
Abstract
Human pluripotent stem cells (hPSCs), including human embryonic stem cells (hESCs) and human induced pluripotent stem cells (hiPSCs), share the properties of unlimited self-renewal and the capacity to become any cell type in the body, making them well suited for regenerative medicine and cell therapy. So far, almost all hPSC lines have been directly or indirectly exposed to animal-derived products, which would hinder their use for clinical purposes. One of the biggest challenges in this area is to remove animal components from the derivation, propagation, and cryopreservation of hPSCs. Moreover, the presence of undefined components of animal or human origin in culture system may interfere with the interpretation of the effect of exogenous agents on the growth and differentiation of hPSCs and are prone to significant variability. To explore hPSC expansion in defined, xeno-free conditions, 2 different groups of culture systems were used to culture different hESC and hiPSC lines. Our results suggested that (1) medium, matrix, and exogenous factors have synergistic effects on the adhesion and growth of hPSCs; (2) cooperation of exogenous factors including basic fibroblast growth factor, Rho-associated kinase inhibitor (ROCK), and other growth factors is critical for hPSC adhesion and proliferation; (3) basal media have different effects on hPSC attachment to the culture surface; and (4) a medium or matrix component can work synergistically in one culture system, and not at all in another. In this study, we found that Vitronectin/TeSR2 and PDL/HEScGRO (Y-27632) systems were optimal for maintaining the long-term culture of 3 hESC lines and 2 hiPSC lines under defined, xeno-free conditions.
Introduction
T
At present, 2 culture systems are being widely used to maintain hPSCs worldwide. One is a feeder-dependent culture system consisting of knockout serum replacement (KSR) medium and feeder cells, which either mouse embryonic fibroblasts (MEFs) or human fibroblasts. Another is feeder-free culture system consisting of mTeSR1 medium and Matrigel coating matrix. Both culture systems are not suitable for clinical trials or future applications, as KSR and mTeSR1 media contain a large amount of xenogeneic bovine serum albumin (BSA), and Matrigel, a reconstituted extracellular matrix (ECM) derived from Engelbreth-Holm-Swarm mouse tumor [12,13].
The presence of animal products in hPSC culture media, as well as feeder cell populations, drives 2 main concerns [14]. First, animal products may contain toxic proteins or immunogens that evoke an immune response and thus lead to rejection upon transplantation of hPSCs [15]. Second, the use of animal products increases the risk of hPSC contamination by the animal pathogens, such as viruses or prions [16].
The presence of undefined components of animal or human origin in these systems, such as feeder cells and Matrigel, is also problematic. First, unknown factors in Matrigel or produced by the feeder cells may interfere with the interpretation of the effect of exogenous agents on the growth and differentiation of the stem cells. Second, components such as Matrigel, unknown growth factors, and feeder cells are prone to significant variability [17]. To address these issues, purified proteins from human tissues, recombinant proteins, and synthetic matrices have been used as xeno-free, defined substrates in conjunction with several chemically defined culture media [18,19]. However, in these studies, animal component–containing media were still used. As an alternative, 1 group reported a novel method using poly-
The widespread clinical use of hPSC derivatives will require xeno-free derivation as well as passage, differentiation, and cryopreservation. Selection of xeno-free culture systems by long-term culture of hPSCs is a key step to find optimum environments for hPSC derivation. Based on these previous studies and our own work, we hypothesized that there may be synergies between specific media, matrix, and exogenous factors, which are critical for the adhesion and long-term self-renewal of hPSCs. It is hard to say which medium or matrix is optimal for hPSC growth in an undifferentiated state. A medium or matrix probably is not ideal in one culture system, but it may be optimal in another culture system. In this study, we designed 2 groups of experiments and tried to verify our hypothesis. The first group includes culture systems consisting of defined, xeno-free protein matrices and defined, xeno-free media. The second group includes culture systems containing PDL matrix and defined, xeno-free media supplemented with ROCK inhibitor, Y27632. In addition, we also used undefined, xenogeneic Matrigel/mTeSR1 and PDL/mTeSR1 culture systems as the controls for group 1 and group 2, respectively. hPSCs were transferred into each combination of matrix and medium and the optimal systems were identified for supporting hPSC growth. The results confirmed our speculation that synergy between medium, matrix, and exogenous factors is critical for hPSC adhesion and proliferation. Most importantly, from comparative experiments, we found that Vitronectin/TeSR2 in group 1, and PDL/HEScGRO (Y27632) in group 2 were optimal for maintaining the long-term culture and pluripotency of 3 hESC lines and 2 hiPSC lines in defined, xeno-free conditions.
