
Abstract
The rodent pituitary gland undergoes prominent maturation during the first weeks after birth, including a well-known increase in hormone-producing cells. In the past, it has frequently been postulated that stem cells are involved in this early-postnatal growth phase. This hypothesis can now be explored, as pituitary stem/progenitor cells were recently identified. Here, we analyzed in detail the mouse pituitary stem/progenitor cell compartment during the first postnatal week and compared its phenotype with that at the end of the first pituitary growth wave and at adult age. Stem/progenitor cells, as assessed by both side population phenotype and Sox2 expression, are most abundant at birth and gradually decline toward adulthood. The neonatal stem/progenitor cell compartment is clearly more active in terms of proliferation, stemness gene expression, and stem cell-related functional activity including sphere formation and multipotent differentiation capacity. In situ examination of pituitary sections reveals peculiar topographical arrangements of Sox2+ cells, again more pronounced at the neonatal age. Sox2+ cells are particularly prominent at the wedge junction of the anterior and intermediate lobe, and clusters of Sox2+ cells appear to sprout from this and other cleft-lining, marginal zone regions. Colocalization of Sox2 and hormones is generally not observed, thus suggesting mutually exclusive expression. Together, the neonatal pituitary stem/progenitor cell compartment displays an activated phenotype, thus supporting its involvement in the early-postnatal maturation process of the gland.
Introduction
T
In the present study, we started to explore the stem/progenitor cell compartment during the vivid maturation process of the murine gland in the first week after birth, and compared this neonatal phenotype with that at the end of the first growth phase (3 weeks after birth) and at adult age. The stem/progenitor cell compartment is clearly more active in the neonatal versus the mature pituitary in terms of abundance and proliferative activity, topographical organization aspects, and in vitro differentiation capacity.
Materials and Methods
Animals
Mice (FVB strain) of different ages (postnatal day 1 [P1] to P7; P21; and 12-week-old) were purchased from Elevage Janvier (BioServices, Uden, The Netherlands), or bred within our own Animal Facility (University of Leuven/K.U.Leuven, Belgium). For the analyses at P21 and adult age, female mice were used; for the examination of P1 to P7 ages, random mixtures of male and female pups were used. Preliminary experiments did not show gender differences relevant to our study at these neonatal ages (data not shown).
All animals were kept in the Animal Housing Facility under conditions of constant temperature, humidity, and day-night cycle and had access to food and water ad libitum. Animal manipulations were approved by the K.U.Leuven Ethical Committee and performed in accordance with the ethical guidelines.
Dissociation of AP cells
Mice were anesthetized with CO2 and killed by decapitation. The neurointermediate lobe of the pituitary was discarded using a stereomicroscope, and the AP was isolated and dissociated into individual cells with trypsin as described earlier [10]. Cells were resuspended in pituitary-optimized cell culture medium (serum-free defined medium [SFDM]; Invitrogen, Grand Island, NY; [10]) and were quantified using a Z2 Coulter Counter (Analis, Suarlée, Belgium).
Flow-cytometric analysis of AP cells
AP cells were incubated with Hoechst 33342 (Sigma-Aldrich, Bornem, Belgium) at a final concentration of 2.5 μg/mL for 90 min, subsequently stained with phycoerythrin (PE)-conjugated rat anti-mouse Ly6A/stem cell antigen 1 (Sca1) antibody (BD Biosciences, San Jose, CA), and finally examined by dual-wavelength flow cytometry (FACSVantage; BD Biosciences) for analysis and cell sorting, all as previously described in detail [10,11]. The Hoechst-effluxing SP was divided into a subset of cells displaying high Sca1 expression (Sca1high SP) and a nonSca1high subpopulation, which we recently demonstrated to enrich for the pituitary stem/progenitor cells [11], for simplicity further referred to as the “stem cell”-SP (SC-SP) fraction. The SP phenotype was confirmed with verapamil (Sigma-Aldrich), which blocks Hoechst efflux, and the Sca1 staining was verified, and gates positioned, using controls with isotype antibodies (BD Biosciences) (data not shown).
Whole-genome expression profiling
The SC-SP fractions from neonatal (P2, P5) and adult mice were sorted by FACS into a cold lysis solution of the RNeasy Micro Kit (Qiagen, Venlo, The Netherlands) (n=2). Total RNA was extracted immediately after cell collection according to the manufacturer's protocol, and RNA samples were stored at −80°C until further processing in collaboration with the Microarray Facility (MAF-VIB, K.U.Leuven) as previously described [11]. Briefly, RNA quality and concentration were determined using Agilent Picochips on an Agilent BioAnalyzer 2100 (Agilent Technologies, Santa Clara, CA). Two ng of high-quality RNA was amplified using the NuGen Pico WTA kit (NuGen Technologies, San Carlos, CA). Cy3 label was incorporated into cRNA probes that were then hybridized onto Agilent whole mouse genome oligonucleotide arrays (Ag-Mo-WG-G4122F). Slides were scanned with an Agilent DNA microarray scanner, and data were processed using the Agilent's Feature Extraction Software. Genes were considered differentially expressed between neonatal and adult SC-SP when at least 1.5-fold different with P<0.05.
