
Abstract
Understanding the mechanism by which embryonic stem (ES) cells self-renew is crucial for the realization of their therapeutic potential. Earlier, overexpression of Id proteins was shown to be sufficient to maintain mouse ES cells in a self-renewing state even in the absence of serum. Here, we use ES cells derived from Id deficient mice to investigate the requirement for Id proteins in maintaining ES cell self-renewal. We find that Id1−/− ES cells have a defect in self-renewal and a propensity to differentiate. We observe that chronic or acute loss of Id1 leads to a down-regulation of Nanog, a critical regulator of self-renewal. In addition, in the absence of Id1, ES cells express elevated levels of Brachyury, a marker of mesendoderm differentiation. We find that loss of both Nanog and Id1 is required for the up-regulation of Brachyury, and ectopic Nanog expression in Id1−/− ES cells rescues the self-renewal defect, indicating that Nanog is the major downstream target of Id1. These results identify Id1 as a critical factor in the maintenance of ES cell self-renewal and suggest a plausible mechanism for its control of lineage commitment.
Introduction
B
Early research on Nanog demonstrated that the majority of ES cells that lack Nanog quickly differentiate into endoderm, although it was noted that a small sub-population of cells could be maintained and passaged continuously in culture [9,10]. Subsequent work using Nanog heterozygote ES cells or shRNA knockdowns also demonstrated that having insufficient amounts of Nanog made maintaining ES cells in culture difficult, due to the propensity of such cells to differentiate [11 –13]. Recently, it was reported that normal ES cells, grown in self-renewing conditions, transiently turn off Nanog expression. These studies also showed that Nanog−/− ES cells could still self-renew and differentiate into cell types of all 3 germ layers, suggesting that Nanog was not absolutely required for self-renewal, but rather functioned in a rheostat-like manner to safeguard self-renewal and pluripotency of ES cells [14]. Nanog works in combination with Oct4 and Sox2, each regulating the other's expression, as well as that of a variety of other factors involved in maintaining pluripotency and a self-renewing state [4]. However, how these “master regulators” are themselves controlled and what is the identity of their downstream effectors are unresolved questions of considerable interest.
Id proteins regulate the differentiation programs of a variety of cell types. In neural stem cells and progenitors, they inhibit differentiation and promote proliferation [15 –18]. In hematopoietic stem cells, Ids are required to maintain multipotency, as Id1 loss leads to premature myeloid differentiation and reduced self-renewal [19 –22]. In ES cells, BMP4 stimulates the expression of Ids via Smad signaling, and thereby blocks neuroectoderm differentiation [23].
We set out to determine whether Id proteins are required to maintain ES cell self-renewal and inhibit differentiation. We show that in ES cells, loss of Id1 is sufficient to tip the balance from self-renewal to differentiation. The absence of Id1 results in down-regulation of Nanog expression, leading to decreased ES cell self-renewal. In addition, we find that in ES cells, Id1 plays a role in the negative regulation of Brachyury, a T-box transcription factor and early marker of mesendoderm and mesoderm [24], as Id1−/− ES cells express elevated amounts of Brachyury, levels of which correlate with an increased propensity for cellular differentiation. In conjunction with Nanog, Brachyury has been shown to regulate the balance between self-renewal of ES cells and differentiation into early mesendoderm [25,26], and we find that the loss of both Nanog and Id1 are necessary for the up-regulation of Brachyury expression.
Thus, loss of Id1 expression results in both the down-regulation of Nanog and the up-regulation of Brachyury expression, and these changes are responsible for the observed decrease in self-renewal and the propensity to differentiate. These results establish a requirement for Id1 in maintaining ES cell self-renewal and suggest an unanticipated role in its control of lineage commitment.
Materials and Methods
ES cell lines and culture
Wildtype ESD3 ES cells were obtained from ATCC. Wildtype, Id1−/− , and Id3−/− ES cells were derived from wildtype and knockout mice using protocol as previously described [27]. Essentially, mice with the appropriate genotypes were crossed and plugs were checked each morning. The day of the observed plug was considered d0.5. On d3.5, a pregnant female was sacrificed; the uterus was removed and rinsed in phosphate buffered saline (PBS). The uterus was cut in half, separating the 2 horns of the uterus. Each horn was flushed with M2 media (Sigma M7167), and flushed media was collected in pools on sterile tissue culture grade plates. Pools of media were inspected for blastocysts using a dissection microscope, and identified blastocysts were transferred via a very finely drawn transfer pipette to mitomycin C treated mouse embryonic fibroblast feeder cell-coated wells of a 96-well tissue culture treated microtiter plate, one blastocysts per well. Media in well was ES cell media [Dulbecco's modified Eagle medium (DMEM) supplemented with 15% ES cell-qualified fetal bovine serum (FBS), 1,250 U/mL leukemia inhibitory factor (LIF), 140 μM 2-mercaptoethanol, 1×MEM nonessential amino acids, 2 mM L-glutamine, and 1×Pen-Strep].
