
Abstract
Conventional human induced pluripotent stem cells (hiPSCs), reprogrammed from somatic cells by induced expression of Oct4, Sox2, Klf4, and c-Myc, are phenotypically different from mouse embryonic stem cells (ESCs). In mice, culture in N2B27 serum-free 2i media (mitogen-activated protein kinase/extracellular signal-regulated kinase and glycogen synthase kinase 3 inhibitors; PD0325901 and CHIR99021) plus leukemia inhibitory factor (LIF) (2i+LIF medium) enriches for germline competent ESCs. Here, we demonstrate that flat-shaped hiPSC colonies can be reprogrammed into bowl-shaped multi-potent stem cells (2i-hiPSCs) by using 2i+LIF medium. Mechanical dissociation of 2i-hiPSC colonies enables stable maintenance for >20 passages. Importantly, gene expression profiling demonstrated that 2i-hiPSCs more closely resemble primitive neural stem cells (PNSCs). Notably, this 2i-induced phenotype was generated from conventional hiPSCs, but not human ESCs (hESCs), thus correlating with the observation of neuroectodermal SOX1-positive colonies in conventional hiPSCs, but not hESCs in 2i+LIF medium. Thus, 2i-hiPSCs, which are nonteratoma forming PNSCs, may represent a safe source of cells for neural research and regenerative medicine.
Introduction
G
Oct4, Sox2, Klf4, and c-Myc (OSKM)-hiPSCs resemble blastocyst-derived hESCs in colony morphology, global gene expression and capacity for teratoma formation [5,6]. Intriguingly, hESC and hiPSC colonies exhibit characteristic flat-shaped morphology, which is clearly distinct from mESC and mouse iPSC (miPSC) colonies that have bowl-shaped morphology [7]. Further, hiPSCs and hESCs can be maintained in human ES medium with knockout serum replacement (KSR) and basic fibroblast growth factor (bFGF) but not in mESC medium with fetal bovine serum (FBS) and LIF. Thus, hESCs and hiPSCs differ markedly from mESCs and miPSCs with regard to cellular physiology. Consistent with this, several lines of evidence have suggested that hESCs are more closely related to mouse epiblast-stem cells (mEpiSCs) than they are to mESCs [8 –11]. Notably, mESC-like naïve hESCs have been generated from conventional hESCs by growth in 2i+LIF medium in the presence of Forskolin, and from conventional hiPSCs by Dox-inducible expression of OSKM in 2i+LIF medium, although establishment of stable lines from these naïve cells has been unsuccessful to date [12]. Similarly, mESC-like hESCs can be generated from conventional hESCs in Dulbecco's modified Eagle's medium (DMEM) with KSR, LIF, MEK inhibitor PD0325901 and p38 inhibitor SB203580, although the properties of these cells have yet to be fully characterized [13]. Thus, it remains unclear whether mESC-like hiPSCs can be reproducibly generated, and what characteristics mESC-like hiPSCs established in 2i+LIF medium may have.
Here, we demonstrated that bowl-shaped hiPSCs (2i-hiPSCs) can be reproducibly isolated from conventional and integration-free (Infr) hiPSCs, but not from hESCs, in 2i+LIF medium on feeder-free matrigel-coated dishes. Stable maintenance of 2i-hiPSC lines was enabled by mechanical but not enzymatic dissociation for >50 passages over a period of 9 months. High-level PAX6 and SOX2 expression and low-level OCT4 and NANOG expression demonstrated that 2i-hiPSCs had characteristics closer to multi-potent primitive neural stem cells (PNSCs) [14] also known as neural precursors [15] than miPSCs and mESCs. On withdrawal of 2i+LIF, 2i-hiPSCs efficiently differentiated into neurons and astrocytes in vitro. Further, 2i-hiPSCs were found to be incapable of forming teratomas in vivo, when implanted under the kidney capsule of immunodeficient mice. Notably, the differential ability of hiPSC and hESC lines to form PNSCs in 2i+LIF medium is due to differential induction of Sox1-positive ecotodermal cells.
Materials and Methods
Cell lines
miPSC lines were established from amniotic cells, which had been obtained from E18.5 embryos by mating of Oct4-GFP (C57BL/6) and Rosa26 (129/Sv) transgenic mice, by retroviral transduction of mouse OSKM and DsRed [16]. R1 mESC (129/Sv) and miPSC lines were maintained in mES medium [DMEM/F12 supplemented with 15% FBS, L-glutamine, penicillin-streptomycin, sodium bicarbonate, sodium pyruvate, 2-mercaptoethanol, and 1,000 U/mL of LIF (Chemicon)]. mEpiSC lines were derived from epiblasts of E6.5 embryos obtained from mating of DBA and C57BL/6 with the ubiquitously expressing GFP transgene and cultured in mEpi medium [hES medium supplemented with 8 ng/mL Activin A (Peprotech)] on mouse embryonic fibroblast (MEF) feeder cells [8,9]. Human fetal lung fibroblasts (TIG1 and TIG3) provided by the JCRB Cell Bank were used to derive hiPSC lines through retroviral induction of OSKM or OSK [16]. Infr hiPSC lines were established by OSKM transduction with episomal vectors from female adult dermal fibroblasts, HDF1388 [17]. hESC lines, KhES1, and KhES3 [18] were used according to the guidelines for derivation and utilization of hESCs from the ministry of education, culture, sports, science, and technology of Japan. hESC and hiPSC lines were stably maintained in MEF-conditioned hES medium [DMEM/F12 supplemented with 20% KSR (Invitrogen), L-glutamine, nonessential amino acids, 2-mercaptoethanol, and 10 ng/mL bFGF (Peprotech)] on matrigel-coated dishes. To address the effect of apoptosis on survival, 10 μM ROCK inhibitor Y27632 (Wako) and 10 μM Myosin IIA inhibitor Blebbistatin (Toronto Research Chemicals) were added to 2i+LIF medium. All experiments were performed with PSCs at 10–25 passages.
Cell culture in 2i+LIF medium
N2B27 medium was prepared as a 1:1 mixture of N2 medium [DMEM/F12 medium with N2 supplement (Invitrogen)] and B27 medium [Neurobasal medium with B27 supplement (Invitrogen)] supplemented with L-glutamine and penicillin-streptomycin. 2i+LIF medium was prepared by supplementing N2B27 medium with 1 μM MEK inhibitor PD0325901 (Cayman chemical), 3 μM GSK3 inhibitor CHIR99021 (Axon biochemicals), and 1,000 U/mL LIF (Chemicon).
For isolating GS-miPSC and GS-mESC lines, mES culture medium was replaced with 2i+LIF medium. For establishing GS-mEpiSC lines, 2.0×105 mEpiSCs were plated on 3.5 cm culture dishes containing feeder cells, cultured in mEpi medium overnight, and then cultured in 2i+LIF medium. Cells were split and reseeded into four 3.5 cm dishes with feeder cells after 3 days, at which time they were cultured in 2i+LIF medium. GS-mEpiSC colonies were picked 7 days after reseeding and expanded in 2i+LIF medium. Established GS-mEpiSC lines were sub-cultured with 2i+LIF medium every 3 days.