Pituisphere assay
Pituitary stem/progenitor cells generate spheres, a hallmark of stem/progenitor cells of multiple tissues [10 –12]. To study pituisphere-forming capacity, dispersed AP cells were seeded at a density of 100,000 cells/mL in 35-mm nontreated culture dishes (Iwaki, Scitech Division, Chiba, Japan) as described earlier [10,11]. Briefly, cells were cultured in SFDM (2 mL/dish) containing B27 (1:50; Invitrogen) and recombinant human basic fibroblast growth factor (20 ng/mL; R&D Systems, Minneapolis, MN) in a humidified 1.9% CO2 incubator at 37°C. Fresh medium was added after 3 days. Live pictures of the spheres formed were taken using a Nikon Eclipse TS100 microscope (Nikon Instruments, Melville, NY). Spheres were counted under the microscope at day 5 of culture.
Pituisphere-initiating cells display multipotent differentiation capacity similar to the stem cells of other tissues [11,12]. To compare the differentiation competence between neonatal and adult cells, pituispheres from P1 and adult mice were manually picked at day 5 and transferred to glass coverslips coated with growth factor-reduced Matrigel (1:10 in SFDM; BD Biosciences). Pituispheres were either immediately processed for expression analysis of Sox2 or hormones by immunofluorescence (see below), or first cultured in SFDM (without supplements) for either 3 or 7 days.
Immunofluorescence analysis of dissociated AP cells
Freshly dispersed AP cells and FACS-sorted cell populations were deposited on Superfrost Plus glass slides (Menzel, Germany) by cytospin. Samples were fixed with paraformaldehyde (PFA, 4% in PBS; Invitrogen) and processed for immunostaining. Therefore, cells were permeabilized with saponin (0.5% w/v in PBS; Sigma-Aldrich) and aspecific binding sites preadsorbed with 20% v/v normal donkey (Sigma-Aldrich) or goat (Millipore, Brussels, Belgium) serum. Subsequently, primary antibodies (see Supplementary Table S1; Supplementary Data are available online at
Immunofluorescence examination of pituitary vibratome sections and pituispheres
Whole pituitaries of neonatal and mature mice were carefully isolated and fixed in 4% PFA. After embedding in agarose (2% w/v in PBS; Lonza, Rockland, ME), the pituitaries were coronally sectioned to 50-μm slices using a vibratome (Microm HM 650V; Prosan, Merelbeke, Belgium). The slices were further processed essentially as described earlier [10,11,15]. Sections were rinsed with PBS and permeabilized with Triton X-100 (0.4% v/v in PBS; Sigma-Aldrich). Sections were treated with normal goat or donkey serum and subsequently incubated with the appropriate (one or more) primary antibodies (see Supplementary Table S1). After rinsing thoroughly, slices were incubated with the appropriate (one or more) secondary antibodies and nuclei labeled with ToPro3 (1:100; Invitrogen). Finally, sections were mounted in Vectashield on glass slides and covered with glass coverslips. Omission of primary antibodies as negative controls did not result in signals (data not shown).
Pituispheres were fixed with 4% PFA and stained for Sox2 and the pituitary hormones (GH; ACTH or its precursor, pro-opiomelanocortin; PRL; and glycoprotein hormone α-subunit (αGSU), the common subunit of TSH and LH/FSH), either all together or separately, as just described.
Stained vibratome sections and pituispheres were scanned with a confocal laser scanning microscope (LSM 510; Zeiss, Zaventem, Belgium). Z-stacks were analyzed and edited using Zeiss LSM Image Browser software and Microsoft PowerPoint (2007).
Statistical analysis
Analysis of variance and Tukey post hoc analysis were performed using SPSS software (Version 13.0; IBM, Brussels, Belgium). Data were considered statistically significant when P<0.05.
Results
The SC-SP is abundant at birth and declines toward mature age
To characterize the stem/progenitor cell compartment of the postnatally developing pituitary, we first analyzed the SP of the AP from mice at different days after birth (P1 to P7), and compared these data with the age of 3 weeks (P21) when the first pituitary postnatal growth wave is finalized, and to adult age (Fig. 1; see also [10,11,16]). The nonSca1high subset of the SP, recently identified to enrich for pituitary stem/progenitor cells (for simplicity here referred to as the SC-SP), is largest at P1 (75% of the SP), and declines with progressing development to 52% at P7 and further to 45% and 37% in P21 mice and adult animals, respectively (Fig. 1). These results indicate that pituitary stem/progenitor cells are abundant at birth and regress in number with further development, thus suggesting that during the early-postnatal phase, stem/progenitor cells participate in AP maturation and decline in number by giving rise to hormone-producing cells.