Media on blastocysts remained unchanged for 3 days as blastocysts swelled, hatched from zona pellucida, and attached to feeder cell-lined plate. By day 5 post-transfer, when the trophoblast cells had spread out and the inner cell mass (ICM) was visible as a mound of cells, cells were trypsinized to break up the ICM, and transferred to a new feeder-cell lined well. Wells were monitored daily and passaged regularly until ES cell colonies, based on morphology, were visible or until ∼25 days had passed.
Wells containing ES cell colonies were passaged into larger wells, as required, and ultimately frozen down in ES cell freezing medium (80% ES cell media, 10% additional ES cell-qualified FBS, and 10% DMSO).
ES cells were maintained on gelatin, without feeders, in DMEM supplemented with 15% ES-cell qualified FBS, 1,250 U/mL LIF, 140 μM 2-mercaptoethanol, 1×MEM nonessential amino acids, 2 mM L-glutamine, and Pen-Strep. RUES1 human ES cell line was maintained as previously described [28].
Tet-inducible ES cell line was generated by introducing a CAGGS-rtTA-IRES-Puromycin construct (generously provided by D. Melton, developed as previously described [29]) into wt ES cells via electroporation. The Tet-on Id1 shRNA construct and Luciferase control shRNA construct (generously provided by V. Mittal, developed as previously described [30]) were introduced via lentiviral transduction.
Proliferation rate
Proliferation rate was measured by seeding a fixed number of cells (4×104 cells/cm2) on gelatin-coated plates, in triplicate, and growing for 48 h. Cells were trypsinized and counted using a haemocytometer and tryphan blue. Fixed number of cells was reseeded, and this process was repeated over multiple passages.
Self-renewal assays
Single-cell cloning self-renewal assay: individual cells were plated, via limited dilution, into gelatinized wells of 96-well plate. After 8 days of growth, individual self-renewing colonies, as judged by morphology, were trypsinized, and cells were counted and reseeded as individual cells into gelatinized 96-well plates. After 8 days, colonies with a self-renewing appearance were counted.
Colony-forming assay: cells were trypsinized to a single-cell suspension and replated at 600 cells per 10 cm2 well, in triplicate. After 6 days, plates were stained for alkaline phosphatase following directions of the AP stain kit (Chemicon) and scored for differentiation status.
To compare the level of self-renewal in the Id1−/− ES cells shown in Fig. 2E, to previously published results [14], the number of self-renewing colonies generated was multiplied by the ratio of self-renewing colonies formed from the wildtype cells in the previously published work to the number of wildtype self-renewing colonies generated in our assay.
Microarray
Microarray was performed using total RNA extracted via Rneasy kit (Qiagen) and MOE430A (Affymetrix) chip.
Expression and shRNA plasmids
Expression plasmids utilized the pWPI plasmid backbone containing a GFP marker, from D. Trono's lab. Brachyury, Nanog, and human Id1 shRNA constructs utilized the pLKO vector [31] containing a puromycin resistance gene. ShRNA sequences are listed in Table 1 below.
To produce virus, 293T cells, switched to ES cell media, were transfected with VSVG, delta8.9, and shRNA or expression plasmids via calcium phosphate precipitation transfection. 36 h posttransfection, virus-containing media was harvested from transfected 293T, filtered, and added, with polybrene, to the target ES cells. Transduction of target cells was repeated thrice over a 24-h period. 48 h postlast transduction, positively transduced ES cells were selected for either by FACS, selecting for GFP, or via puromycin resistance. Puromycin selection was performed by growing cells in 2.5 μg/mL puromycin for 48 h.
Reverse transcriptase-polymerase chain reaction
Total RNA was generated using Rneasy kit (Qiagen) and reverse transcribed using Superscript III first strand cDNA kit and random hexamer primers (Invitrogen). Polymerase chain reaction (PCR) primers are previously published, as noted, or generated using the Primer3 program [32].
Previously published primers are Brachyury and Flk-1 [33]; Sox2, Stella, and Rex1 [14]; Sox17 and Pax6 [34]. Additional primers listed in Table 2.
Western blots
Western blots were performed using 50 μg whole cell lysate generated as previously described [35]. Membranes were blocked in 5% milk PBS-T (0.05% Tween-20), and antibody incubations were performed in 1% milk PBS-T at dilutions indicated: mouse Nanog (Abcam #ab21603) [1:1,000], human Nanog (Abcam # ab21624) [1:1,000], Oct-3/4 (BD #611202) [1:1,000], Brachyury (R&D systems #af2085) [1:1,000], Id1 (Biocheck) [1:5,000], Id3 (Biocheck) [1:5,000], and Actin (Sigma #A5060) [1:5,000].
Statistical analysis
Data presented are averages±standard deviation. When comparing 2 data sets, P values were generated using Student's t-test. P<0.05 was considered significant.