Materials and Methods
Culture systems for hPSCs
Two groups of culture systems were used to culture hPSCs in defined, xeno-free conditions (see Table 1 for abbreviations). The first group included culture systems consisting of defined, xeno-free protein matrices and media (PM/M). Human plasma fibronectin (FN) purified protein (Millipore), Vitronectin (VN, isolated from human plasma; Chemicon), Coating matrix kit (CM, recombinant human type-1 collagen; Invitrogen), and CELLstart™ humanized substrate (CS; Invitrogen) (Fig. 1a) were used to coat culture dishes according to manufacturer's instructions. Defined, xeno-free media used in this study include (1) HEScGRO basal medium (Millipore) supplemented with 100 ng/mL basic fibroblast growth factor (bFGF) (human animal-free recombinant; Millipore) (HGM); (2) xeno-free knockout serum replacer medium (XSR) consisting of KO-Dulbecco's modified Eagle's medium (DMEM) (KO-D) or DMEM/F12 (D-F12) supplemented with 20% XSR (Invitrogen), 1.0% nonessential amino acids, 2 mM GlutaMax, 0.1 mM β-mercaptoethanol, 25 units 25 μg/mL penicillin–streptomycin, and 100 ng/mL bFGF, with or without xeno
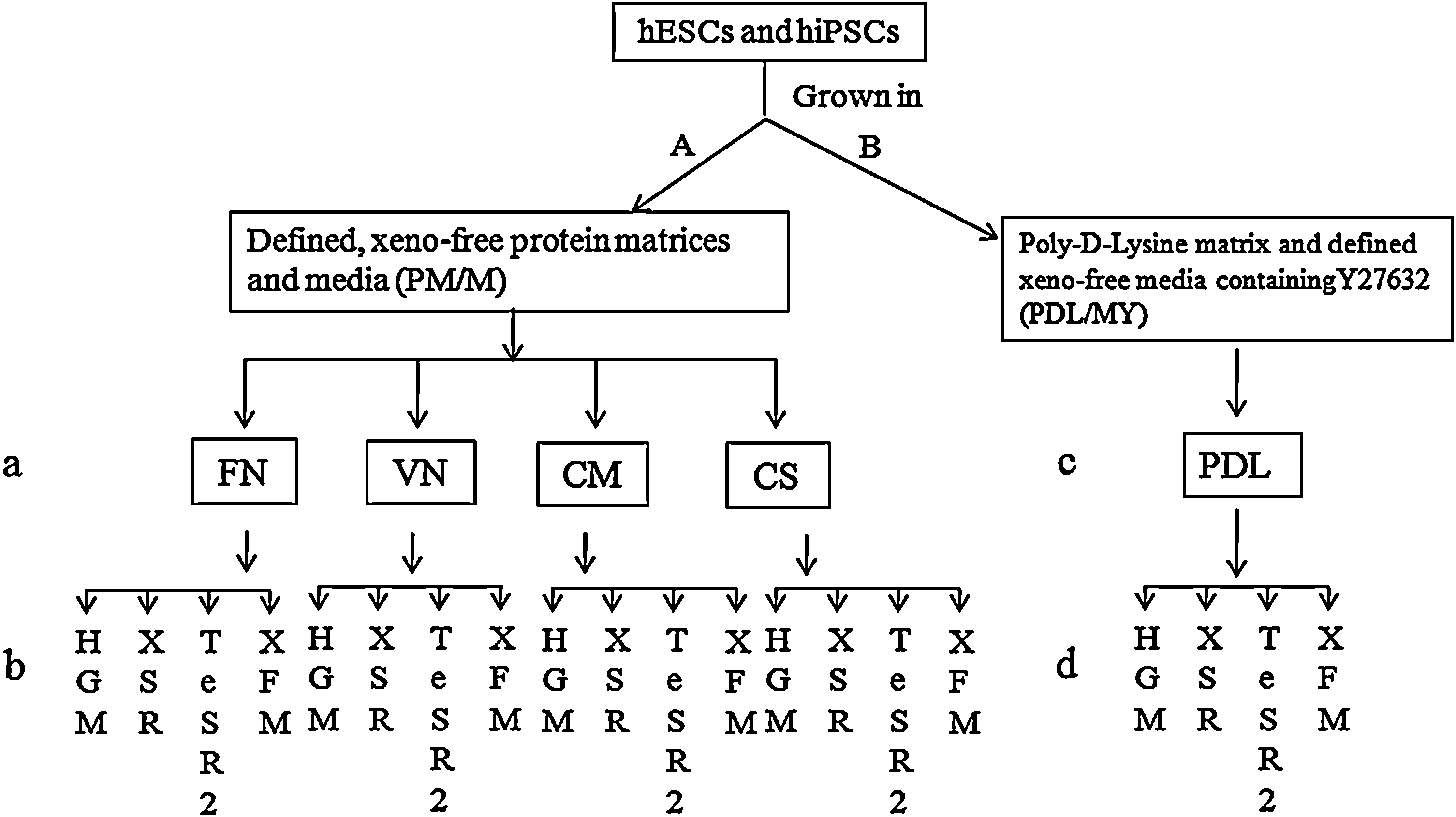
Experimental outline. Group 1
HESC, human embryonic stem cell; DMEM, Dulbecco's modified Eagle's medium; PDL, poly
The second group included culture systems consisting of PDL (Fig. 1c) and defined xeno-free media supplemented with Y27632 (P/MY). The previously mentioned 4 media supplemented with Y27632 (PeproTech) were used to culture hPSCs (Fig. 1d). PDL/mTeSR1 (P/m) culture system was used as the control. In this group, 1 mg/mL Accutase (xeno-free; Stemcell technologies) was used to dissociate hPSC colonies into single cells for passaging and freezing.
For more information about the culture media used in this study, we made a list of their ingredients as much as possible (Supplementary Table S1; Supplementary Data are available online at
Culture of hPSCs
For protein matrices, 35-mm culture dishes are coated respectively with Matrigel, FN, VN, CM, and CS substrates for about 2 h at room temperature in tissue culture hood. The culture dishes are ready to use after aspirating solution completely. For PDL matrix, 35-mm culture dishes are treated with diluted PDL for at least 5 h or overnight at room temperature in culture hood and ready to use after being washed 2 times with D-F12. These PDL-coated dishes can also be stored at 4°C for later use.
hESC lines—H1, H9 (WiCell), and CA1 (Nagy lab, University of Toronto)—and hiPSC lines—BJ-EOS, 4YA, 4YE, and 4YF (James Ellis Laboratory; Ontario Human iPS Cell Facility, Hospital for Sick Children)—used in this study have normal karyotypes (H1 and CA1: 46, XY; H9: 46, XX; 4YA, 4YE, and 4YF: 46, XY) and routinely cultured in Matrigel/mTeSR1 system in our lab.
To test different defined, xeno-free culture combinations of matrices and media in group 1, hESCs and hiPSCs grown in the M/m culture system were dissociated into cell clumps with dispase and transferred to previously mentioned different culture systems. Several days later, the hESCs and hiPSCs grown in different xeno-free culture systems were dissociated into cell clumps and transferred to fresh culture dishes in the same conditions. Continuous culture was performed in this way. For testing the adhesion and viability of hESCs, about 50 dispase dissociated cell clumps were selected and plated in different culture conditions. On the second day (day 1) after plating, the attached cell clumps were counted under inverted microscope.
To test different defined, xeno-free culture combinations of PDL- and Y27632-containing media in group 2, hESCs and hiPSCs grown in the M/m culture system were dissociated into single cells with Accutase and transferred to different culture systems. Several days later, the hESCs and hiPSCs grown in different PDL culture systems were dissociated into single cells and transferred to fresh culture dishes in same conditions. Continuous culture would be performed like this. In this group, we also tested the concentrations of Y27632 and bFGF on hPSC adhesion and proliferation, respectively. In this culture group, we first used the hESC line, H9, along with HGM medium to test the appropriate concentration of Y27632 and bFGF. On the second day morning after plating dissociated single cells, the dead cells were counted and viability was calculated based on this data. The dead cells include 2 parts: cells floating in the medium and mechanically attached cells to PDL. After gently blowing air across the culture surface, the attached dead cells would detach, while small live cell colonies remained attached on the culture surface. The old medium was collected, replaced with fresh medium, and the dead cells were counted.
For XSR medium in both groups, we compared the different effects of 2 basal media, KO-D and D-F12, on hPSC adhesion and viability, respectively.