For establishing 2i-hiPSC lines, 2.0×105 conventional hiPSCs were plated on matrigel-coated 3.5 cm dishes in 2i+LIF medium. Candidate colonies were picked 5 days after 2i+LIF culture by using a micropipette. Colonies were mechanically dissociated into 40–50 cells by repeated micropipetting. This process was repeated until 2i-hiPSC lines were established. Established lines were subsequently sub-cultured by mechanical dissociation in 2i+LIF medium every 5–7 days.
Chimera and teratoma formation
For producing chimeric embryos, GS-miPSCs and GS-mEpiSCs ubiquitously expressing β-geo and EGFP, respectively, were microinjected into C57BL/6J x BDF1 blastocysts. Embryos were collected at E15.5 and analyzed by X-gal staining and GFP fluorescence [19].
For obtaining teratomas, undifferentiated 2i-hiPSC lines (about 5.0×106 cells/site) and derived EBs (1.0×107 cells/site) were transplanted under the kidney capsule of immunodeficient SCID mice (Clea) by a transfer pipette. Teratoma formation was assessed after 8–10 weeks.
Gene expression
For reverse transcription (RT)-polymerase chain reaction (PCR) analyses, total RNA of cultured cells was extracted with TRIzol reagent (Invitrogen). cDNA was synthesized from 500 ng total RNA with Superscript III (Invitrogen) by using random hexamers while following the manufacturer's instructions. For mEpiSC RNA extraction, mEpiSC colonies grown on feeder cells were purified by mechanical picking and pooled in TRIzol. Primer sequences used in this study are summarized in Supplementary Table S1; Supplementary Data are available online at
For global gene expression analyses, 250 ng total RNA was labeled according to standard Affymetrix protocols and hybridized to the Affymetrix mouse genome 430 2.0 Array (samples of mouse origin) or the human genome U133 Plus 2.0 Array (samples of human origin). Raw data were normalized by the MAS 5.0 method by using bioconductor package on R program (
Immunocytochemistry
Cultured cells were fixed with 4% paraformaldehyde (PFA)/phosphate-buffered saline (PBS) for 10 min at room temperature, washed with PBST (0.1% Triton X-100 in PBS), and then pretreated with blocking solution [3% bovine serum albumin and 2% skim milk (DIFCO) in PBST] at 4°C overnight. The cells were then stained with fluorescence-conjugated secondary antibodies (1:500; Invitrogen), following an immunoreaction with primary antibodies; anti-OCT4 (1:50; Santa Cruz Biotechnology), anti-SOX2 (1:500; Abcam), anti-Klf4 (1:200; Santa Cruz Biotechnology), anti-NANOG (1:200; ReproCELL), anti-SSEA1 (1:200; DSHB), anti-SSEA4 (1:200; DSHB), anti-E-CADHERIN (1:50; Takara), anti-TRA1-60 (1:500; Millipore), anti-Tubulin J1 (TuJ1) (1:600; COVANCE), anti-neurofilament-M (NF-M) (1:1,000; Millipore), anti-glial fibrillary acidic protein (GFAP) (1:1,000; Dako), anti-O4 (1:60; Millipore), anti-H3K27me3 (1:200; Millipore), anti-SOX1 (1:100; R&D systems), and anti-N-CADHERIN (1:100; Chemicon). The cells were counterstained with 4,6-diamidino-2-phenylindole and mounted with SlowFade light antifade kit (Invitrogen).
XIST RNA FISH
Cells were fixed with 4% PFA/PBS. RNA FISH was performed according to standard procedures with an RNA probe prepared by in vitro transcription with Cy3-UTP (GE Healthcare) and a human full-length XIST cDNA construct [20] using T7 RNA polymerase.
Cell differentiation in vitro
For neural differentiation, 2i-hiPSCs were cultured on matrigel-coated dishes in N2B27 medium for 4 days after withdrawal of 2i+LIF. Differentiated cells were fixed with 4% PFA/PBS and analyzed by immunocytochemistry.
For EB formation, small 2i-hiPSC aggregates were formed by overnight hanging drop cultures in DMEM containing 10% FBS. Aggregates were grown and differentiated in bacterial culture dishes for 5–7 days.
Results
Mouse GS pluripotent stem cells in 2i+LIF medium
To investigate the suitability of serum-free neurobasal N2B27 medium with 2i+LIF (2i+LIF medium) for generation of GS PSCs, we tested 3 types of mouse conventional PSCs: mESCs, miPSCs, and mEpiSCs. GS-mESCs, GS-miPSCs, and GS-mEpiSCs, similarly characterized by bowl-shaped morphology of colonies expressing Oct4, Sox2, and Nanog proteins, were reproducibly isolated (Fig. 1A). GS-mEpiSCs were isolated as a small number of colonies from conventional mEpiSCs as a consequence of stringent selection in 2i+LIF medium, whereas only mild cell selection was required for the transition from conventional mESCs and miPSCs to GS-mESCs and GS-miPSCs, respectively. These results were verified by triplicate experiments. Before being cultured in 2i+LIF medium, miPSCs exhibited incomplete silencing of exogenous Oct4, Klf4, and c-Myc, whereas mEpiSCs exhibited low expression of Rex1, Nr0b1, and Stella accompanied by high expression of Lefty1, Sox17, and Fgf5. After culture in 2i+LIF medium, both GS-miPSCs and GS-mEpiSCs exhibited similar gene expression patterns to GS-mESCs (Fig. 1B). This was verified by analyses of global gene expression profile with expression of microarray assays (Fig. 1C). The efficient contribution to a broad range of cell types in E15.5 chimeric embryos demonstrated pluripotency; of GS-miPSCs with the LacZ reporter gene and GS-mEpiSCs with the GFP reporter gene (Fig. 1D). Our observations on GS-mESCs and mEpiSCs are consistent with findings previously reported [1,8,9,21,22], thus clearly indicating that the 2i+LIF medium used here is suitable for generating and maintaining mouse GS PSCs.