Results
Id levels are transiently down-regulated during embryoid body differentiation
Id mRNA levels are elevated in self-renewing ES cells and decrease on differentiation (e.g., in the absence of LIF [23]). As in development, on embryoid body formation, the levels of Ids vary with time: levels of Id1 and Id3 decrease by day 1 postinduction, reach a minimum around day 3, and then return to base line by day 5. Levels of Id2 remain constant during the first 2 days of differentiation and then decline by day 3, similarly recovering to baseline by day 5. Id4 levels increase initially on the first day of differentiation, decline on day 2 and day 3, and then recover fully by day 4 (Fig. 1A). Accordingly, on embryoid body formation, protein levels of Id1 are high in undifferentiated ES cells, noticeably decrease by day 1 of differentiation, remain low for the next 2 days, and then return to base line by day 5 (Fig. 1B). The differential expression of the 4 Ids suggests that in ES cells, as in development, they may have functionally distinct roles.
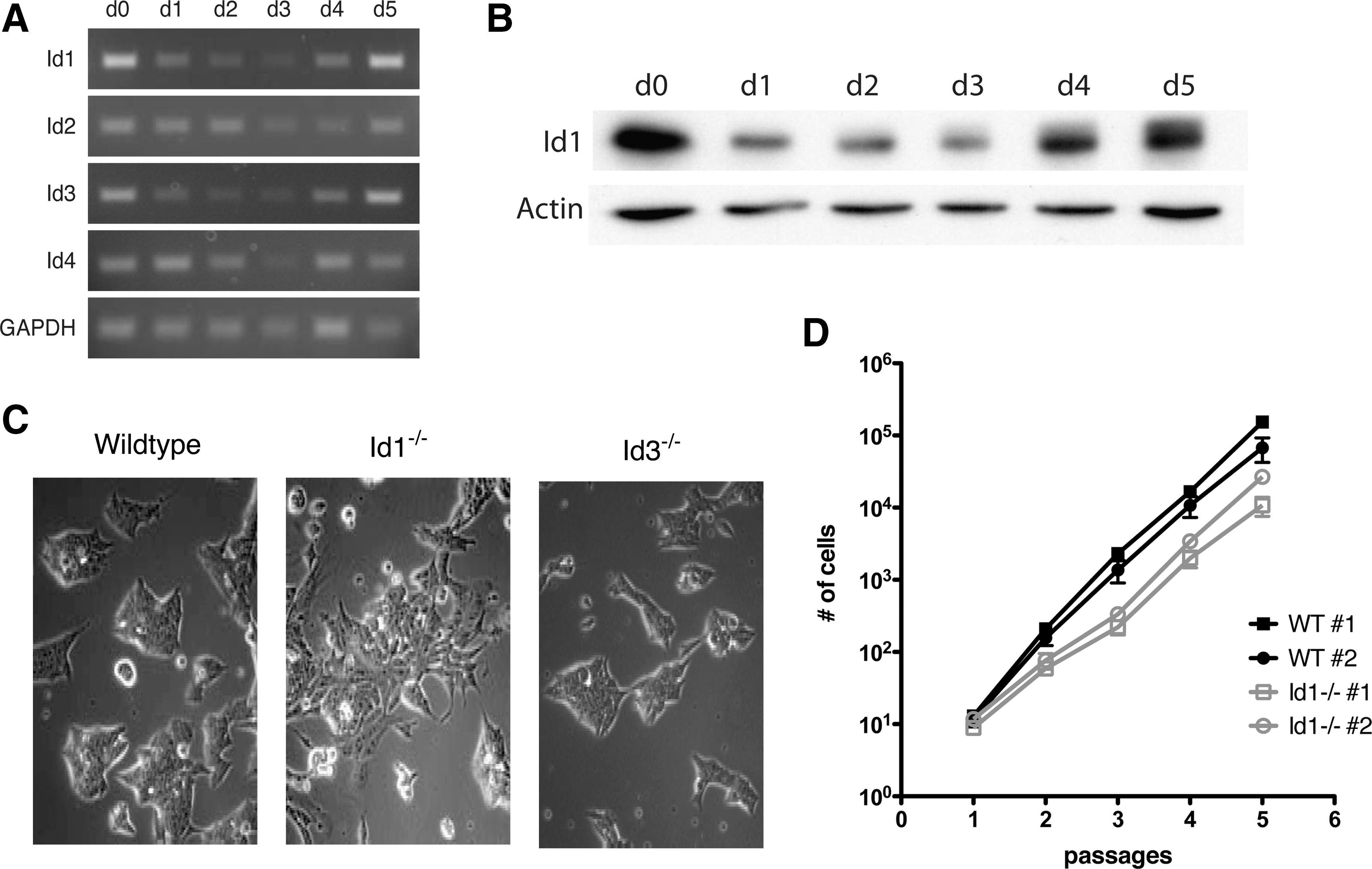
Expression of Ids is down regulated during differentiation. Cells devoid of Id1 exhibit a changed morphology and reduced rate of proliferation.