Determination of cell number
Cell number was determined as described previously [25]. Briefly, hPSC colonies were dissociated into single-cell suspension using 0.05% trypsin/0.53 mM ethylenediaminetetraacetic acid solution. The cells were counted using a hemocytometer and an inverted microscope. The fold expansion for each passage (4–8 passages) was calculated as the cell number at the time of harvest divided by the starting cell number. Statistical analysis on cell expansion was carried out using unpaired Student's t-test for the comparison of 2 groups with a minimal significance of P<0.05.
Characterization of hPSCs grown in different culture systems
Immunocytochemistry was used to examine the expression of pluripotency markers using antibodies: OCT4 (sc5279 Santa Cruz), SSEA-4, TRA-1-60, TRA-1-81, and Nanog (Invitrogen). hPSC colonies were fixed in 4% paraformaldehyde for 15 min at room temperature, and then permeabilized with 0.1% Triton-X 100 in phosphate-buffered saline (PBS) for 15 min at room temperature. The samples were then washed 3 times with PBS and incubated with PBS containing 10% normal goat serum for 30 min at room temperature. Primary antibody was applied at a 1:100 dilution at 4°C overnight. After washing 3 times with PBS, cells were incubated with the appropriate secondary antibodies (Alexa Fluor 546 goat anti-mouse and Alexa Fluor 594 donkey anti-rabbit, 1:200; Invitrogen) for about 2 h at room temperature.
Karyotype analyses were carried out using the G-banding method. Briefly, cells were incubated with 0.1 μg/mL of colcemid at 37°C for 1 h, then trypsinized, resuspended, and incubated in 68 mM HCl for 20 min at room temperature. The samples were then fixed with 3:1 methanol:glacial acetic acid and dropped onto prepared slides to make the chromosome spreads. The dried slides were baked for 90 min at 80°C, treated with 0.05% trypsin for 30 s to 1 min, and then stained with Giemsa and Leishman's solution. At least 10 metaphases spreads were analyzed for hPSCs grown in each culture condition.
In vitro and in vivo differentiation was also carried out in order to investigate the pluripotency of hPSCs grown in optimal culture conditions. hPSCs were spontaneously differentiated as embryoid bodies (EBs) and the resulting cell types were analyzed by marker immunofluorescence using monoclonal antibodies against β-tubulin III (Sigma T8660; 1:400) (ectoderm), smooth muscle actin (Sigma A5228; 1:200) (mesoderm), and α-fetoprotein (Sigma A8452; AFP, 1:400) (endoderm). Antibody localization was detected using Alexa Fluor 546 goat anti-mouse (1:200; Invitrogen). For in vivo differentiation, hPSC colonies were dissociated into cell clumps, centrifuged, and resuspended in culture medium without bFGF. These clumps were injected into the rear leg of 6- to 8-week-old severe combined immunodeficient (SCID) mice (106 cells/injection). Mice were sacrificed at 10–12 weeks postinjection, and the resulting teratomas were fixed with 4% paraformaldehyde and embedded in paraffin. The paraffin blocks were sectioned at a thickness of 5 μm, stained with hematoxylin and eosin, and observed under a brightfield microscope.
Flow cytometry
The expression of pluripotency markers, Nanog, Oct4, and Sox2, of hPSCs grown in several optimal culture conditions was also analyzed by fluorescence-activated cell sorting (FACS). hPSCs were dissociated into single cells and subjected to FACS using a FACS Calibur instrument and the CellQuest software from Becton Dickinson. Ten-thousand events were registered per sample, and analysis of whole cells was performed using appropriate scatter gates to avoid cellular debris and aggregates. The cells were stained using the following antibodies: Anti-Oct4, Alexa Fluor 488 conjugate, Anti-Nanog, N-terminus-Alexa Fluor 488 conjugated, and Anti-SOX-2, FITC conjugate (all from Millipore). Mouse IgG1 Alexa Fluor 488, rabbit IgG Alexa Fluor 488, and mouse IgG2b FITC were used as isotope control depending on the primary antibody isotype.
Results
Synergistic effect of protein matrix, medium, and exogenous factors on hPSC adhesion and proliferation in the PM/M group
hESC lines, H1 and H9, were used to test the optimal culture systems in group 1 (PM/M). When hESCs were cultured in XFM on different matrix protein–coated dishes (FN, CS, CM, and VN), they displayed different colony morphologies and growth rates in different culture conditions. Grown on VN, hESCs displayed a small and round morphology accompanied by a high nucleus-to-cytoplasm ratio, a notable presence of 1–3 nucleoli, and a typical spacing between cells. hESC colonies were elliptical or circular in shape, and formed a distinctly smooth border between hESC colonies and the coated matrix (Fig. 2a). Compared with hESCs cultured in M/m system, hESCs grown in VN/XFM had a slower growth rate. Cultured on other 3 matrices, hESCs showed 2 different colony morphologies without remarkable differentiation. One type of colony was similar to colonies formed in M/m where the cells displayed a small and round morphology as previously mentioned (Fig. 2b1–d1). Another type of colony showed a network structure with cavities and spindle cells inside (Fig. 2b2–d2). Over time, more and more cells inside this kind of colony detached and died. As a large proportion of these colonies (40%–70%) existed in every passage, hESCs grew slowly and appeared in bad condition.

hESCs grown in each combination of protein matrix and medium under defined, xeno-free conditions.
When hESCs grown in XSR (without GFC) on protein matrix–coated dishes (VN, FN, CS, and CM), almost all of the cell clumps attached on VN-coated surface, part of the cell clumps loosely attached to FN-coated surface, and few or no cell clumps attached on CS- or CM-coated surface. Most of the hESC colonies grown on these substrates showed the above-mentioned network structures (Fig. 2e–g), but no cell colony formed on CM (Fig. 2h). Small colonies that formed 1–2 days after plating usually contained several cavities. Over time, most of the cells died and colonies were split into many loose parts. Under these culture conditions, hESCs grew very slowly and also appeared in bad condition.
When GFC was added into XSR medium according to manufacturer's instruction, there is no significant improvement in hESC adherence ability; however, the cell and colony morphologies were improved greatly and displayed typical hESC and colony morphologies (Fig. 2i–k) except on CM (Fig. 2l). While bFGF, even at high concentrations, could not support hESC growth, the addition of unknown factors included in GFC together with bFGF could maintain normal growth. Grown in VN/XSR (with GFC), more than 90% colonies grew healthily and showed typical cell and colony morphology (Fig. 2i). However, compared with M/m, hESCs grown in this culture condition still had a slower growth rate, with bigger cells and thinner colonies. The other 3 culture systems could only maintain partial (FN and CS) or no (CM) growth of hESCs, as these 3 culture systems have a common defect that the cells have poor attachment to the culture surface. Our comparative experiments also suggested that hESCs grown in XSR/D-F12/GFC had stronger adherent ability to the culture matrix than the cells cultured in XSR/KO-D/GFC.