Side population (SP) analysis of anterior pituitary (AP) cells from neonatal to mature age.
Sox2+ stem/progenitor cells are more abundant, and their topographical phenotype more pronounced, in neonatal versus mature pituitary
The SC-SP of the adult pituitary shows prominent expression of Sox2 [11]. In recent studies, Sox2+ cells were designated as stem/progenitor cells of the gland [11 –14]. In the pituitary in situ, Sox2+ cells are found in the marginal zone (MZ) that borders the cleft - a remnant of the embryonic pituitary anlage, Rathke's pouch [1]−, and in groups scattered over the anterior lobe's parenchyma [11 –13]. This topography suggests the existence of multiple stem/progenitor cell niches in the gland. Moreover, besides the cells with Sox2 in the nucleus – the expected location for a transcription factor –, cells with Sox2-immunoreactive signal in the cytoplasm are also observed [11], further referred to as “Sox2-cytoplasmic” cells.
To characterize the Sox2+ stem/progenitor cell compartment in the pituitary in situ during the different phases of postnatal development, we performed immunofluorescence analysis in vibratome sections of early-postnatal (P1, P4, P7) pituitaries, and compared the observations with P21 and adult mice [11] (Fig. 2). Here, we focus on 3 particular regions of interest because of striking Sox2 expression (see Fig. 2A):

In situ Sox2 expression in the pituitary of neonatal and mature mice.
i) the MZ around the cleft consisting of the bordering layer and adjacent submarginal strip;
ii) the area where the AP and IL meet, further termed the “wedge” region;
and iii) the anterior lobe's parenchyma.
MZ
During the first postnatal week, Sox2+ cells lining the cleft constitute at many sites a multicellular layer of compact organization, which is most prominent at P1 (Fig. 2B). In contrast, the marginal layer at P21 and adult age is at most places unicellular, and appears less compact (Fig. 2B). Bunches of Sox2+ cells are sometimes observed, thus suggesting budding from the periluminal layer at these locations (Fig. 2B, encircled). These bulges are more expanded and are encountered more frequently in the neonatal than the older pituitary (Fig. 2B). The bulges sometimes develop into strings of Sox2+ cells (Fig. 2B; Supplementary Fig. S1A, arrows), thus suggesting the existence of sprouting, “migratory streams.”
Adjacent to the marginal layer(s), a submarginal strip is present that at many places is devoid of Sox2+ cells (dashed lines in Fig. 2B and Supplementary Fig. S1B). Further distal from the marginal layer, Sox2+ cells are again found and most abundant at P1, further declining in number with proceeding development. In addition, Sox2-cytoplasmic cells appear in the marginal region; they remain very sporadic at P1 but increase in occurrence at more advanced ages (Fig. 2B, arrowheads).
Finally, the periluminal layer at the IL side of the cleft contains clearly more Sox2+ cells in neonatal than in older mice (Fig. 2B; Supplementary Fig. S1B).
Wedge area
In early-postnatal mice, Sox2+ cells at the junction of the AP and IL as well as in the close vicinity of this wedge are most abundant at birth (P1), have somewhat decreased by P4 and P7, and further decline toward adult age (Fig. 2B; Supplementary Movies S1–S5; still images of all Supplementary Movies are provided in Supplementary Fig. S9).
At some places of the wedge, extensions of the cleft into the AP parenchyma are observed (at all ages analyzed). These invaginations are bordered by Sox2+ cells that again are more abundant and more multicellular in neonatal than older mice where the invaginations are predominantly lined by a single Sox2+ cell layer (Supplementary Fig. S1C). At some locations, bifurcation of the invaginations is observed, also more frequently encountered at neonatal than P21 age and almost absent at adult age (Supplementary Fig. S1C). Finally, the wedge region at some locations displays an AP-to-IL transition picture with a portion of the AP folding back over the IL (Supplementary Fig. S1D, boxed; also sketched in Fig. 2A). At this spot, Sox2-cytoplasmic cells can be highly concentrated in P21 and adult mice (Supplementary Fig. S1D).
AP parenchyma
As reported earlier [11 –13], Sox2+ cells are also present within the lobe's parenchyma. Cells with nuclear Sox2 signal are clearly more abundant in neonatal than in adult pituitary (Fig. 2B). Whereas Sox2+ cells look evenly distributed at P1, a picture of groups of cells emerges with progressing development (Fig. 2B, arrows). These groups, consisting of a small number of Sox2+ cells (<10), are scattered over the anterior lobe. In addition, larger co-arrangements are observed in which Sox2+ cells border a lumen. These clusters (Fig. 2C; see further below) are lower in occurrence than the smaller Sox2+ cell groups.
In addition, Sox2-cytoplasmic cells are found in the parenchyma, again only sporadically at P1 and P4 and more prominent at P7, with a further rise in abundance toward adult age (Fig. 2B, arrowheads). At all ages analyzed, the in situ incidence of Sox2-cytoplasmic cells remains lower than that of the Sox2-nuclear cells.