Id1−/− ES cells have an altered morphology and growth rate
In ES cells grown in the presence of LIF but in the absence of serum or BMP4, over-expression of Ids is sufficient to maintain ES cells in a self-renewing state [23]. To understand the individual role of Id proteins in ES cells, Id1- and Id3-deficient ES cells were derived from the ICM of d3.5 blastocysts obtained from Id1- and Id3-knockout mice, generated as previously described [36,37]. Id1−/−
ES cells grown on gelatin (Fig. 1C) displayed a more flattened and spread out appearance when compared with wildtype or Id3−/−
ES cells. Additionally, 2 independently derived Id1−/−
ES cell lines with normal karyotypes could be expanded over multiple passages, but had a slower growth rate compared with wildtype ES cells (Fig. 1D, Supplementary Table S1; Supplementary Data are available online at
Self-renewal factors are altered in Id1−/− ES cells
Gene expression profiles of Id1−/− and wildtype ES cells were compared via microarray analysis. Of the core factors previously implicated in the regulation of self-renewal, Oct4, Sox2, and Nanog, only the change in Nanog mRNA levels was significant (GEO series #GSE19144). These observations were confirmed by reverse transcriptase-PCR (RT-PCR) (Fig. 2A). Id1−/− cells expressed Stella (Dppa3) (Fig. 2A), confirming that they are indeed ES cells and not epiblast cells [38]. The microarray results, confirmed by RT-PCR also indicated that Id1−/− ES cells expressed lower levels of Rex1 and Rex2 (Fig. 2A), 2 factors used as markers of pluripotency [39 –41], suggesting that the Id1−/− ES cells may be differentiating spontaneously.
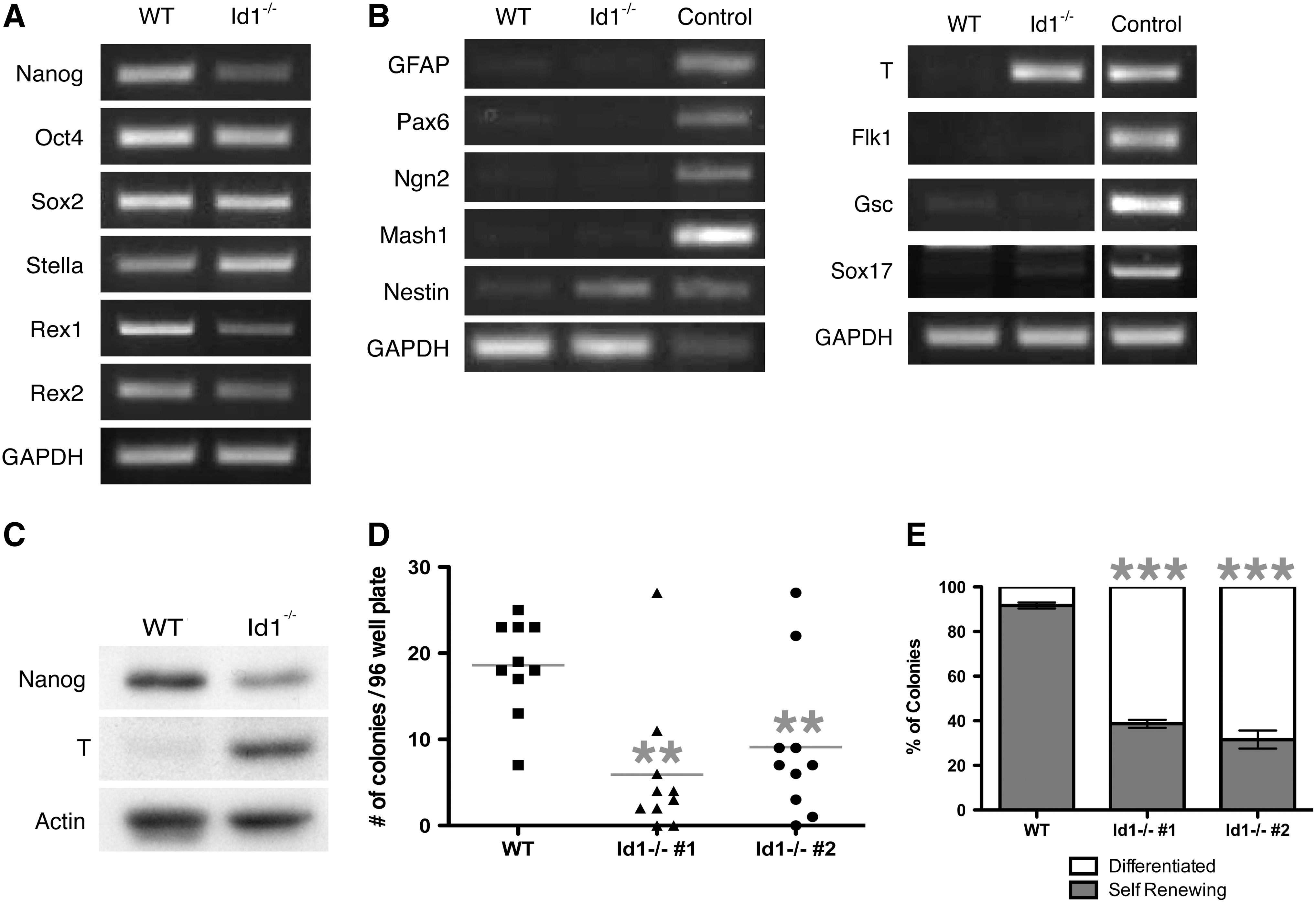
Id1−/−
ES cells have a defect in self-renewal.