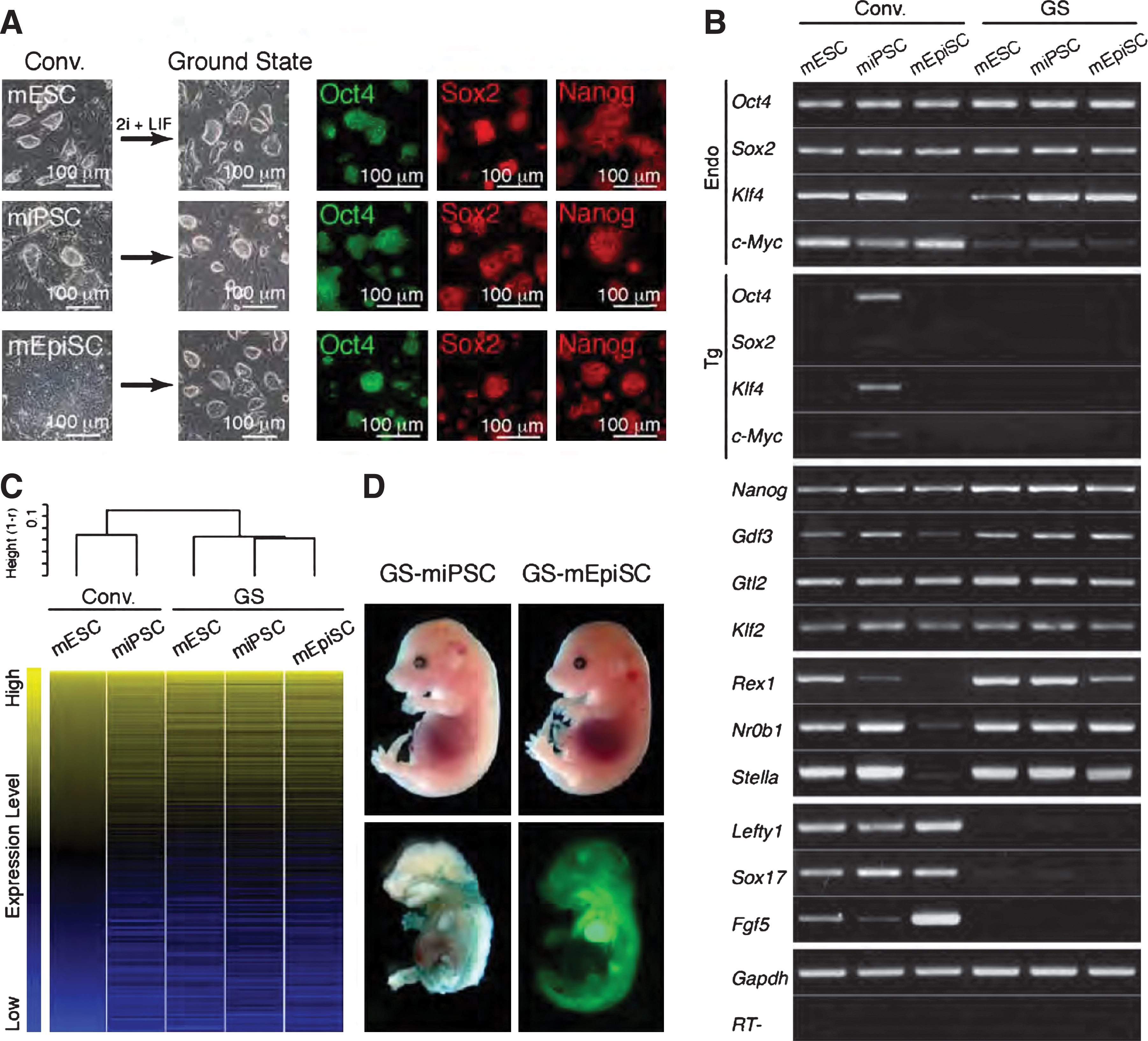
Generation of mouse ground state (GS) PSC lines.
Generation of human iPSC lines in 2i+LIF medium
To determine whether human somatic cells can be directly reprogrammed into bowl-shaped 2i-hiPSCs, the human primary fetal lung fibroblast lines, female TIG1 and male TIG3, were cultured for 20 days after retroviral transduction of OSKM in human ES medium with KSR and bFGF. Twenty-four hours after replacing the culture medium with 2i+LIF, surviving colonies were picked and expanded by gentle dissociation with a glass pipette for 3–4 passages. This resulted in numerous 2i-hiPSC colonies with bowl-shaped mESC-like morphology (Fig. 2A).
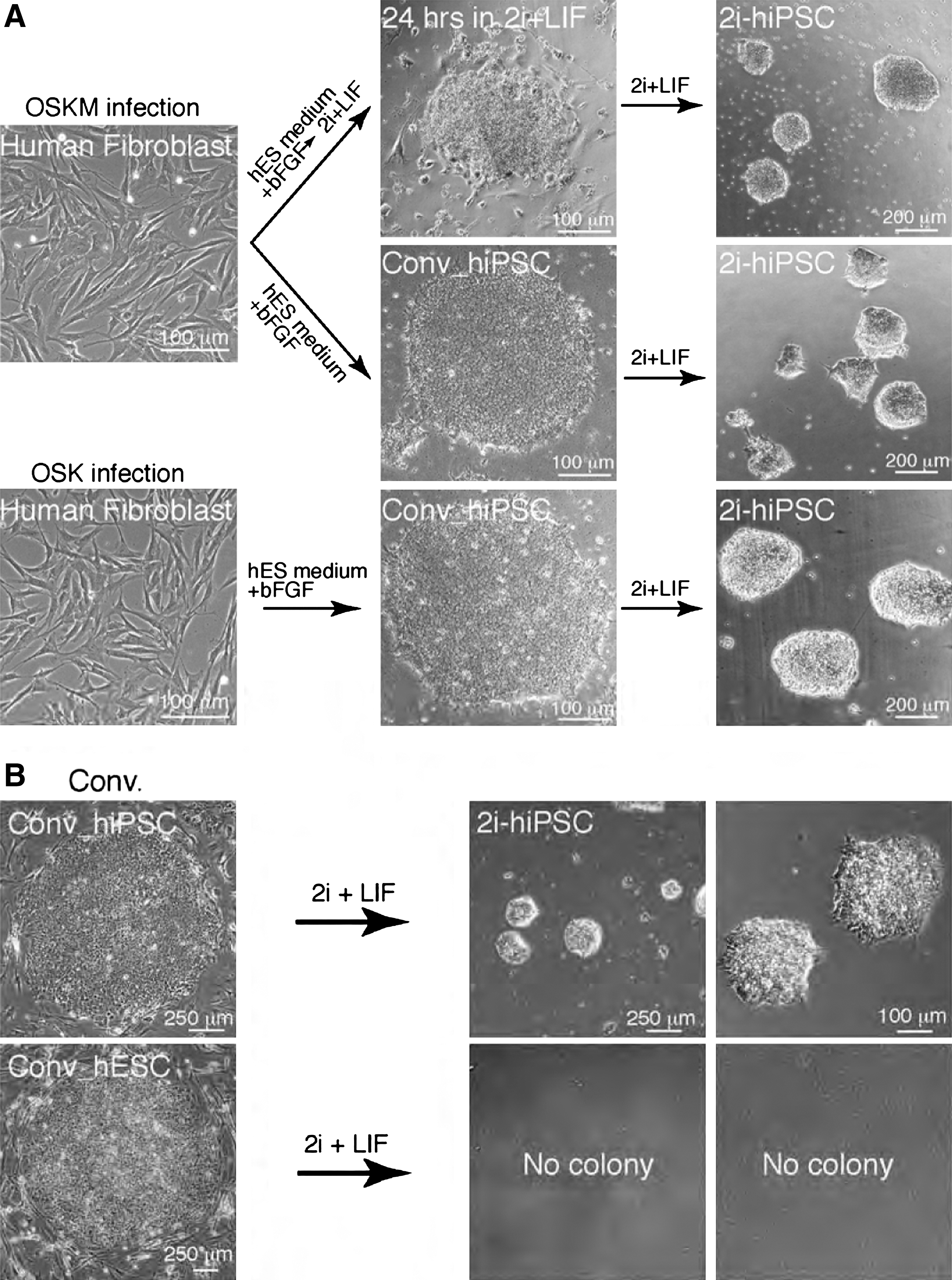
Generation of human multipotent stem cell lines in 2i+LIF medium.