Taken all together, the Sox2+ cells show higher abundance in neonatal than in further developed and mature pituitary. To validate these in situ observations, we quantified the Sox2+ cells in cytospin samples of dissociated AP cells. The proportion of Sox2+ cells decreases 2.5-fold from P1 to adult age, with only 1% of these cells showing a Sox2-cytoplasmic signal at birth as opposed to 10% in adulthood (Fig. 2D). The proportion of total Sox2+ cells is higher than the SC-SP proportion as described earlier. Sox2+ cells are enriched in the SC-SP fraction (Supplementary Fig. S2; see also [11] for adult mice), but are also found in the cell population between the SP and the main population (MP) as gated in the FACS profile (referred to as the “inter-population” or IP; see Supplementary Fig. S2). In addition, in other tissues, different regions can be distinguished in the SP FACS profile, with decreasing capacity for Hoechst efflux corresponding to progressive stages in commitment and differentiation [17,18]. Of further note, the Sox2+ cell proportion in adult mice is higher than the number reported by others (3%–5% in [12]). One reason is that the mouse strain used here (FVB) visibly contains more Sox2+ cells in the AP than the C57Bl6 strain as used by Fauquier et al. [12] (data not shown). Moreover, Sox2-cytoplasmic cells were not reported by others.
In conclusion of this section, the Sox2+ cell compartment shows striking differences between neonatal and adult pituitary with regard to abundance and topographical appearance, pointing to a more active stem/progenitor cell compartment during the early-postnatal pituitary modeling process.
Higher proliferative activity in the stem/progenitor cell compartment of neonatal versus mature pituitary
The high abundance of Sox2+ cells in the newborn mouse pituitary indicates that a large fraction of the embryonic progenitor cells persists and only further matures after birth, thereby plausibly driving, or participating in, the early-postnatal maturation wave. We further analyzed whether the Sox2+ cell compartment at birth simply represents the population remaining from embryonic development [12,19], or whether it is still proliferating during the early-postnatal period. Pituitary vibratome sections of different ages (P1, P7, P21 and adult) were immunostained for Sox2 and the cell-cycling marker Ki67. Overall, Ki67+ cells are more abundant at early-postnatal than adult age when only rare cells are dividing (Supplementary Fig. S3A). During the first postnatal week, Ki67 is readily detected within, and in the vicinity of, the Sox2+ marginal layers including the wedge region (Fig. 3A, arrows; Supplementary Movies S6–S7) as well as in the Sox2+ cell clusters (Fig. 3B, arrows). Although less frequently, double (nuclear) Sox2+/Ki67+ cells are also observed within the AP's parenchyma (Fig. 3A, arrows). Ki67-containing Sox2+ cells decline toward adulthood. At P21 and adult age, the majority of dividing cells is Sox2-negative (Sox2−), although Ki67 is still sometimes detected in Sox2-cytoplasmic cells (Fig. 3A-P21, inset), and even more sporadically, in Sox2-nuclear cells (Fig. 3A, B, arrows; Supplementary Movies S8–S9). The Sox2−/Ki67+ cells as observed at the different ages may represent cells in which Sox2 expression went down before or during proliferation. In addition, it is not unlikely that Sox2 is at the same time excluded from the nucleus and/or precluded to enter the nucleus from the protein-synthesis location of the cytoplasm, thus explaining the cytoplasmic Sox2 signal in some Ki67+ cells. Alternatively, or additionally, the Sox2−/Ki67+ cells may embody other (nonSox2) cells that are proliferating such as hormonal cells. Some proliferating GH+ cells are found at P1 (Supplementary Fig. S3B, arrows); these double-immunopositive cells are more scarce at P21 and adult age, which is not unexpected, as proliferating cells in total are less abundant at the older ages. Staining for all hormones together and for Ki67 in AP cell cytospin preparations revealed that among the Ki67+ cells at P1 (10.6%±0.4; n=3), 15.2% (±2.7) are hormone positive, whereas in adult mice, 68.5% (±9.1) of the small number of Ki67+ cells (1.8%±0.1) contain hormones. These observations are in accordance with previous reports in rats that during neonatal development only part of the proliferating cells contain hormones, which at that time led to the hypothesis that the majority of the hormonal cells arising during the early-postnatal maturation phase derives from proliferating nonhormonal (stem/progenitor) cells (reviewed in [9]). Finally, some of the new hormonal cells may also originate through transdifferentiation ([20,21]; reviewed in [9]), although this process is not directly expected to contribute to the gland's expansion in size. We analyzed co-expression of GH and PRL as embodiment of the transition phase during transdifferentiation of somatotropes and lactotropes. Colocalization is found in some but not many cells, and not different between the different ages (Supplementary Fig. S4, arrows), analogous to previous studies in rats [6].