No up-regulation of GFAP, a marker of neural differentiation, was observed in the Id1−/− ES cells. There was also no up-regulation of Neurogenin 2 or Mash1, 2 neural specific bHLH factors, in the Id1−/− cells (Fig. 2B). Id1−/− cells did, however, express a low level of Nestin (Fig. 2B), an early marker of neural differentiation that is also expressed in early muscle progenitors and epithelial derivatives [42].
In contrast, relative to wildtype ES cells, Id1−/− ES cells grown under self-renewing conditions expressed significantly higher levels of the T-box transcription factor Brachyury, a marker of mesendoderm differentiation (Fig. 2B). Flk1, a gene expressed later in mesoderm differentiation, as well as Goosecoid and Sox17, markers of endoderm, were not expressed in Id1−/− ES cells, suggesting that the Id1−/− cells may be at a very early stage of mesendoderm differentiation (Fig. 2B).
In agreement with the RNA data, Nanog protein levels in Id1−/− ES cells were reduced from approximately one half to one third of the level observed in wildtype cells, and Brachyury protein expression was robust in Id1−/− ES cells, while very low in wildtype ES cells (Fig. 2C).
Id1−/− ES cells have a decreased ability to self-renew
Id1−/− ES cells demonstrated traits that suggested they might be differentiating and losing their capacity to self-renew. To quantitate self-renewal, a single-cell cloning self-renewal assay was performed where single cells from individual colonies were seeded into 96-well microtiter plates; at day 8, the resulting ES cell colonies were counted and assessed morphologically. As shown in Fig. 2D, Id1−/− ES cells formed ∼40% of the number of self-renewing colonies generated when using wildtype ES cells.
These results were confirmed using an alkaline phosphatase activity assay. For each cell line examined, 600 cells were seeded in triplicates and allowed to form colonies. Self-renewing colonies have a distinct and compact structure that stains strongly for alkaline phosphatase activity; more differentiated colonies have a spread-out morphology and very little or no alkaline phosphatase activity. Fewer than 40% of the colonies formed from the Id1−/− ES cells were self-renewing, compared with more than 90% of the colonies from wildtype cells (Fig. 2E). Despite being grown under conditions that support self-renewal, Id1−/− ES cells demonstrated a propensity to differentiate, indicating that ES cells require Id1 to maintain a normal rate of self-renewal.
To demonstrate that loss of self-renewal was Id1 dependent, we reintroduced expression of Id1 into the Id1−/− ES cells via lentiviral transduction, and assayed for self-renewal via alkaline phosphatase staining. As seen in Supplementary Fig. S1, transducing the cells with Id1 partially rescued the self-renewal defect. We find that reintroducing Id1 expression in the Id1−/− ES cells significantly increased the number of self-renewing colonies generated from the Id1−/− ES cells (i.e., doubling the number of self-renewing colonies generated when Id1 −/− ES cells were transduced with an Id1 expressing construct as compared to control transduced ES cells). We hypothesize that the limitation of the rescue may have been due to some of the Id1−/− ES cells having already been committed to differentiate at the time of transduction or perhaps, because the transduced cells do not have properly regulated levels of Id1.
Nanog levels are reduced by acute loss of Id1
Id1−/− ES cells expressed reduced levels of Nanog. To determine whether acute Id1 loss similarly led to down-regulation of Nanog, a tetracycline-inducible, Id1-specific short hairpin RNA (Id1-shRNA) was introduced into wildtype ES cells expressing the reverse tetracycline transactivator (rtTA). Addition of doxycycline to the media resulted in a dose-dependent reduction of Id1 protein levels (Fig. 3A), which were reduced by ∼20%. Knockdown of Id1 resulted in 30% down-regulation of Nanog protein levels (Fig. 3A), confirming that, in mouse ES cells, Id1 plays a role in maintaining normal Nanog expression.
