, and human E-cadherin, a specific marker for epithelial cells. Our results displayed abundant presence of cells positively stained for human vimentin and E-cadherin in tongue sections of mice receiving spheroid GMSC treatment as compared with those of adherent GMSC-treated mice; the majority of E-cadherin-positive cells were located in the epithelial layers (Fig. 7B). These findings support the notion that GMSCs derived from 3D spheroids are capable of increased homing or engrafting functions, specifically to the injured sites, and potentially trans-differentiate into epithelial cells, in the regeneration of the mucositic lesions of mice tongue. The increased plasticity of spheroid GMSCs may contribute to their enhanced therapeutic efficacy for regeneration or repair of injured tissues in our experimental model of oral mucositis.
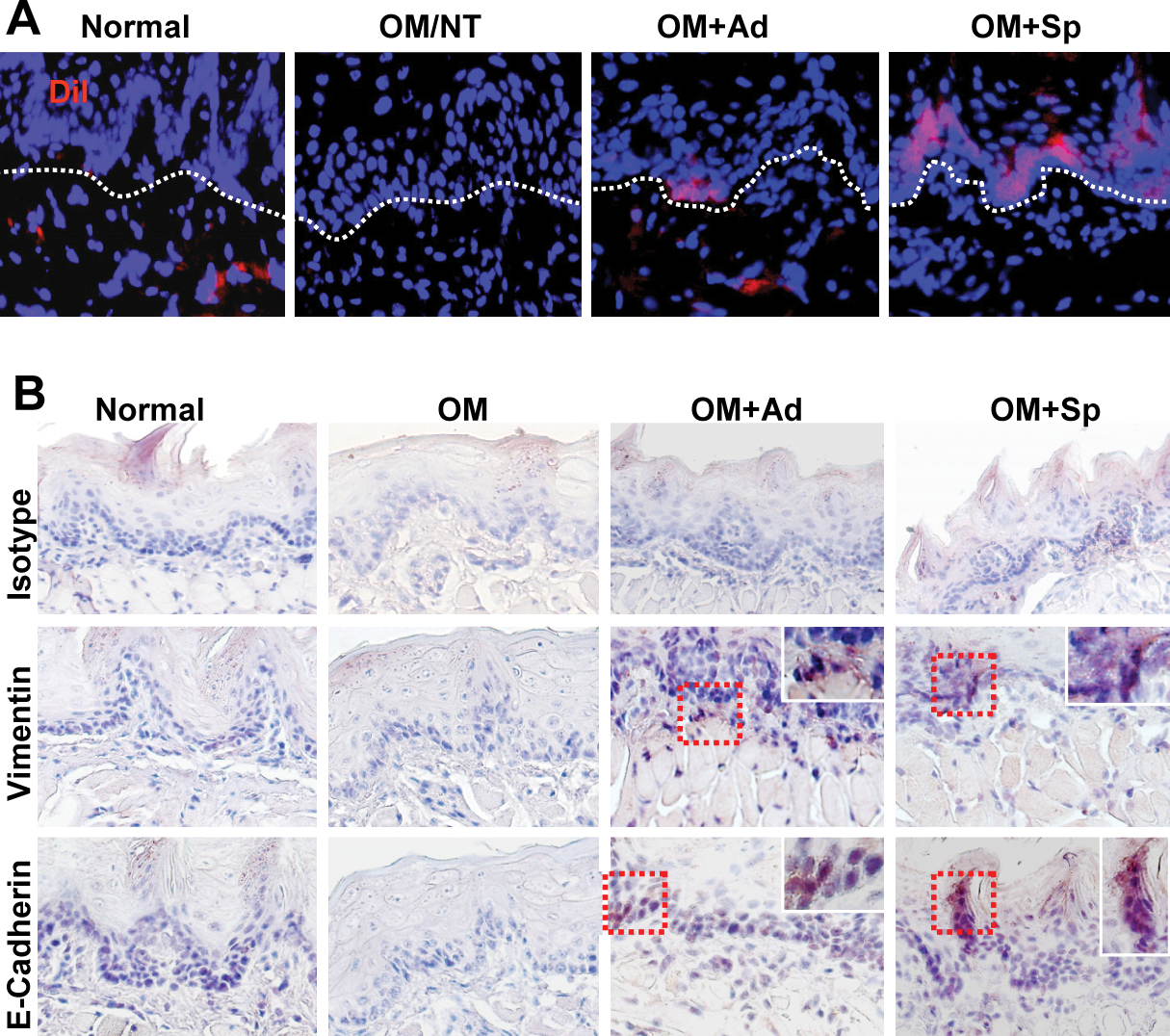
Spheroid GMSCs show increased homing ability and transdifferentiation potential to epithelial-like cells.