Grown in HGM on VN-coated dishes, hESCs had normal attachment and proliferation only in passage one (Fig. 2m). From the second passage onward, the cells had very poor attachment to the VN matrix. Grown in FN-, CS-, and CM-coated dishes, respectively, hESCs failed to adhere to the matrices (Fig. 2n–p).
After transferring hESCs into TeSR2 in dishes coated with FN, VN, CS, and CM, respectively, the performance of hESCs were as follows: almost all cell clumps attached well and had high proliferative capacity on VN-coated dishes, which were close in appearance to hESCs grown on M/m. hESCs grown in VN/TeSR2 displayed typical hESCs with colony morphologies similar to those grown on MEF feeder cells. However, the colonies were thicker than those grown in M/m (Fig. 2q; Supplementary Fig. S1). Almost all of the hESC clumps failed to attach to FN-, CS-, and CM
Adhesion ability: +(<25%), ++ (25%–50%), +++ (50%–75%), and ++++ (>75%) compared with hESCs (H9 ++++) grown in M/m. Growth rate: + (<25%), ++ (25%–50%), +++ (50%–75%), and ++++ (>75%) compared with hESCs (H9 ++++) grown in M/m.
hESC, human embryonic stem cell; n/a, not available.
The hiPSC lines, 4YA and 4YF, were used to test these culture systems in group 1. Similar as hESCs, hiPSCs grew normally in VN/(D-F12/XSR/GFC) and VN/XFM, and grew well in VN/TeSR2 (Supplementary Fig. S2; Table 3) for more than 10 passages, respectively.
The data on adhesion ability are the mean value of 3 repeat experiments, and expansion fold of cells is the mean value of 5 passages. The data were expressed as mean±SD. The average passage time and split ratio of cells cultured in VN/TeSR2 is 4 days and 1:6. Different from the above, the average passage time and split ratio of the cells grown in both VN/(D-F12/XSR/GFC) and VN/XFM are 5 days and 1:4.
SD, standard deviation.
From these results, we concluded that medium, protein matrix, and exogenous factors played a synergistic role in supporting undifferentiated hPSC growth. The TeSR2/VN system was the best culture system, with VN/(D-F12/XSR/GFC) and VN/XFM being second best culture for hPSC adhesion and proliferation. All other combinations were suboptimal for hPSC culture.
Synergistic effect of PDL matrix, medium, and exogenous factors on hPSC adhesion and proliferation in P/MY group
First, the hESC line, H9, together with HGM were used to test the appropriate concentration of Y27632 and bFGF in group 2 (P/MY). The comparative experiments indicated that 5–10 μM Y27632 and 100 ng/mL bFGF were optimal concentrations for supporting undifferentiated hESC growth (Fig. 3). hESCs could be maintained on PDL in HGM supplemented with 100 ng/mL bFGF and 10 μM Y27632 for at least 34 passages (Fig. 3g). The concentration of Y27632 has a close relation to hESC morphology. Without addition of Y27632, single hESCs physically attached for a short time, then detached and completely died (Fig. 3a). The minimum concentration of Y27632, which could be used to maintain hESC growth in this culture system, was 2.5 μM. With an increase in Y27632 concentration, hESC colony would become flatter (Fig. 3b–e). In a certain range of Y27632 concentration (from 2.5–5 μM), hESC colonies grown in PDL/HGM were similar to mouse embryonic stem cell (mESC) colonies grown on MEF feeder layers (Fig. 3b, c). Cultured in medium supplemented with high concentrations of Y27632 (>10 μM), hESC formed flatter colonies and resembled mESC colonies grown on feeder-free conditions (Fig. 3d, e). The concentration of Y27632 also had a great impact on hESC attachment to the culture surface. When the concentration of Y27632 was less than 5 μM, hESC colonies attached loosely and some colonies were easy to detach from the culture surface when refeeding. When the concentration of Y27632 was more than 15 μM, the cell colonies tightly attached and would take a long time to dissociate them into single cells. To determine whether the concentrations of Y27632 had an effect on the pluripotency of hESCs, alkaline phosphatase staining (Supplementary Fig. S3a, c) along with immunocytochemistry staining for Oct4 (Supplementary Fig. S3b, d) and FACS (Supplementary Fig. S3e, f) were performed on hESCs cultured in low (2.5 μM) and high (10 μM) concentrations, respectively. The results indicated that the concentrations of Y27632 had no effect on the pluripotency of hESCs. Surprisingly, almost all cells died in 1 passage when cultured on PDL in HGM supplemented with Y27632, without addition of bFGF (Supplementary Fig. S4). hESCs would differentiate at passages 2–4 in 20 ng bFGF/mL (Fig. 3h, j) and at passages 9–11 in 60 ng bFGF/mL (Fig. 3i, k). The cell density used in these experiments was approximately 3×104/cm2. When starting to differentiate, the cells almost stopped growing, separated from the colonies, and became Oct4 negative (Fig. 3j, k).

Test of appropriate concentration of bFGF and Y27632 in group 2 (PDL/MY) for hESC culture.
From single cells the process of colony formation consisted of 4 steps: attachment of individual cells, migration, reaggregation, and proliferation (Fig. 4a–d). In PDL/HGM, the proliferation rate of H9 cells was comparable to cells cultured in M/m (Tables 4 and 5). When grown in medium containing 15 μM Y27632, hESC lines, H1 and CA1, showed similar growth rate (Table 5) and cell and colony morphology (Supplementary Fig. S5) in this culture system. When grown in different concentrations of Y27632 (2.5–15 μM), the second day after single-cell plating (Day 1), the percentage of attached and viable cells can be calculated. From this data, we know that there is no obvious difference in their adhesion capacity among the different cell lines in each concentration. With the increase of Y27632 concentration, the attachment ability increased (Supplementary Table S2). However, along with the growth of hESC colonies, the difference in attachment ability began to appear in different cell lines cultured in each concentration of Y27632 from the third day (Day 2). Different from H9 cells, H1 and CA1 hESCs formed compacted dome-shaped colonies with poor attachment capacity in 7.5 μM Y27632 (Supplementary Fig. S5a, c). Normally, when the concentration of Y27632 was less than the optimal concentration, hESCs attached loosely and part of colonies were easy to detach from the culture surface when refeeding cells from day 2. It is hard to estimate the detached colony number, which depended on careful manipulation. HGM containing 15 μM Y27632 was suitable for cell attachment and proliferation of both hESC lines (Supplementary Fig. S5b, d). This result suggested the different needs of Y27632 concentration for supporting different hESC lines, and also revealed differences among the different cell lines.