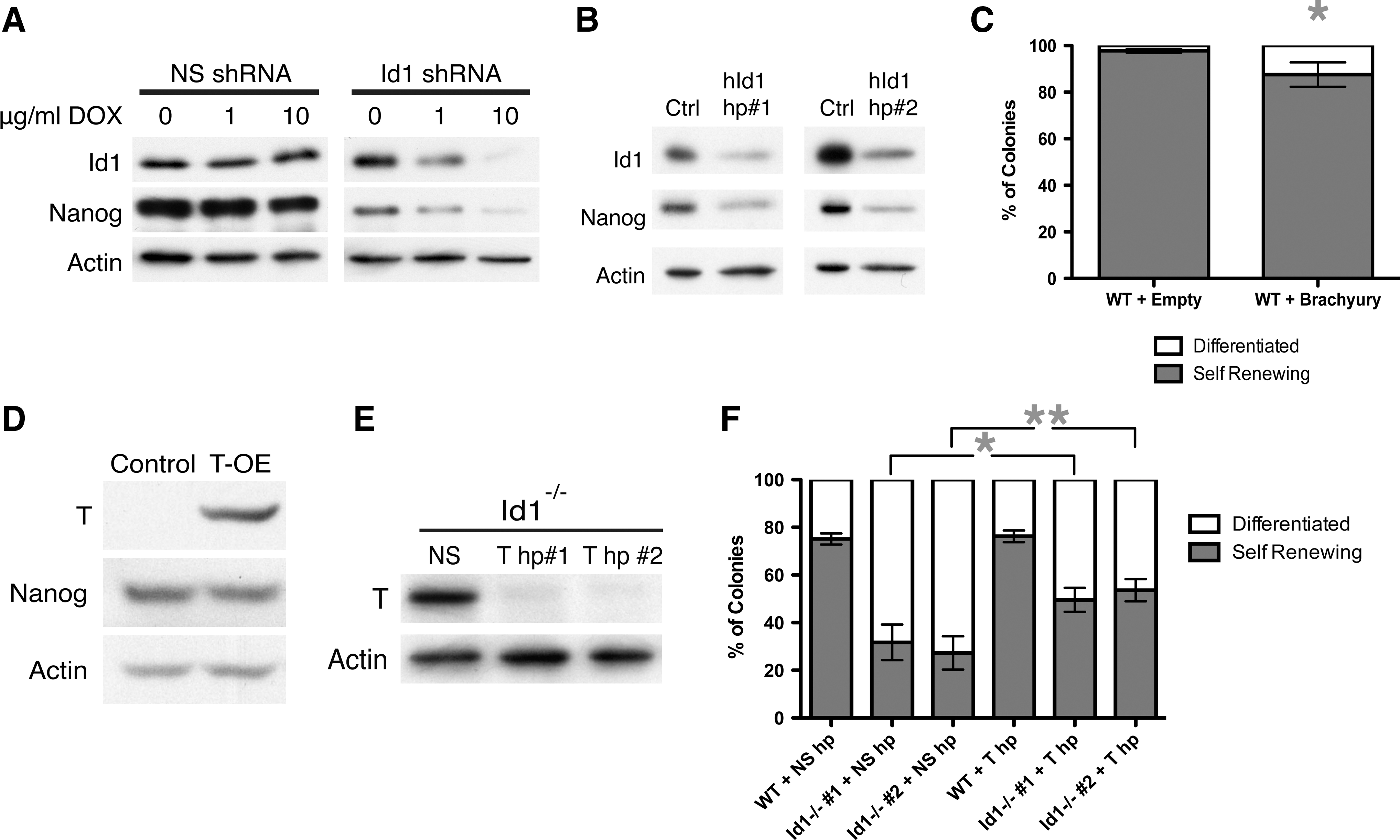
Brachyury expression results in a decrease of self renewal.
To determine whether the loss of Id1 may have a similar effect in human ES cells compared with mouse ES cells, we introduced Id1-specific shRNA constructs or a nonspecific shRNA negative control into human ES cells via lentiviral transduction. As seen in Fig. 3B using 2 different Id1-shRNAs, reduction of Id1 to ∼40% of normal results in decreased Nanog levels that are ∼35% of normal expression, indicating that in human ES cells Id1 expression also plays a role in maintaining Nanog expression.
Previous work demonstrated that the ability of ES cells to self-renew decreased as a function of Nanog expression levels. Approximately 70% of the colonies generated from wildtype cells were self-renewing, versus 50% of the colonies formed from Nanog+/− cells, and 25% of the colonies formed from cells were devoid of Nanog [14]. Id1−/− ES cells self-renewed at levels comparable to that of Nanog null cells, despite expressing levels of Nanog RNA and protein roughly comparable to Nanog+/− cells. This suggested that another factor might play a role in the Id1−/− ES cells' reduction of self-renewal.
Brachyury over-expression leads to a reduction in self-renewal
As earlier shown, Id1−/− ES cells markedly up-regulate Brachyury mRNA and protein levels. Nanog can repress Brachyury expression [26]; however, Nanog−/− ES cells do not display elevated Brachyury levels [9,14], nor were Brachyury protein levels elevated in wildtype cells when Nanog levels were reduced by more than 90% via Nanog specific-shRNAs (data not shown). Thus, the reduction in Nanog by itself is not sufficient to induce Brachyury expression, and there should be a Nanog-independent mechanism leading to Brachyury up-regulation in Id1−/− ES cells.
Brachyury is a known marker of mesendoderm differentiation and a transcription factor that, in conjunction with changes in Nanog expression, has been previously implicated in regulating the balance between self-renewal of ES cells and differentiation into early mesendoderm [25,26]. We, therefore, tested the hypothesis that the elevated levels of Brachyury found in our Id1−/− ES cells play a role in the self-renewal defect of these cells.