Discussion
Cancer therapy-induced oral mucositis represents a compromised oral wound characterized by atrophy, erythema, ulceration, and, eventually, loss of the mucosal barrier functions secondary to impaired regenerative capacity of the epithelium [11 –14]. Using a murine model of chemotherapy-induced oral mucositis, we have demonstrated for the first time to our knowledge that spheroid-derived GMSCs are capable of enhanced therapeutic efficacy to reverse body weight loss, regenerate the epithelial lining, and recover mucosal disruption as compared with their adherent cells. The improved therapeutic benefits of spheroid-derived GMSCs for oral mucositis may be attributed to their enhanced capabilities for engraftment and survival at the injury sites, trans-differentiation into epithelial cells, and preconditioning to hypoxic and oxidative challenges existing in the 3D spheroid cultures.
Despite overwhelming benefits of MSC-based therapeutics for a variety of diseases, several obstacles clearly remain for their large-scale clinical applications. It has been recognized that progressive subculturing of MSCs potentially leads to changes in cellular phenotypes affecting their regenerative and homing abilities [5,6]. Currently, intravenous delivery of a large bolus of cells and direct tissue injection are the 2 major methodologies for preclinical studies and clinical trials of MSC-based therapy. However, intravenous delivery requires a relatively large dose of cells (from 1 to 5×106 cells/kg body weight) at significant production cost and increased risk of side effects such as pulmonary embolism and organ infarction, whereas direct tissue injection requires invasive, technique-sensitive, and precise infusion methodologies [5,6]. Therefore, how to maximize therapeutic efficacy and safety of MSC-based therapy while minimizing adverse effects, overall manufacture, and procedure costs remains a great challenge. To date, transgenic approaches have been developed to force MSCs to overexpress certain defined factors with either trophic or tropic, survival, proangiogenic, immunomodulatory, or anti-inflammatory functions [5,6]. In recent years, numerous nontransgenic approaches aiming to increase the multipotency of MSCs or to evoke the secretion of these functional factors by MSCs have attracted more and more attention due to the lack of viral and mutation risks. One of the common nontransgenic approaches for optimizing MSC functions involves priming or preconditioning MSCs in vitro with certain proinflammatory cytokines such as IFN-γ or TNF-α, which may be encountered by MSCs at the sites of inflammation or injured tissues [27]. Another strategy for optimization of MSCs involves preconditioning cells under low oxygen tension or hypoxic condition, a common physiologic or metabolic milieu of stem cell niche, that potentially play a critical role in the maintenance of an undifferentiated state of both embryonic and adult stem cells, and in the regulation of proliferation and cell fate commitment [28]. For instance, previous studies have shown that bone marrow MSCs cultured in hypoxia exhibit increased expression of Oct-4 and telomerase activity [29,30] and decreased differentiation into adipogenic and osteogenic lineages [31,32]. In addition, MSCs exposed to hypoxic conditions display a more migratory or proangiogenic phenotype, and improved survival due to an increased expression of a variety of hypoxia-responsive genes involved in angiogenesis and cell migration, such as VEGF, SDF-1α, and CXCR-4 [33 –35]. Therefore, hypoxic preconditioning of MSCs is an effective methodology to optimize MSC functions, thus enhancing their global functions and therapeutic benefits after migrate to sites of inflammation and injured tissues, where they may encounter an environment of hypoxia and oxidative stresses [36,37].