To further investigate whether these 2i-hiPSCs had been directly derived from the conventional flat-shaped OSKM-hiPSCs established from the TIG1 and TIG3 cells, the culture medium of conventional hiPSCs was switched from human ES medium with KSR and bFGF to 2i+LIF medium. Conventional hiPSC colonies were severely selected for 5 days of culture. However, after the longer selection period (5–7 days), a small number of bowl-shaped colonies (5–10 colonies from about 2.0×105 conventional hiPSCs) were observed. The bowl-shaped colonies were picked and expanded in fresh 2i+LIF medium after mechanical dissociation by using a glass micropipette (Fig. 2A). Bowl-shaped 2i-hiPSC colonies were reproducibly established from OSKM-hiPSC lines derived from both independent somatic cell lines, TIG1 and TIG3. In total, more than fifty 2i-hiPSC lines were established. Of 5 2i-hiPSC lines examined, all were karyotypically normal 2n=46, XY or XX (data not shown).
To examine whether 2i-hiPSCs can be generated from iPSC lines that do not contain the integrated exogenous c-Myc gene, conventional iPSCs produced by retroviral OSK transduction were cultured in 2i+LIF medium. Notably, 2i-hiPSC lines were reproducibly isolated from conventional OSK-hiPSCs at a similar frequency to that observed from conventional OSKM-hiPSCs. The 2i-hiPSC colonies resembled those derived from OSKM-hiPSCs, with characteristic bowl-shaped colony morphology and cell growth (Fig. 2A). Thus, integration of c-Myc is dispensable for 2i-hiPSC generation from conventional hiPSCs.
To expand the number of 2i-hiPSCs, we also attempted enzymatic dissociation with trypsin and collagenase. Strikingly, none of the cells were capable of complete re-attachment to matrigel-coated culture dishes. Subculture in the presence of the small molecule Tzv [13] was found to improve attachment, but not survival. In fact, Tzv induced rapid cell differentiation toward neuronal lineages (data not shown). This suggested that subculture by enzymatic dissociation is not suitable for expansion and long-term maintenance of 2i-hiPSC lines. Consequently, 2i-hiPSC colonies were routinely dissociated to clusters of 20–50 cells by using a glass micropipette under a stereomicroscope. For stable maintenance, dissociated 2i-hiPSC clusters were sub-cultured into fresh 2i+LIF medium in matrigel-coated dishes every 5–7 days. In this manner, we were able to stably maintain and expand 2i-hiPSC clones for >50 passages over a period of 9 months under feeder-free conditions. Further, 2i-hiPSCs were successfully cryo-preserved and thawed with hESC freezing medium.
Conventional hESCs and nonintegrated hiPSCs in 2i+LIF medium
To determine whether 2i-hESCs can be generated from conventional hESCs without integration of exogenous genes, conventional hESCs were cultured in 2i+LIF medium. No 2i-hESC colonies were detected despite 3 repeated experiments with 2 different hESC lines, KhES1 and KhES3 [18], with or without ROCK/Myosin IIA inhibitors [23,24]; whereas 2i-hiPSCs were repeatedly generated in the same 2i+LIF medium (Fig. 2B). Further, no 2i-hESC colonies appeared even after addition of Forskolin to the 2i+LIF medium [12]. These findings could be explained by 2 possibilities: (1) transient expression of exogenous OSK is required for 2i-hiPSC generation, or (2) generation potential into multipotent stem cells in 2i+LIF medium is retained in conventional hiPSCs but not hESCs.
To address this, 3 independent OSKM Infr conventional iPSC lines derived from adult dermal cells HDF1388 [17] were cultured in 2i+LIF medium. Importantly, 2i-iPSC lines were repeatedly isolated from each conventional cell line, even without exogenous OSKM integration. Therefore, it is likely that generation of 2i-hiPSCs but not 2i-hESCs reflects differential potential of cell reprogramming into multipotent stem cells in 2i+LIF medium between conventional hiPSCs and hESCs.
Gene expression in 2i-hiPSCs
To explore the differential reprogramming potential of conventional hiPSCs and hESCs in 2i+LIF medium, the global gene expression profiles of conventional hESCs and hiPSCs were compared with those of 2i+hiPSCs by microarray analyses. Notably, 2i-hiPSCs exhibited significant differences from the conventional lines (Fig. 3A). 2i-hiPSCs showed reduced expression of pluripotent marker genes with the exception of SOX2. To verify the expression profile, selected genes were further investigated by RT-PCR (Fig. 3B). Pluripotent marker genes, OCT4, NANOG, NR0B1, and STELLA, were expressed at much reduced levels. However, consistent with the whole genome analysis, markedly higher levels of SOX2 expression were observed. Notably, the expression profiles and bowl-shaped colony morphology resembled those of PNSCs, which are capable of contribution into chimeras, and are defined by low level expression of the pluripotent marker genes, Oct4 and Nanog, and high level expression of the neuroectodermal marker genes, Nestin, Sox1, and Pax6 in mice [14]. Thus, expression of PNSC marker genes was analyzed. Notably, considerable high-level SOX1 and PAX6 expression was observed in 2i-hiPSC lines but not conventional hESC and hiPSC lines (Fig. 3C). Expression of NESTIN was also detected in 2i-hiPSC lines, as well as in conventional hESC and hiPSC lines. Further, an inactivated X chromosome was detected by H3K27me3 staining and RNA FISH analysis in each female 2i-hiPSC (Supplementary Fig. S1). Therefore, we conclude that 2i-hiPSCs are phenotypically equivalent to mouse PNSCs, and not authentic mESCs. 2i+LIF medium does not provide appropriate culture conditions for generating authentic hESCs, in contrast to generation of GS-mESCs.