Wildtype ES cells transduced with lentivirus encoding either Brachyury, upstream of an IRES-GFP, or the empty vector control, were sorted for GFP, and only GFP+ cells were propagated in culture. A small but reproducible and significant decrease in the ability of the Brachyury/GFP+ cells to self-renew was observed (Fig. 3C). However, Brachyury over-expression did not alter the levels of Nanog (Fig. 3D), indicating that the decrease in self-renewal due to Brachyury over-expression was independent of Nanog levels.
Reducing Brachyury levels in Id1−/− ES cells partially rescues their ability to self-renew
To test whether the expression of Brachyury in Id1−/−
ES cells decreased the ability of these cells to self-renew, a Brachyury-specific shRNA or a nonspecific shRNA control was introduced into the Id1−/−
ES cells via lentiviral transduction. A greater than 90% knockdown of Brachyury was achieved using 2 independent Brachyury-specific shRNAs (Fig. 3E). Id1−/−
ES cells in which Brachyury levels had been reduced generated 1.5-fold more self-renewing colonies than Id1−/−
ES cells transduced with a nonspecific-shRNA, while introducing a Brachyury-specific shRNA into wildtype ES cells (where Brachyury is undetectable) had no effect (Fig. 3F). Thus, on Brachyury reduction, the self-renewal capacity of Id1−/−
ES cells was restored to about 50% of the self-renewal capacity of wildtype ES cells, which is roughly equivalent to that of Nanog+/
Increasing levels of Nanog in Id1−/− ES cells rescues the defect in self-renewal
To test whether the reduction in self-renewal of Id1−/− ES cells could be rescued by increasing Nanog levels, exogenous Nanog was introduced into the Id1−/− ES cells. Id1−/− ES cells over-expressing Nanog formed colonies that were morphologically indistinguishable from wildtype ES cells transduced with an empty vector control (Fig. 4A).

Expression of Nanog rescues self-renewal defect.
No statistically significant difference was observed in the number of colonies generated from wildtype cells transduced with the Nanog expression vector or an empty vector control (Fig. 4B). In contrast, on average, over-expression of Nanog in Id1−/− ES cells increased the number of self-renewing colonies to numbers similar to that of wildtype ES cells (Fig. 4B).
Scoring for alkaline phosphatase positive colonies in a self-renewal colony-forming assay confirmed these results. Although the number of adherent colonies increased on transduction with Nanog, as has been previously seen [12,14], there was no significant difference in the relative number of self-renewing and differentiated colonies generated from wildtype cells transduced with the Nanog expressing vector or an empty vector control. In contrast, the number of self-renewing colonies generated from Id1−/− ES cells increased nearly 2-fold when Nanog was over-expressed, verifying that an increase in Nanog levels in the Id1−/− ES cells achieved a near complete rescue of self-renewal in these cells (Fig. 4C).
We observed that over-expressing Nanog in the Id1−/− ES cells resulted in a suppression of Brachyury (Fig. 4D), accounting for the ability of increased Nanog levels to nearly completely rescue the self-renewal capacity of Id1−/− ES cells.
Discussion
Previous work in ES cells showed that, in the presence of LIF, Id proteins are sufficient to maintain self-renewal by inhibiting differentiation into the neuroectoderm lineage, most likely by blocking precocious activation of neural lineage-specific bHLH transcription factors [23]. We show here that Id1 is necessary for ES cells to maintain their full capacity for self-renewal and provide a mechanistic understanding of how this is accomplished. Our results support a model whereby Id1 regulates self-renewal by driving Nanog expression and repressing Brachyury expression via a Nanog-independent mechanism (Fig. 5).
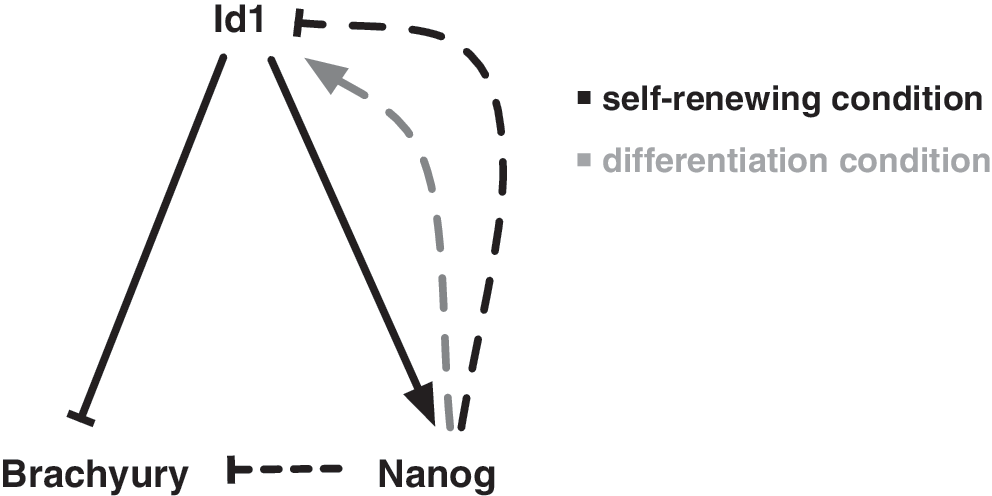
Id1 regulates self-renewal of ES cells by regulating levels of Nanog and Brachyury. ES cells have the ability to perpetually self-renew while retaining sensitivity to differentiation cues. Our work supports the model that Id1, whose levels are high in self-renewing ES cells and are down-regulated on differentiation, helps maintain ES cells by suppressing differentiation caused by Brachyury expression as well as supporting self-renewal by driving Nanog expression. It has been previously shown (indicated by dashed lines) that Nanog can repress Brachyury expression [26] and maintain levels of Id1 under differentiation conditions (gray arrow) [23] but represses Id1 when over-expressed under self-renewal conditions [25]. These feedback loops support a model in which Id1 is an important factor in maintaining ES cell self-renewal and suggest a plausible mechanism for its control of lineage commitment.