Formation of hESC colonies at high and low cell density.
Adhesion ability: + (<25%), ++ (25%–50%), +++ (50%–75%), and ++++ (>75%) compared with hESCs (H9 ++++) grown in M/m. Growth rate: + (<25%), ++ (25%–50%), +++ (50%–75%), and ++++ (>75%) compared with hESCs (H9 ++++) grown in M/m.
The cell expansion is the mean value of 5 passages. The data were expressed as mean±SD. Statistical analysis of the data indicated no significant difference (t-test, P<0.05) between PDL/HEScGRO (Y) culture and the control culture in cell expansion fold of H1, H9, and CA1, respectively. The different hESC lines have the similar passage time (average 3.5 days) and split ratio (1:8). No significant difference (t-test, P<0.05) was observed between PDL/HEScGRO (Y) culture and the control culture in cell expansion fold of 4YE and 4YF, respectively. Both of the human induced pluripotent stem cell lines have the similar passage time (4 days) and split ratio (1:6).
H1 and H9 cells were also used to test other defined, xeno-free, Y27632-containing media, such as mTeSR1 (Control, containing xenogeneic BSA), TeSR2, XFM, and XSR (KO-D, or D-F12 with or without GFC), in PDL-coated dishes. Cultured in mTeSR1 medium at low density (1×104 cells/cm2), hESCs attached, searched each other, formed small aggregates, and grew normally in small colonies. However, as the colonies became larger and larger, a lot of the cells died and detached. When cells were seeded in normal density (3×104 cells/cm2), the colonies grew normally in first 2 days with less cell death. However, from day 3, more and more cells began to die, and the colonies became smaller and smaller as time went on (Fig. 5b, i, p). Cultured in XSR medium consisting of KO-D or D-F12 supplemented with XSR, bFGF, and Y27632 (KO-D/XSR or D-F12/XSR), cells attached loosely and did not proliferate even at high concentration (20 μM) of Y27632, and almost all cells died on the second day (Fig. 5c, j, q).
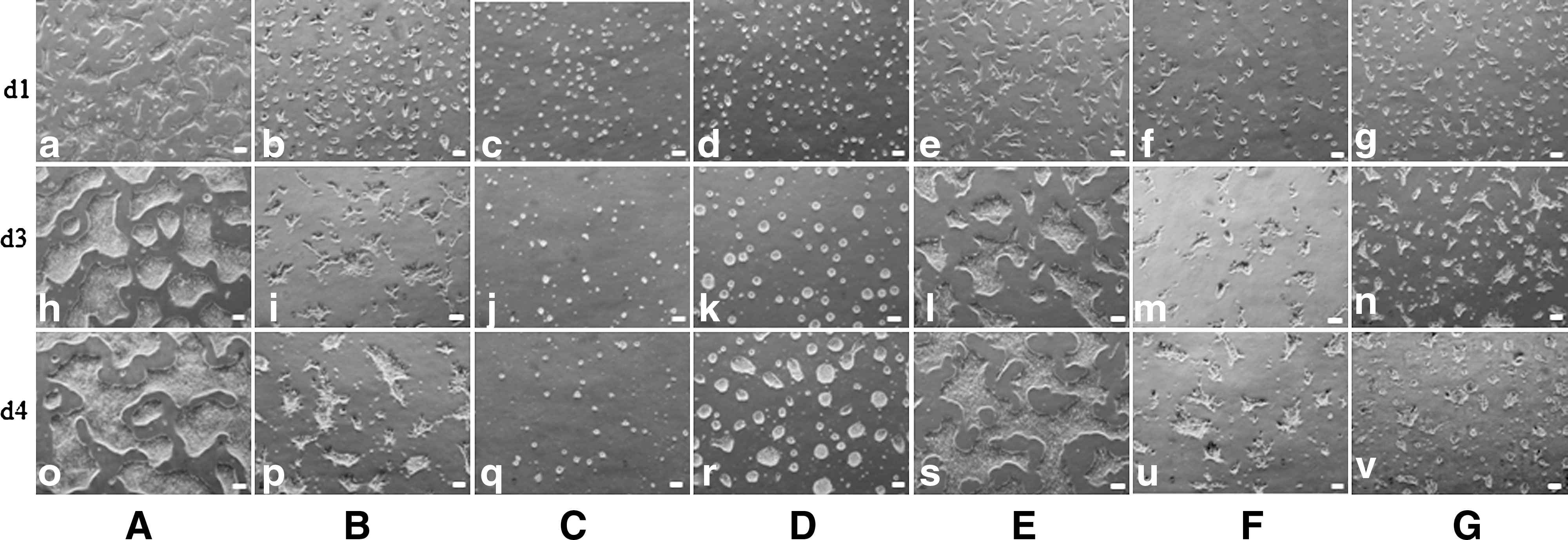
hESCs cultured in different systems in group 2.
Interestingly, with the addition of GFC, both media could support hESC proliferation, but the cells had stronger adherence capacity in D-F12/XSR/GFC than in KO-D/XSR/GFC (Fig. 5d, k, r and 5e, l, s). Cultured in XFM, cells had the similar appearance with those cultured in mTeSR1 (Fig. 5f, m, u). Grown in TeSR2 medium, cells attached well, but showed poor proliferation, most of them dying 2–3 days later (Fig. 5g, n, v). All of the above experiments were repeated at least 3 times. The results indicated that HGM supplemented with Y27632 can support hESC growth well (Fig. 5a, h, o). Long-term xeno-free expansion in PDL/HGM system was found to maintain hESC properties, including morphology, pluripotency, stable karyotype, and expression of hESC-specific markers, for more than 20 passages. The results of H9 grown in the different culture systems in group 2 (P/MY) are summarized in Table 4.
hiPSCs, 4YA, 4YE, and 4YF, were used to test these culture systems in group 2 (P/MY). Compared with hESCs, 4YE and 4YF cells can also grew well in PDL/HGM culture system for long-term culture (Supplementary Fig. S6a, b; Table 5), but with slower growth rates. However, 4YA cells started to differentiate at passages 3–5 and completely differentiated at passages 5–7 (Supplementary Fig. S6c). HGM supplemented with GFC could improve the adherence and proliferation of 4YE and 4YF cells, but could not prevent the differentiation of 4YA cells (data not shown). Comparing the proliferation of different hESC and hiPSC lines cultured in P/MY with conventional culture methods (M/m) indicated no significant difference (t-test, P<0.05) between PDL/HEScGRO (Y) culture and the control culture in cell expansion fold of H1, H9, CA1, 4YE, and 4YF, respectively. Naturally, differences in the proliferation rates existed among different cell lines (Table 5).