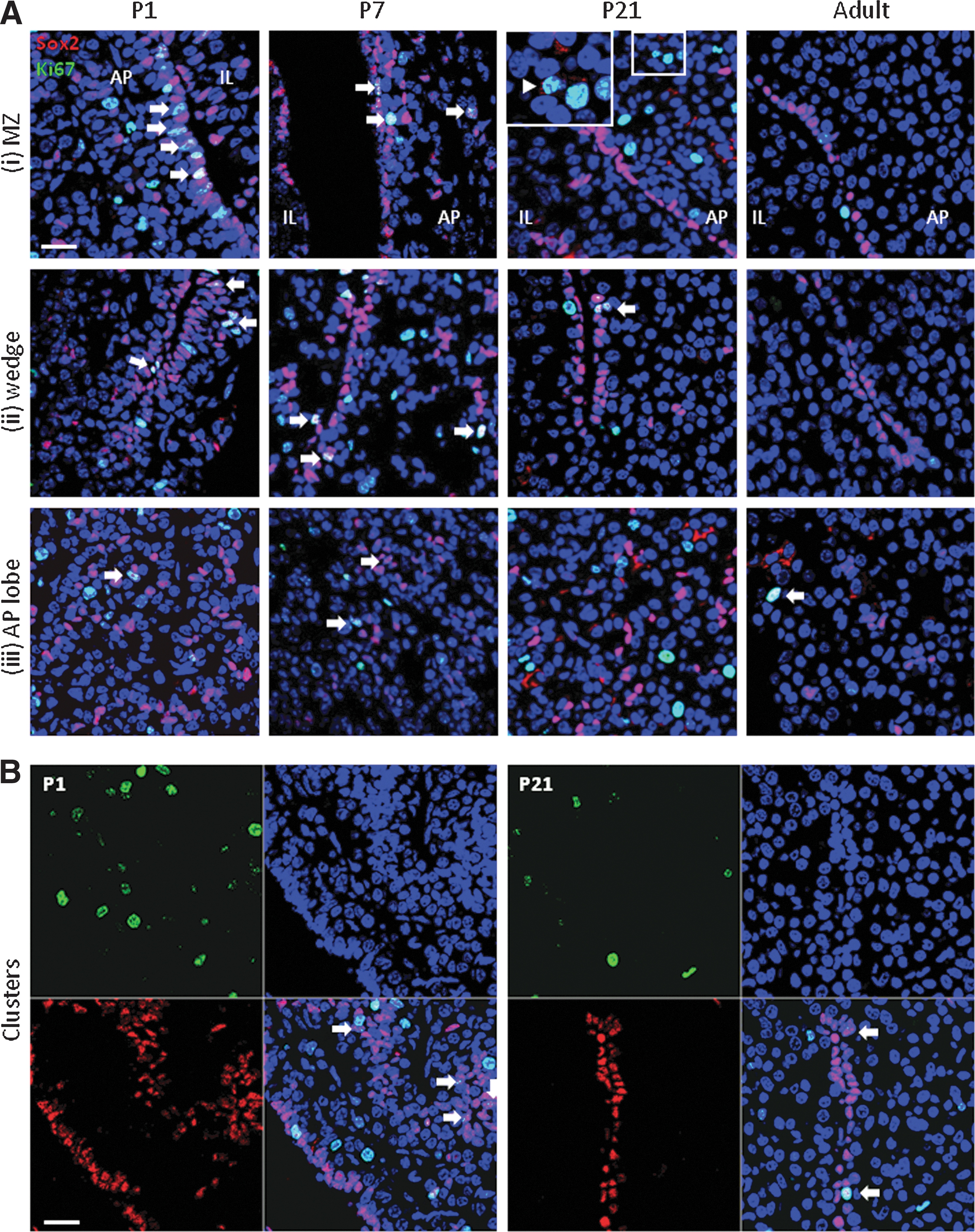
Sox2 expression and proliferation in the pituitary at different time points of postnatal development.
Together, our findings demonstrate higher proliferative activity in the pituitary stem/progenitor cell compartment of the early-postnatal than of the older mouse, further suggestive of their involvement in the gland's maturation.
Phenotype and physical arrangement of Sox2+ cell clusters in the postnatally developing pituitary
As just mentioned, confocal scanning of the pituitary sections revealed peculiar arrangements of Sox2+ cells into clusters, thereby surrounding a nonvascular lumen (Fig. 2C; Supplementary Fig. S5A, arrows) (as also described before in adult mice, see [11]). These Sox2+ cell clusters are more prominently observed at the younger ages, especially at the wedge region. Since the latter clusters are located adjacent to the cleft and sometimes oriented in line with the wedge or the wedge's invagination (see Supplementary Fig. S5A), and because the bordering Sox2+ cells display an abundance (i.e., single- or multiple-cell layer) comparable to that of the cleft-lining Sox2+ cells, we hypothesized that these clusters may sprout from the cleft. Three-dimensional reconstruction of z-stacks through 50-μm sections indeed reveals wedge Sox2+ cell clusters that are connected to the cleft (Supplementary Fig. S5B, arrow; Supplementary Movies S10–S13).