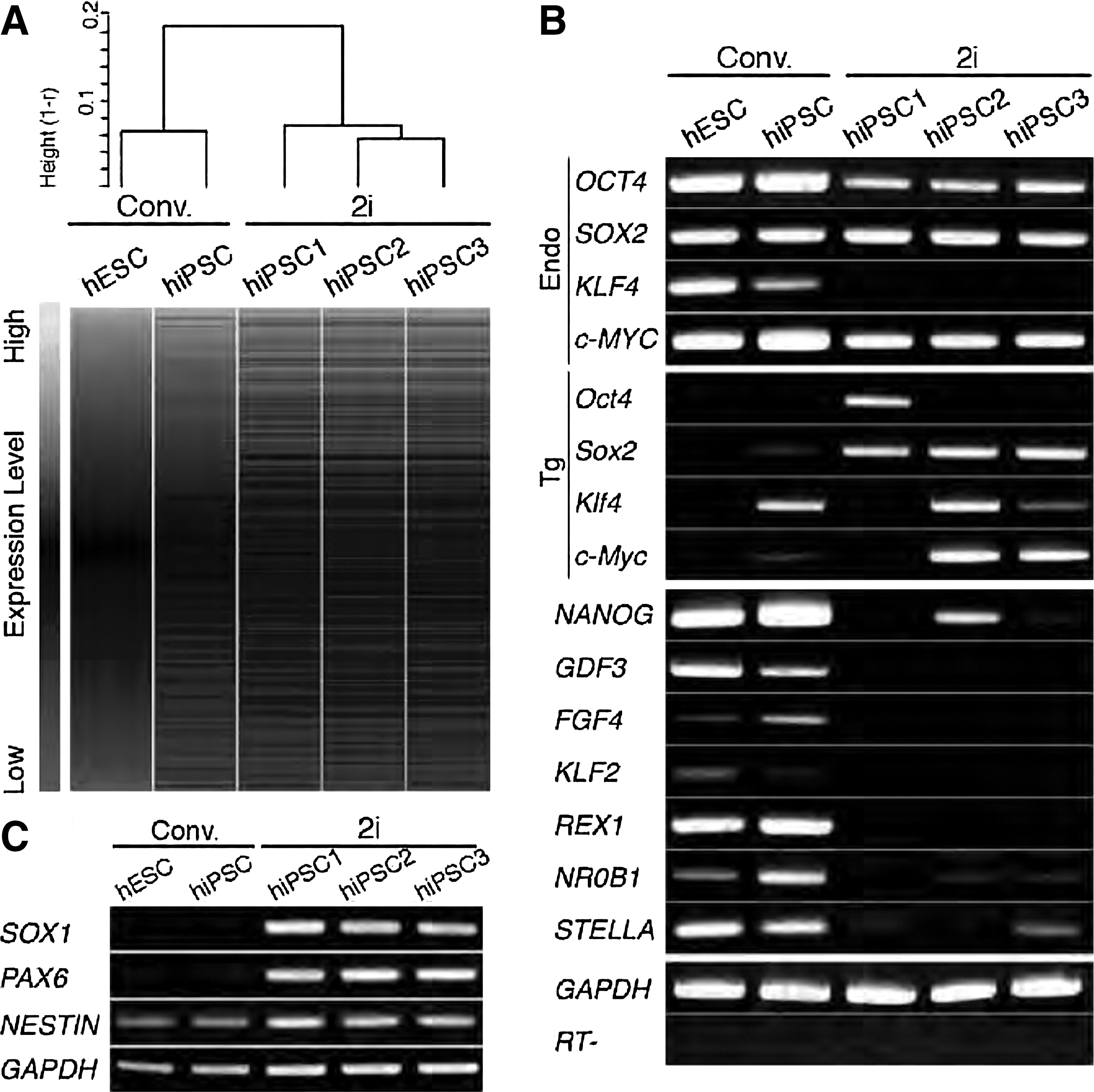
Gene expression analysis of human iPSCs in 2i+LIF medium (2i-hiPSCs).
Emergence of differential lineages from hESC and hiPSC in 2i+LIF medium
Human PNSCs were generated from conventional hiPSCs but not hESCs. Conventional hESC lines, KhES1 and KhES3, were derived from Japanese male and female blastocysts; whereas conventional hiPSC lines and conventional Infr hiPSC lines were derived from fetal lung fibroblasts TIG1 and TIG3, and adult dermal fibroblasts HDF1388, respectively. It is, therefore, likely that origin of the somatic cells is not associated with differential emergence of differentiated cells from hESC and hiPSC lines in 2i+LIF medium. To explore how the differential response occurs, we examined expression of lineage marker genes, including PNSC markers, PAX6 and SOX1, in colonies picked for 2i-hiPSC generation 5 days after culture in 2i+LIF medium (Fig. 4A). The reprogramming frequency to pick a colony was ∼30% (164 out of 536 colonies examined). Candidate colonies of 2i-hiPSCs that appeared during culture of conventional hiPSCs and Infr hiPSCs specifically expressed SOX1; whereas other cell populations expressed endodermal (GATA4), mesodermal (RUNX1 and T), and trophectodermal (CDX2) lineage markers associated with down-regulation of the pluripotent marker OCT4 (Fig. 4B). NESTIN was expressed in both picked and other colonies, whereas PAX6 was undetectable at the earliest conversion stage. Specific expression of the SOX1 protein in candidate 2i-hiPSC colonies was verified by immunostaining (Fig. 4C). These findings in human cells were consistent with properties of PNSCs in the mouse. Sox1 is the earliest specific marker of neuroectoderm in mouse embryos in vivo [25]. Neural precursor cells were FACS-purified as Sox1-positive cell populations in differentiation of mouse Sox1-GFP knock-in ES cells in vitro [15]. These data support the notion that 2i-hiPSCs are phenotypically equivalent to mouse PNSCs.
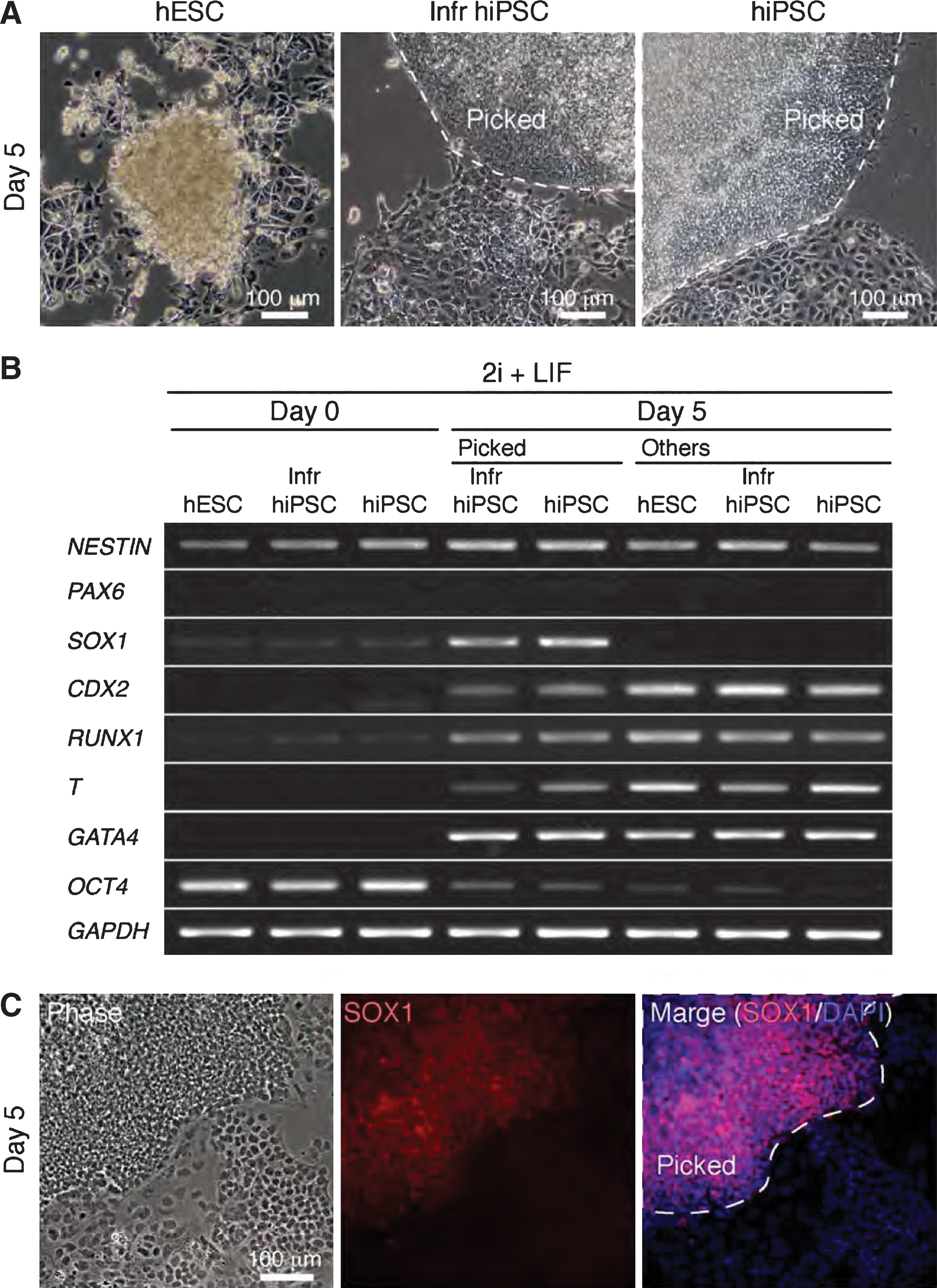
Emergence of human iPSCs in 2i+LIF medium (2i-hiPSCs).