hiPSCs grown in both KO-D/XSR and D-F12/XSR medium without GFC attached loosely and did not proliferate even at high concentrations (20 μM) of Y27632. After addition of GFC, KO-D/XSR medium still did not support hiPSC adherence and proliferation. Interestingly, with the addition of GFC, D-F12/XSR medium could support hiPSC growth well, even though their adherence was still not comparable to hESCs cultured in the same medium (data not shown). From these experiments, we concluded that (1) synergy between the basal medium, XSR, PDL bFGF, Y27632, and GFC is critical for maintenance of hESC and hiPSC growth; (2) in the presence of XSR, bFGF, Y27632, and GFC, D-F12 has a stronger ability to improve hESC or hiPSC attachment to culture matrix; (3) in addition to bFGF, other growth factors present in GFC are necessary for hESC or hiPSC adherence and proliferation in this culture system; and (4) the main role of Y27632 is to improve the attachment of single cells to culture surface. However, in some culture media, such as KO-D/XSR (without GFC) medium containing Y27632, hESCs or hiPSCs do not attach or only attach loosely. This means that the cooperation between Y27632 and other factor(s) is necessary to keep cells attached to each other or to the culture matrix. However, as the formulation of GFC is proprietary, the company is unable to release detailed information on it. We only know that it is composed of human and recombinant proteins.
Characterization of hPSCs grown in different defined, xeno-free culture systems
H9 hESCs grown in both VN/TeSR2 and PDL/HGM culture systems and 4YF hiPSCs grown in PDL/HGM culture system were characterized completely according to the criteria for hESCs. These criteria include the expression of pluripotency markers of hESCs, karyotype stability, and 3 germ layer differentiation in vitro (embryonic bodies, EBs) and in vivo (teratoma formation). Grown in VN/TeSR2 culture system, hESCs were stained positive for the expression of pluripotency markers, Oct4, Nanog, SSEA-4, TRA-1-60, and TRA-1-81, using immunofluorescence (Fig. 6a–e). They differentiated into cell types derived from all 3 germ layers in vitro (Fig. 6f–h) and in vivo (Fig. 6i–k), and exhibited normal and stable karyotype (Fig. 6E). The FACS results indicated that the cells expressed very strong positive for Oct4, Nanog, and Sox2 (Fig. 6l–n). As for hESCs (H9) and hiPSCs (4YF) grown in PDL/HGM system, the previously mentioned items were performed on them and similar results were obtained. Also, the expression of alkaline phosphatase of both cell lines was also assayed, which was intensely positive throughout each colony (Figs. 7 and 8).
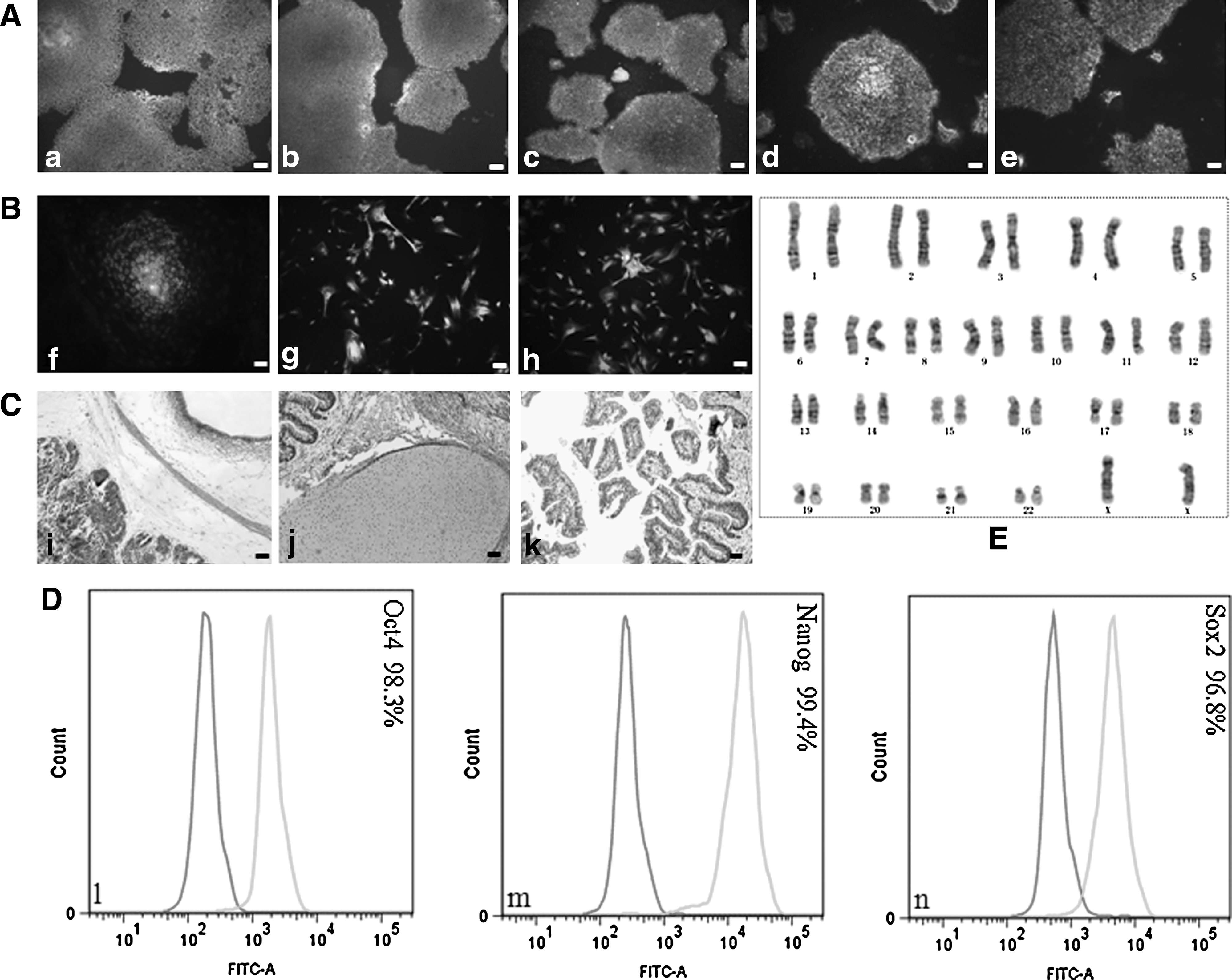
Characterization of hESCs (H9) grown in VN/TeSR2 culture system.