In addition, in the vicinity of other regions of the cleft, comparable Sox2+ cell clusters are observed, some of which are also found connected to the cleft's marginal layer (Supplementary Fig. S5C, arrows; and data not shown). In adult mice, Sox2+ clusters are generally seen more distal from the MZ than in neonatal mice, suggesting that they “drift away” (and eventually may lose connection) when the lobe expands during postnatal maturation.
Finally, also in the AP's parenchyma Sox2+ cell clusters are present (Fig. 2C), which show the same age-related phenotype as in the submarginal zone and wedge, namely a more abundant appearance of the Sox2+ cells and a sometimes multicellular border in the neonatal mouse whereas mostly unicellular in the adult mouse. Thus, the parenchymal clusters resemble the clusters near the MZ and in the wedge region as just described, suggesting that they all have a common origin. Cell proliferation in the parenchymal clusters still occurs but looks less active (Fig. 3B, arrows; and data not shown), indicating that expansion mainly takes place in the cleft and wedge region, before or while “moving” into the parenchyma.
Together, detailed in situ examination encourages the idea that Sox2+ cell clusters originate from the cleft, eventually disconnecting during development when the pituitary matures and expands.
Hormones and Sox2 are mainly not coinciding
To analyze the Sox2+ cells in relation to the hormonal cells during postnatal development, double immunofluorescence was performed in sections of neonatal (P1, P4, P7), P21, and adult pituitaries. As shown in Fig. 4, colocalization of Sox2 and hormones is in general not observed at the ages examined. Only sporadically, cells are detected in which hormones and nuclear Sox2 are colocalized (Supplementary Fig. S6, arrows; and data not shown). More often but still scarce, cells are observed in which hormone immunoreactivity sits together with cytoplasmic Sox2 signal (Fig. 4, arrows and insets) (as also reported before in adult mice, see [11]). Of note, the submarginal strip that is mostly devoid of Sox2+ cells contains some hormonal cells, in particular GH+ and αGSU+ cells (at all ages tested), and sometimes PRL+ but virtually no ACTH+ cells.

Sox2 and hormone expression in the pituitary during postnatal development. Immunofluorescent staining of pituitary sections for Sox2 (red) and hormones (green) at the different time points of postnatal development indicated. Representative pictures are shown of the domains demarcated in Fig. 2A. Sox2 typically does not coincide with hormone expression at all ages tested. Some sporadic cells with colocalization of cytoplasmic Sox2 and hormone are indicated (arrows; insets show some additional double-immunopositive cells). In addition, budding of Sox2+ cells from the periluminal layer (encircled) and “migratory streams” (long arrow) are indicated. Some co-arrangements of Sox2-nuclear cells with hormonal cells are remarkable (arrowheads). Scale bar=20 μm.
Our observations indicate that in general, Sox2 expression is not coinciding with hormone expression, thus suggesting that Sox2 should go down before differentiation can take place (see Discussion). Alternatively, hormonal cells after birth originate from self-duplication, although proliferation studies (see results above and [9,19]) do not advance this process as the sole and main mechanism and rather support participation of nonhormonal cells.
The stem/progenitor cell population of the neonatal pituitary displays higher molecular and functional stemness activity than the population of the adult gland
SC-SP cells were isolated from neonatal and adult AP glands, and microarray analysis was performed. Expression of a selection of stemness- and embryogenesis-related genes (see [1]) was evaluated (Table 1). In general, the neonatal SC-SP shows higher expression levels of these genes, pointing to a higher developmental activity within the stem/progenitor cell compartment at this stage. Sox2 mRNA signals were below background level in the particular microarray analyses, probably due to the presence of only one single probe on the particular arrays.
For selection of genes, see [1].
Average fold difference in gene expression between neonatal (P2-P5) and adult stem cell-side population. Genes are considered significantly upregulated when >1.5-fold (P<0.05) and significantly downregulated when <1.5-fold (meaning a ratio <0.6; P<0.05).
ND, not detectable.