To examine which defined supplements in the culture medium were responsible for differential emergence of SOX1-positive colonies, conventional hiPSCs and hESCs were cultured in N2B27 medium with or without 2i+LIF. Differential emergence of differentiated cells between hiPSCs and hESCs was detected only in N2B27 medium with 2i+LIF (Supplementary Fig. S2). Withdrawal of 2i+LIF induced cell differentiation into both neural and non-neural cells. Thus, we conclude that a combination of stimulatory effect of N2B27 on cell differentiation and chemical selection of 2i+LIF-resistant neural cells is required for 2i-hiPSC generation.
Characterization of multipotent 2i-hiPSCs
To further characterize 2i-hiPSCs, pluripotent marker protein expression in undifferentiated human and mESCs was examined by immunostaining. 2i-hiPSCs were positive for OCT4, SOX2, and KLF4, weakly positive for NANOG, and negative for E-CADHERIN, SSEA1, SSEA4, and TRA-1-60 (Fig. 5A). Consistent with data on gene expression analyses (Fig. 3A, B), the protein expression profile of 2i-hiPSCs was clearly different from that of conventional hESCs which are positive for E-CADHERIN, SSEA4, and TRA-1-60. Next, to analyze the similarity of 2i-hiPSCs to PNSCs, differentiation of 2i-hiPSCs was induced in vitro. Cystic embryoid bodies (EBs) were successfully generated within 5–7 day suspension culture for all 2i-hiPSC lines tested (Fig. 5B), thus indicating that 2i-hiPSCs can differentiate at least into the endodermal lineage. Ectodermal differentiation was evidenced by efficient differentiation into neural cells, induced 4–5 days after withdrawal of chemicals, 2i and LIF, from 2i+LIF medium (Fig. 5B). Similar neural differentiation was induced several days after replacement of 2i+LIF medium with DMEM-F12 medium containing FBS (data not shown).
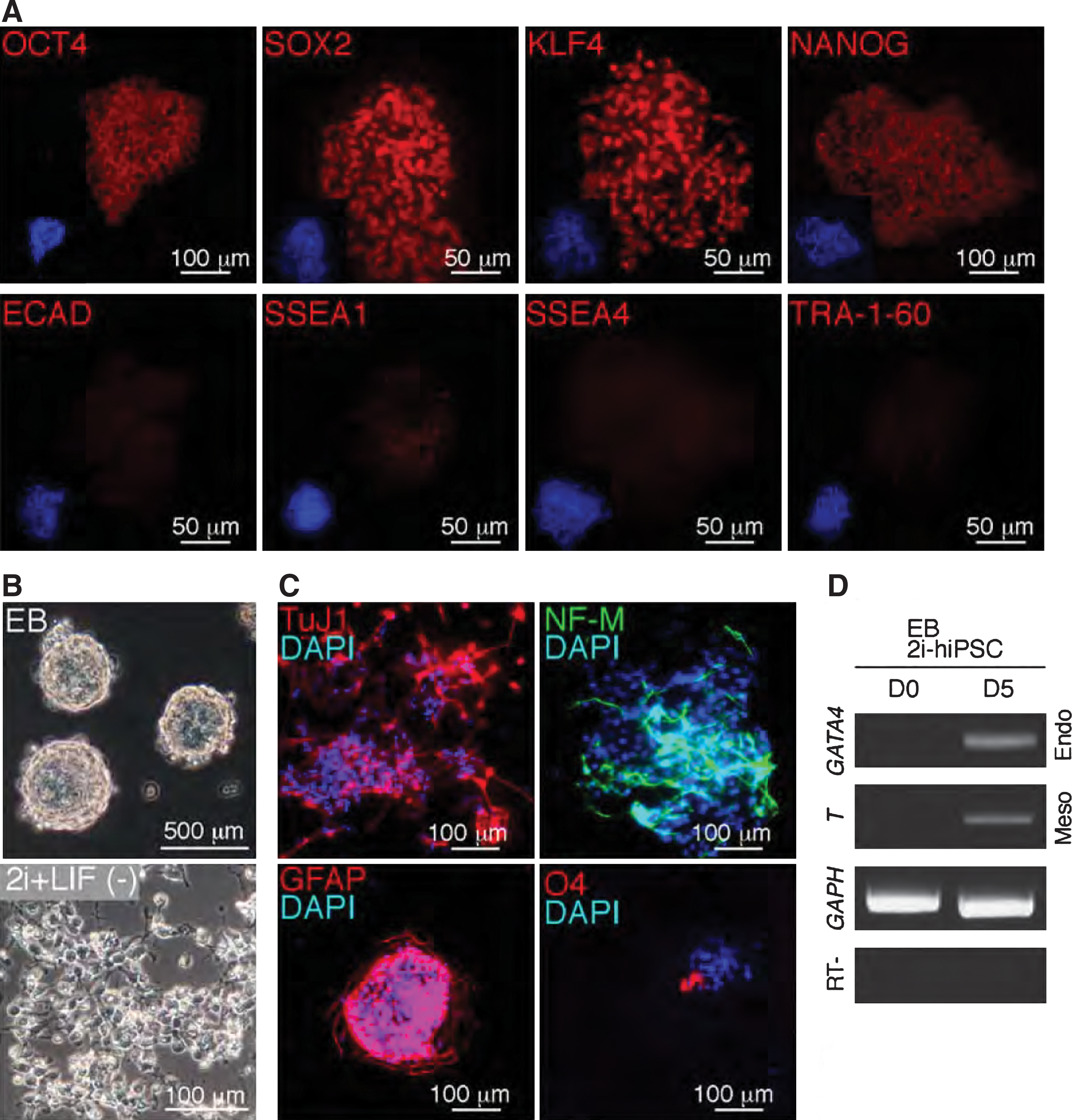
Characterization of differentiated and undifferentiated 2i-hiPSCs in vitro.