As previously seen, the loss of Nanog expression reduces the ability of ES cells to self-renew [14]. Thus, Id1's ability to regulate ES cell self-renewal is strongly linked to its regulation of Nanog expression levels. While Id1−/−
ES cells express Nanog at levels comparable to that found in Nanog+/
Elevated Brachyury expression is a marker of mesendoderm differentiation, but the previously identified role for Id1 in ES cells was to block neuroectoderm differentiation. Our results, however, are not inconsistent with the previously reported role for Ids: removal of serum or over-expressing inhibitory Smads in ES cells led to a reduction of multiple Ids and could have effects on other regulatory circuits. Over-expressing E-proteins presumably over-rode the inhibitory action of all Ids while also potentially affecting the expression of other E-box dependent transcription units. These changes promoted neuroectodermal differentiation [23], but the requirement for specific Ids in lineage commitment was not addressed. Here, Id1−/− ES cells were grown in normal self-renewing conditions (i.e., in the presence of serum and LIF). Therefore, as expected, other Id proteins were still expressed, and serum and LIF dependent events were unperturbed. Thus the specific loss of Id1, consequent reduction of Nanog expression, and release of Brachyury repression could drive mesendoderm differentiation while neuroectoderm differentiation is still repressed. By serving as a joint regulator of both Nanog and Brachyury expression, Id1 can serve as a central factor in the maintenance of ES cell self-renewal and in the regulation of the first steps of mesendoderm differentiation.
Interestingly, in ES cells grown in the absence of serum or BMP4 signaling (conditions where levels of Id1 would normally decrease), forced expression of Nanog maintains levels of Id1, and creates a positive feedback loop that supports ES cell self-renewal [23]. In contrast, we (data not shown) and others have found that over-expression of Nanog in ES cells grown under self-renewing conditions results in a reduction of Id1 [25], suggesting that overly high levels of Nanog under conditions of self-renewal may activate a negative feedback loop in an attempt to reduce levels of Nanog and reinstate a condition where the cells would be sensitive to differentiation cues.
These multiple feedback loops may play essential roles in regulating Nanog expression and demonstrate how Id1 may maintain a balance in the ability of ES cells to self-renew and their sensitivity to differentiation signals. They may define a transcriptional switch in self-renewing ES cells where the loss of the Id1 signal due to appropriate differentiation stimuli sets off the differentiation pathway: Nanog expression is repressed while Brachyury levels are up-regulated. Once Nanog levels are too low to maintain self-renewal, the cells become committed to differentiation. Even if Id1 levels rise, as they do during embryoid body formation, this may not be sufficient to reactivate the expression of Nanog.
Previous work has established Id proteins as inhibitors of differentiation. Our results demonstrate that, in ES cells, Id1 has a role not just in blocking differentiation to the mesendoderm lineage, but also in actively promoting self-renewal by driving Nanog expression. Thus, we identify Id1 as an important factor in the maintenance of self-renewal in ES cells and demonstrate an unanticipated mechanism for its control of lineage commitment. Further studies will be required to determine what signals induce a drop in Id levels, whether the signal reduces the level of a particular Id or all Ids in general, and how mechanistically the loss of Id1 leads to a reduction in Nanog expression.
Footnotes
Acknowledgments
The authors thank B Lanman and JM Schvartzman, as well as past and present members of the Benezra lab, for their helpful discussions and assistance in the preparation of this article. They thank H Kim and D James for their work with human ES cells, M Leversha and the Molecular Cytogenetics CF for assistance in karyotyping analysis, J Hendrikx, M Menon, and the FCCF for FACS sorting, and A Viale and the GCL for performing the microarray experiments. They also thank Vivek Mittal, Andrea Ventura, Hisato Kondoh, Yujiro Higashi, Douglas Melton, Dider Trono, Tyler Jacks, and David Root for sharing reagents. This work was supported in part by a grant from the C.V. Starr Foundation.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.