To investigate whether the more abundant stem/progenitor cells of the neonatal pituitary are also more active in terms of functional stem-cell properties, we compared pituisphere-forming and differentiation capacity between neonatal and mature AP. The spheres in adult AP cell cultures most often display a smooth contour and bright phase-contrast appearance (Supplementary Fig. S7, arrows), as also reported earlier [10,11]. Sox2 is found in the nucleus of these cells (Fig. 5B-adult-d0; [11]). Although pituispheres from P21 pituitary are comparable, the neonatal AP cell cultures contain not only bright round-shaped spheres but also numerous, more irregular sphere-like structures of similar brightness, often deposited at the bottom of the culture dish (Supplementary Fig. S7, arrowheads). These sphere-like structures are also composed of Sox2-nuclear cells (Fig. 5B-P1-d0), thus indicating that they represent stem/progenitor cell-derived spheres that develop an irregular shape. Clonal origin of the spheres obtained by this protocol was clearly demonstrated earlier [10,11]. Moreover, no hormone-immunoreactive cells were found within the spheres formed, thereby excluding formation by random aggregation and further supporting clonal development. Quantification of all spheres (smooth and irregular) revealed that the proportion of sphere-initiating cells is highest at birth (∼2% at P1), then rapidly declines to P7 (∼1%), and further decreases to adult age (∼0.2%) (Fig. 5A). This finding of declining stem/progenitor cell proportions between birth and adulthood is in agreement with the recent report of Gaston-Massuet et al. [22] showing a decrease in colony-forming cells during postnatal development. Of note, the 10-fold difference in sphere-initiating cells that we observe between P1 and adult pituitary is not fully explained by the 2.5-fold higher abundance of Sox2+ (stem/progenitor) cells at P1 versus adult age. Therefore, stem/progenitor cells at P1 are not only more abundant but also more active. It should further be remarked that the sphere-initiating proportion in the adult AP is higher than reported earlier (0.02%, see [10]); first, because irregular spheres – which are also, although more rarely, present in adult AP cell cultures –, were included here since expressing Sox2; second, because counting was done on day 5 of culture, whereas earlier on day 7 when a high number of spheres are not perceptible anymore because of embedded and disappeared in clumps and aggregates (see [10]); and third, because of further-developed skills in the pituisphere assay. In addition, exact numbers (see e.g., [10 –12,22]) may vary with not only the use of different mouse strains but also of different stem/progenitor cell quantification assays (e.g., spheres versus colonies).

Sphere-forming and multipotent differentiation capacity within AP cells from neonatal to adult age.
We further explored functional stem-cell activity by analyzing the differentiation capacity of the stem/progenitor cells. Comparable (nondeposited and smooth) pituispheres from P1 and adult AP cell cultures were transferred to differentiation conditions (Matrigel) and cultured for another 7 days (standard method as described in [11,12]). Immunostaining revealed that spheres from both ages contain hormonal cells (Fig. 5B-d7; immunostained for all hormones together), representing all phenotypes (as observed after immunostaining for GH, PRL, αGSU and ACTH separately; data not shown). These data show that there is no difference in multipotential capacity between P1 and adult stem/progenitor cells. Next, we addressed the question whether there was a distinction in differentiation speed and activity. After 3 days in differentiation conditions, hormone+ cells are observed in all P1 spheres, whereas only half of the adult spheres (48%±9; n=3) display hormone immunoreactivity. A closer look at day 7 of culture revealed further quantitative differences; some of the P1 spheres are completely composed of hormone+ cells (data not shown), whereas such fully differentiated spheres were not observed in the adult cultures. Moreover, there remain ∼10% (11%±10; n=3) completely hormone-negative spheres in the adult cultures at day 7. These results indicate that the pituitary stem/progenitor cells of neonatal mice display swifter and higher differentiation competence and thus are more active in this functional aspect than the stem/progenitor cells of adult mice.
When Sox2+ cells are still present in the spheres after differentiation culture, cells with nuclear Sox2 signal are mostly observed in the interior of the sphere and are typically hormone-negative (Fig. 5B), whereas Sox2-cytoplasmic cells (found at lower incidence than Sox2-nuclear cells) are present within the borders of the sphere and almost all of them (∼90%) contain hormone immunoreactivity (see Fig. 5B-adult-d7, inset; Supplementary Movie S14). In keeping with observations in vivo, Sox2-cytoplasmic cells are much scarcer in P1 than in adult pituispheres (Fig. 5B).
Together, our findings demonstrate that, compared with the adult pituitary, the stem/progenitor compartment of the neonatal pituitary displays more prominent expression of stemness factors, including multiple components involved in pituitary embryonic development, as well as more activity with a higher number of functionally active cells and higher readiness and competence for differentiation.
Discussion
Our study scrutinized the stem/progenitor cell compartment of the mouse pituitary gland at different stages of postnatal development and revealed prominent differences between the early-postnatal maturation phase and mature age. The proportion of stem/progenitor cells, as identified by different methods (SC-SP, Sox2, and sphere formation), is higher in neonatal animals. Moreover, the compartment is more actively proliferating at the younger age, and displays a higher stem-cell expression profile and functionality. The topographical structures including the marginal niche, wedge region, and cluster arrangements are also more pronounced and populated with Sox2+ cells (uni- versus multicellular), and appear more active (with more budding and bifurcation) in the neonatal gland. Together, our observations support an activated phenotype of the pituitary stem/progenitor cell compartment during the early-postnatal vivid modeling of the gland. Earlier studies in neonatal rats suggested the hypothesis that the majority of the newly developing endocrine cells originate from nonhormonal cells, rather than from replication of already differentiated cells ([3 –8]; reviewed in [9]). In support of this hypothesis, our study demonstrates a low proliferation rate of hormonal cells in the neonatal mouse and an activated phenotype of the stem/progenitor cell compartment. The decline of the stem/progenitor (SC-SP/Sox2+/sphere-initiating) cells during the first postnatal week further strengthens the idea that they give rise to the newborn hormonal cells. Moreover, when the first postnatal growth wave is finalized (P21), the stem/progenitor cell compartment indeed presents as less active.