To determine the type of neural cells, we performed immunostaining on the differentiated cells. TuJ1 and NF-M-positive neuronal cells and GFAP-positive astrocytes were detected at a high frequency. O4-positive oligodendrocytes were also observed, albeit at a much reduced frequency (Fig. 5C). Thus, 2i-hiPSCs have the potential to differentiate into all types of neurons, astrocytes, and oligodendrocytes, as previously shown with mouse PNSCs [14]. Multi-lineage differentiation of 2i-hiPSCs was verified by expression of mesodermal (T) and endodermal (GATA4) lineage marker genes in day 5 EBs (Fig. 5D). Experiments to demonstrate contribution of 2i-hiPSCs into normal embryonic development were not approved on ethical grounds, although efficient contribution of PNSCs into aggregation chimeras in mice suggests that 2i-hiPSCs may well have the capacity to do so [26]. As an alternative approach, 2i-hiPSCs were implanted into kidney capsules of immunodeficient SCID mice to assess their in vivo teratoma forming ability. Notably, no teratomas was observed, from 10 independent implantations of undifferentiated and EB-formed 2i-hiPSCs, thus indicating that 2i-hiPSCs have considerably reduced potential for teratoma formation than their parental cells (Supplementary Fig. S3A), in contrast to teratoma formation with parental conventional hiPSCs (Supplementary Fig. S3B). Thus, we conclude that multi-potent 2i-hiPSCs that are primitively committed to neural differentiation, similar to PNSCs in mice, may represent a suitable source of personalized replacement cells in practical regenerative medicine.
Discussion
We demonstrate here that a culture with 2i+LIF medium induces reprogramming of flat-shaped human iPSCs into bowl-shaped PNSCs, which contrasts markedly to reprogramming of mESCs, iPSCs, and EpiSCs into mouse bowl-shaped GS PSCs. Human iPSCs were different from miPSCs even with the same minimal extrinsic stimuli in 2i+LIF medium (Fig. 6). In humans, 2i-hiPSCs resemble PNSCs in gene expression profile and multipotency. They can be stably expanded on matrigel-coated dishes without feeder cells by mechanical dissociation for >50 passages. Generation of 2i-hiPSCs from conventional hiPSCs, but not the hESCs we used, reflects a tendency for hiPSCs toward neuroectodermal lineages. Multi-potential 2i-hiPSCs, capable of differentiating into neurons, astrocytes, and oligodendrocytes in vitro, did not form teratomas in vivo. Together, the results indicate that 2i-hiPSCs are a promising source of cells for human neural research, pharmaceutical screening, and regenerative medicine.
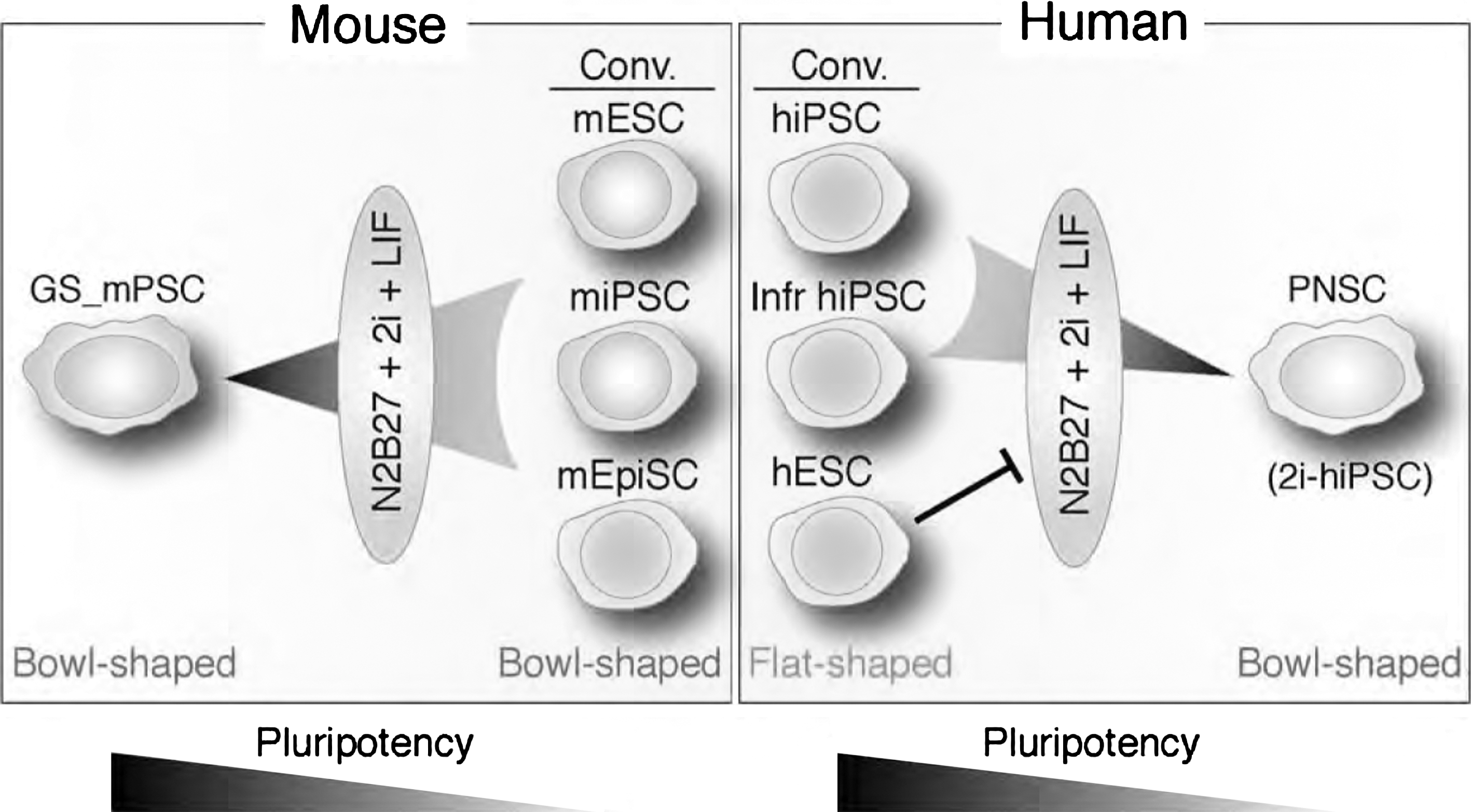
Schema of differential emergence of PSCs in 2i+LIF medium. In humans, PNSC emerge from conventional (Conv.) and Infr hiPSC, but not hESC, whereas mouse GS PSCs are generated from mESC, miPSC, and mEpiSC.
Difference between human and mouse PSCs in 2i+LIF medium
In rodents, 2i+LIF culture enriches GS PSCs characterized by uniform stem cell identity and germline competence [1,3,4]. This suggests that neuroectodermal differentiation induction by neurobasal N2B27 medium and differentiation inhibition by 2i and LIF are functionally balanced for tipping toward stemness defined by prolonged self-renewal in the undifferentiated state [1]. The same 2i+LIF culture conditions induced a multi-potent PNSC state with conventional hiPSCs, differing substantially from the pluripotent ES state seen with conventional mEpiSCs (Fig. 1A) [21,22]. The differences could be explained by at least 2 possibilities: (1) hiPSCs and mEpiSCs contain different subpopulations of cells that preferentially respond to the 2i+LIF culture condition, and (2) although hiPSCs and mEpiSCs exhibit similar flat-shaped colony morphology in a conventional state, hiPSCs retain a uniquely different molecular network signature that serves to promote neural differentiation in 2i+LIF medium.