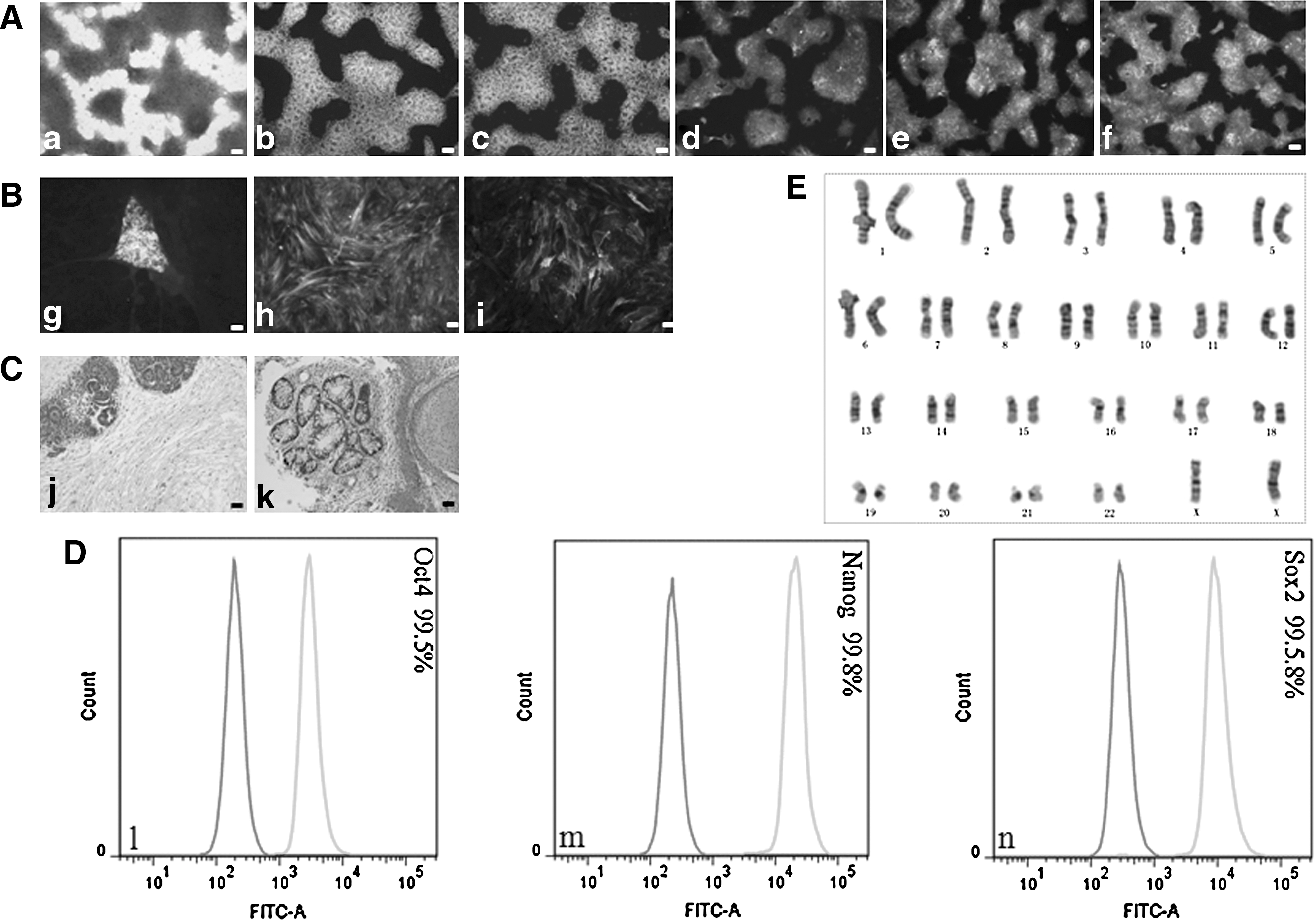
Characterization of hESCs (H9) grown in PDL/HGM culture system.
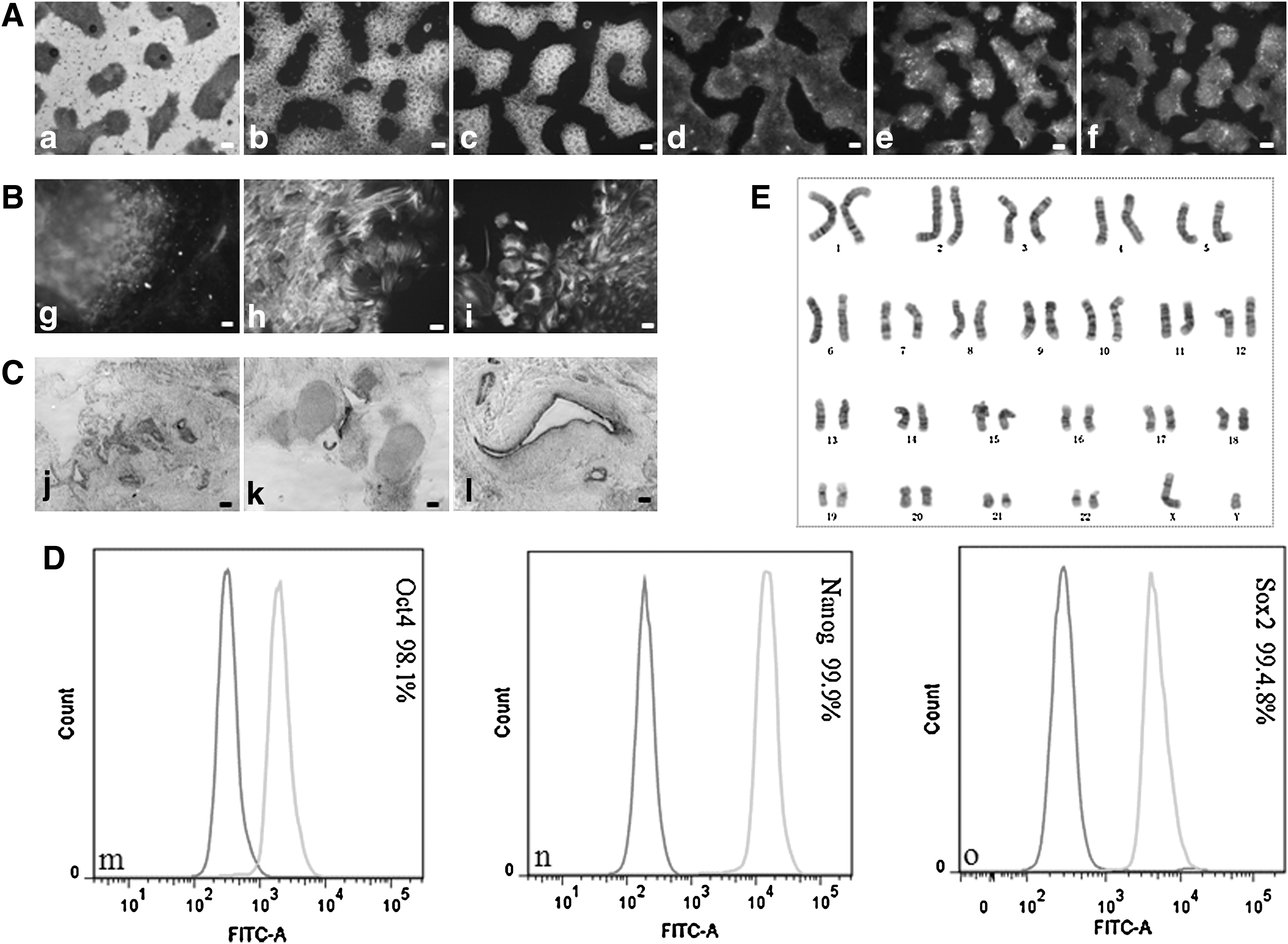
Characterization of hiPSCs (4YF) grown in PDL/HGM culture system.