Most recently, several studies have shown that 3D spheroid cultures of MSCs significantly enhance the stem cell-like properties and therapeutic effects. For example, Bartosh et al. have recently reported that aggregation of human bone marrow-derived mesenchymal stem cells (BMSC) into 3D spheroids not only enhanced their multipotent differentiation capacities, but also led to an increased expression of the TNF-α suppressing gene-6 (TSG-6), an anti-inflammatory molecule, as compared with their adherent counterparts [7]. In addition, Li et al. have shown that growing cardiac-derived progenitor cells as 3D spheroid cardiospheres led to upregulated expression of stem cell related genes and improved cell survival after exposure to oxidative stress as compared with their monolayer counterparts [8]. Most recently, Bhang et al. reported that spheroid cultures were more effective in preconditioning human adipose-derived stem cells to a hypoxic environment, leading to upregulation of hypoxia-adaptive signals and enhancing secretion of both angiogenic and antiapoptotic factors as compared with monolayer cultures [10]. Consistently, in the present study we have demonstrated that 3D spheroid culture of GMSCs not only enhanced the expression of stem cell relevant genes and their multipotent differentiation capacity but also the expression of hypoxia responsive genes such as HIF-1 and −2α, VEGF, SDF-1α, and CXCR-4, as compared with the monolayer cultures. Meanwhile, we have demonstrated that spheroid GMSCs show an increased production of ROS and superoxide dismutase-2 (SOD2) as well as improved survival under oxidative stress conditions. These findings support the notion that the 3D spheroid culture condition recapitulates hypoxic and oxidative microenvironment of the inflammatory niche at the injured or mucositic sites and potentially act as an in vitro preconditioning of GMSCs to be more resistant to apoptotic stress and optimize their therapeutic effects in reversal of oral mucositis.
Of note, the regeneration of injured or damaged epithelium-lined organs requires the coordination of multiple types of cells, the epithelial, endothelial, mesenchymal, and immune cells [38]. Recent studies have shown that mesenchymal stem cells (MSCs) participate in the regeneration and repair of a variety of diseased epithelial tissues, including injured epithelial layers in skin [39], airway [40], cornea [41], gastric and intestine [42,43], kidney [44], and oral cavity [45]. The mechanisms underlying MSC-mediated regeneration of injured epithelial tissues may involve not only the secretion of various factors with antioxidant, anti-inflammatory, antiapoptotic, or proangiogenic functions, but also the transdifferentiation of MSCs into epithelial-like cells possibly through MET [44]. In the present study, we have shown that in comparison with their adherent counterparts, spheroid-derived GMSCs displayed increased cell plasticity and abilities to home to the mucositic lesions. The relatively smaller cell sizes and increased expression of CXCR-4 by spheroid GMSCs may facilitate their faster trafficking through the lung microvasculature and more efficient distribution into tissues. Meanwhile, our findings have revealed an enhanced potential for homed spheroid-GMSCs to transdifferentiate into epithelial-like cells. This might contribute, at least in part, to their enhanced therapeutic efficacy for the regeneration or repair of mucositic lesions. However, further studies are warranted to explore the deep mechanisms.
Despite the observed therapeutic benefits of spheroid-derived GMSCs in cancer therapy-induced mucositis, their potential impact on cancer development remains to be addressed. The literature on tumor tropism, pro-, and antitumorigenicity of MSCs [46,47] is highly controversial. In some studies, MSCs have been reported to promote tumor proliferation, angiogenesis, epithelial–mesenchymal transition, as well as metastasis [46,47], whereas other studies have shown the tumor suppressive effect of MSCs in different tumor models [46]. The unique tumor tropism of MSCs renders them a unique delivery vehicle for antitumor agents [48]. Understanding the interaction between MSCs and tumor cells and their underlying mechanisms will allow development of novel therapeutic approaches to target the stromal effects on tumor growth while reversing the detrimental effect of chemotherapy-induced epithelial injuries.
Conclusion
We have demonstrated that spheroid cultures recapitulate a hypoxic microenvironment that contributes to in vitro preconditioning GMSCs to optimize their therapeutic efficacy for the treatment of oral mucositis due to their enhanced multipotency, homing, survival, and transdifferentiation capacities. These findings further support the notion that 3D spheroid cultures might provide a simple and effective strategy for optimizing MSC-based therapy for regeneration or repair of injured tissues or the treatment of other inflammation-related diseases characteristic of hypoxic and oxidative stresses.
Footnotes
Acknowledgments
This work was supported in part by National Institute of Health Research Grant, R01DE 019932, the Laser Microbeam and Medical Program (LAMMP, NIH), D41-RR01192, the California Institute for Regenerative Medicine, RN1-00572, USC Institutional funding, CTSI and Zumberge award, and Oral & Maxillofacial Surgery Foundation (OMSF).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.