In conventional and Infr iPSC populations, isolation of human PNSCs with 2i+LIF medium demonstrates the possibility that subpopulations of cells directly reprogrammed into PNSCs are generated and stably maintained in human ES medium with bFGF, which is required for primary neural specification [15]. These PNSCs, directly reprogrammed from somatic cell by reinforced transient expression of OSK, are selected and expanded in 2i+LIF medium. This could account for isolation of human PNSCs from hiPSCs but not hESCs.
In mouse PSCs, reprogramming of ESCs into PNSCs can be enhanced by inhibition of TGF β -related signaling mediated by substances including bone morphogenetic proteins (BMPs) as a default manner of neural fate specification [26]. Consistent with this, MEK inhibitor PD0325901, inhibiting neural differentiation through impeding ERK phosphorylation, functions to suppress neural commitment via the BMP pathway [1]. Therefore, another explanation is differential responses between mouse PSCs and hiPSCs to the extracellular signals introduced in 2i+LIF medium. Commitment to neural differentiation occurs in fully reprogrammed pluripotent hiPSCs by imperfect inhibition even in the presence of PD0325901. If so, then commitment of PSCs to form neural cells may be negatively regulated by not only the TGF β-related signaling pathway but also other signaling pathways in the humans, as demonstrated by differential fate determination in mEpiSCs and hESCs [27] and neural conversion of hESCs and hiPSCs using the dual-SMAD-inhibition protocol [28]. Recently, it was reported that hESCs were rapidly converted to homogenous PNSCs by a synergistic effect of inhibitors to GSK3, TGF β, and Notch signaling pathways [29]. hESC (H1, HUES9, and HUES1)-derived PNSC lines efficiently (>80%) generated by 1 week culture with N2B27 supplemented with CHIR99021, SB431542, and LIF were characterized by intensive analyses of expression of rosette-type neural stem cell (NSC), central nerve system NSC, anterior neural, and midbrain marker proteins by immunostaining, and midbrain marker genes by RT-PCR, and functional assays in vitro and in vivo. 2i-hiPSCs resemble hESC-derived PNSCs [29] in the expression profile of marker genes and proteins (Supplementary Fig. S4). Differential requirements between human and mESCs for differentiation into PNSCs may reflect a distinct physiology between mouse and hESCs.
Difference between human iPSCs and ESCs in 2i+LIF medium
Human iPSCs are functionally and molecularly indistinguishable from hESCs on the basis of multipotency, global gene expression, and DNA methylation profile [5,6,30]. Recent intensive research, however, demonstrated differential gene expression and DNA methylation between ESCs and iPSCs. Early passage iPSCs retained epigenetic memory inherited from their somatic cell origin, and dominantly differentiated into the cell type of origin. However, somatic epigenetic memory disappeared on continuous sub-culturing for both the human [31] and mouse cells [32]. Further analyses of whole-genome profiles of DNA methylation revealed that substantial hyper- and hypo-methylated CpG islands (differentially methylated regions) generated by aberrant and incomplete reprogramming existed in hiPSCs compared with hESCs [33,34]. Therefore, differential differentiation potentials between hiPSCs and hESCs in 2i+LIF medium may reflect differential epigenetic memory generated through inadequate reprogramming of somatic cells into hiPSCs. Here, we have ruled out the possibility of residual somatic epigenetic memory by using late-passage hiPSCs derived from 2 different somatic cell types. It is, however, possible that the hiPSCs cell lines we used are primed for differentiation into neuroectodermal lineages by DNA hypo-methylation of the promoter regions of key genes for neural differentiation. This, however, is not consistent with previous reports of low differentiation efficiency of hiPSCs into neural cells [35]. If so, then 2i-hESC lines may be established from some hESC lines, which have robust potentials of differentiating into neural cells [34] in 2i+LIF medium.
Stable maintenance of multipotent 2i-hiPSC lines
An intriguing property of 2i-hiPSCs is lack of expression of SSEA4 and E-CADHERIN, which is different from conventional hESCs and hiPSCs (Fig. 5A). In conventional hESCs, Ca2+-dependent E-CADHERIN-mediated cell-to-cell adhesion prevents apoptosis by ROCK/myosin hyper-activation [36]. No improvement in survival and proliferation of 2i-hiPSCs was detected with the treatment of ROCK inhibitor Y27632, or Myosin IIA inhibitor Blebbistatin [23,24] through subculture by enzymatic dissociation (data not shown). This is consistent with our observation that 2i-hiPSCs can be stably maintained by mechanical dissociation into 40–50 cell clusters but not enzymatic dissociation into single cells. However, it is consistent with no expression of E-CADHERIN in 2i-hiPSCs. Thus, it is speculated that other cell-to-cell adhesion molecules involved in inhibition of apoptosis, such as N-CADHERIN [29,37], may play an important role in survival of 2i-hiPSCs and human PNSCs in 2i+LIF medium. The putative role of cell adhesion molecule function, in maintaining a multipotent state of 2i-hiPSCs, is supported by induction of neural differentiation in chemically defined serum-free, feeder-layer free, and low-density culture conditions [26] and in enzymatic dissociation into single cells in 2i+LIF medium (data not shown).
Establishment of authentic human iPSCs from conventional hiPSCs
A few studies demonstrated that naïve hESCs or hiPSCs, which are an authentic counterpart of mESCs, could be generated but not maintained during long-term in vitro culture [12,13]. Naïve hESCs can be defined by bowl-shaped colony morphology, 2 active X chromosomes in female cells, pluripotency, and gene expression and epigenetic profile closely similar to those of mESCs. With regard to X-chromosome state, 2 active X chromosomes in an XX female cell are a hallmark of naïve ESCs in the mouse. In humans, one of 2X chromosomes is inactivated in conventional hESCs [20,38]. Interestingly, reducing the oxygen concentration from conventional 20% to physiological 5% in culture induces derivation of hESCs retaining 2 active X chromosomes [39]. In miPSCs, it is evident that an adequate culture condition enables establishment of naïve iPSC lines from various types of somatic cells by forced expression of transcription factors, OSKM or OSK, thus suggesting that transient expression of the transcription factors is sufficient for generating naïve hiPSCs with 2 active X chromosomes in females. This suggests that naïve hiPSCs could be established if appropriate specialized culture conditions were found and applied, that differ from current hESC culture conditions with KSR and bFGF. Establishment of naïve hiPSCs would provide a significant contribution to the study of mechanisms involved in early human embryonic development and regenerative medicine.
Footnotes
Acknowledgments
The authors thank Dr. Gen Kondoh and Ms. Hitomi Watanabe for generating chimeras, Dr. Kathrin Plath for the probe DNA to human XIST, and Dr. Takashi Sado for XIST RNA FISH. This work was partly funded by grants from the Japan Society for the Promotion of Science, the Ministry of Education, Culture, Sports, Science, and Technology and the Core Research for Evolutional Science and Technology (Japan Science and Technology Agency) to T.T. K.H is a fellow of JSPS.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.