Discussion
At present, establishment of xeno-free, defined culture conditions for hPSC derivation and expansion is one of the most challenges for the potential application of hPSCs in regenerative medicine and cell therapy. For this purpose, some progress has been made in this area in recent years. Several research groups reported that some kinds of xeno-free, defined media could effectively support hPSC growth in conjunction with human feeders or purified proteins from human tissues, recombinant proteins, and synthetic matrices [20 –24,26].
Based on previous studies, by comparing 2 groups of culture systems, PM/M and P/MY, we concluded that synergistic effect of coating matrix, medium, and growth factors played a critical role in supporting undifferentiated hPSC attachment and proliferation. Our previous data demonstrated that mTeSR1 and TeSR2 could support hESC and hiPSC growth in very good condition on hESC
Our results demonstrated that the VN/TeSR2 culture system in group 1 and the PDL/HGM culture system in group 2 can support hESC and hiPSC adhesion and proliferation. However, VN is very expensive and the higher concentration of it (2 μg/cm2) is required to coat the culture dishes. In the contrary, ROCK inhibitor, Y27632, is much cheaper. Moreover, hPSCs grown in this system are passaged in clumps, which hinder the application of hPSCs in many aspects, such as FACS, electroporation, and clonal selection [30,31]. Compared with the VN/TeSR2 culture system, PDL/HEScGRO(Y) culture system can overcome these weaknesses as Y27632 supports single-cell passaging and is economical to use. In addition, hESCs and hiPSCs grow faster in PDL/HEScGRO(Y) culture system than in VN/TeSR2 culture system. Previous studies have indicated that Y27632 is safe and efficient at supporting hPSC expansion and greatly improves hPSC adherence and viability after cryopreservation [32 –35]. In the P/MY culture group, Y27632 played a critical role for hPSC attachment and viability.
The process of colony formation consists of 4 steps: attachment of individual cells, migration, reaggregation, and proliferation. If hPSCs were seeded into the PDL/HGM culture system without addition of Y27632, they physically attached to the PDL substrate, but failed to migrate toward each other and form cell aggregates. Without cell–cell contact, attached cells would detach from culture substrate and die. When hPSCs were grown in PDL/HGM culture system at low cell density (2000 cells/cm2), only a few colonies were observed in each dish; when grown at clonal density (<1000 cells/cm2), no cell colonies were observed several days later after plating (Fig. 4B). This means that it is hard for cells to migrate toward each other and form aggregates, as the distance between them is too far to contact and aggregate. We speculated that cell–cell contact and not antiapoptosis is predominantly responsible for the hPSC growth in this PDL/HGM culture system.
In conventional culture, as Y27632 can greatly promote hPSC attachment and viability but not proliferation, Y27632-containing medium usually will be replaced with fresh medium without Y27632 the second day after seeding or thawing dissociated cells [14,36]. However, in this culture system, Y27632 must be in the culture medium all the time, because hPSC colonies will detach from the culture surface quickly if refeeding proceeds without Y27632.
Surprisingly, when hESCs were cultured in PDL/HGM without the addition of bFGF, all of the cells died almost immediately in short time, which is different from other culture systems. Grown in other feeder-dependent or feeder-free culture systems, hPSCs will differentiate, but not die without addition of bFGF. We conclude that PDL matrix, basal HEScGRO medium, bFGF, and Y27632 together provide an adequate supply of nourishment and signaling for supporting undifferentiated hPSC growth and self-renewal.
Unexpectedly, 1 of 3 hiPSC lines used in this study, 4YA, could not be maintained more than 5 passages in an undifferentiated state. This line was derived by via the lentiviral transfection of the 4 Yamanaka reprogramming factors [37]. Position effects caused by the random insertion of these genes probably is a major cause of hiPSC diversity. Interestingly in other research we have observed that this line, 4YA, unlike hESCs gives rise to cyst-like structures when induced to differentiate toward that cartilage lineage and continues to express c-Myc during differentiation (unpublished data). Compared with defined culture systems, it is likely that undefined systems can provide many unknown factors, which can effectively support the majority of hiPSC lines. However, as there is a diversity of the hiPSC lines, even if grown in undefined culture conditions, different hiPSC lines will have different growth status.
Normally, in undefined culture systems, 10 μM Y27632 is added into media for improving the survival of single cells after thawing and passaging of hPSCs [32 –34,38]. In this study, the concentration of Y27632 used in the defined PDL/HGM culture system depended on hPSC line's attachment capacity. We observed that 5–10 μM Y27632 was appropriate for maintaining H9, while 15 μM was optimal for all other hPSC lines tested. Therefore, it is necessary to determine the optimum concentration of Y27632 for each cell line cultured in this system.
The results suggest that it is not every pair of medium and matrix made or derived for hPSCs, which can support hPSC adherence and proliferation. The combination of medium and matrix must meet the requirement for signaling pathways to maintain hPSC attachment and growth in an undifferentiated state. The results also highlight 2 culture systems for supporting hPSCs under defined, xeno-free conditions. Despite the ability of these xeno-free conditions to support hPSC maintenance, the utility of our systems for deriving hESC or hiPSC lines has not yet been tested. One of our next tasks is to derive new cell lines under these defined, xeno-free conditions.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.