Rather unexpectedly, co-expression of Sox2 with hormones is only seldom encountered at all ages examined. However, it is not unreasonable to propose that Sox2 expression should be extinguished or at least largely downregulated (below detectable levels) before differentiation can take place. Similar findings were reported in other cells and tissues [23 –25]. In further support, colocalization is, in general, also not detected in the embryonic pituitary (L. Raetzman, personal communication), where the Sox2+ progenitor cells should give rise to the developing hormonal cells [12,19,26]. Our finding of some sporadic cells containing both hormone and nuclear Sox2 in the postnatal pituitary may embody fortuitous observations (snapshots of speedily developing differentiation in these cells) and/or abnormal events. In addition, it is likely that exclusion of Sox2 from the nucleus precedes, or coincides with, its expression fall to stop its transcriptional role as gatekeeper of the stem/progenitor cell phenotype (as also earlier suggested, see [11]). Indeed, nucleocytoplasmic shuttling of Sox2 (through posttranslational modification) has been found to abrogate transcription of Sox2 target genes [23], further in agreement with findings on other transcription factors (Sry, Sox9, Sox10; see [27]). The transportation of Sox2 from the nucleus may explain why hormones are sometimes found co-localized in a cell with Sox2 signal in the cytoplasm (both in situ and in pituispheres). Moreover, the Sox2-cytoplasmic cells are proportionally more found in the IP and MP (as gated in the FACS profiles), again supporting the idea that they represent progenitor cells in further states of development [17,18]. Our data demonstrate that Sox2 marks a rather large population of AP cells. It is proposed that Sox2+ cells do not only represent stem cells, but also stadia of further developed progenitor, transit-amplifying (TA), and precursor cells. Fauquier et al. reached a similar conclusion using BrdU label retaining experiments revealing only few label-retaining (stem) cells within the Sox2+ cell population, and a large number of nonlabel-retaining (TA) cells [12]. A hypothetical scheme clarifying these ideas and proposing the different steps in hormone cell differentiation coupled to the subcellular localization and extinction of Sox2 expression in pituitary stem/progenitor cells, as based on our observations, is provided in Supplementary Fig. S8. Evidently, the other explanation that Sox2+ cells do not give rise to the new hormonal cells after birth should not be overlooked. However, alternate possibilities do not seem to fill this gap, as proliferating hormonal cells and transdifferentiating cells are scarce. Sox2 lineage tracing studies using conditional (e.g., tamoxifen-inducible) approaches are needed to finally settle this issue, but await development of appropriate and fully reliable transgenic models.
Our study revealed some further intriguing findings on stem/progenitor (Sox2+) cell topography. First, Sox2+ cells are most prominent at the junction of the AP and IL and establish a sprouting pattern from this wedge region. In the older literature, this zone has been referred to as the “sex zone” where some endocrine cell types were postulated to originate [28]. This wedge region is expanded in the neonatal pituitary and Sox2+ cells in the neighborhood are clearly more abundant than at later age, further supporting the germinal nature of this area. Interestingly, expanded wedges with numerous Sox2+ cells are also observed during the acute response to transgenically inflicted injury in the pituitary (Fu and Vankelecom, unpublished observations). Similar to other tissues, the stem/progenitor cell compartment may recapitulate developmental processes during the reaction to injury [29 –33]. Second, the cleft invaginates into the lobe's parenchyma and sometimes bifurcates, again most prominent at the younger age. In the neighborhood, as well as at other spots in the submarginal zone, Sox2+ cells appear as clusters surrounding an inner lumen. Intriguingly, these clusters are connected to the periluminal Sox2+ cell layer and cleft. Together with the morphological resemblance of the clusters more distal from the cleft, it is tempting to hypothesize that Sox2+ cell clusters initially germinate from the periluminal layer (which may be regarded as the “primary” niche) and during expansion of the gland, eventually disconnect and form additional, “secondary” niches within the parenchyma. The existence of multiple niches is considered beneficial for swift and/or subtle adaptation of the pituitary cell composition to the continuously changing endocrine demands [1,11]. In older studies, cyst-like structures have been described within the gland (see [9]) that were proposed to be derived from the marginal cells, as particularly observed in damaged and activated conditions (such as in rat pituitaries grafted under the renal capsule [34]). The clusters in the parenchyma show the same age-related phenotype as in the submarginal zone and wedge region, further supporting their common origin. Again, time-specific inducible Sox2 lineage tracing will help to clarify these ideas. Of note, proliferative activity of the Sox2+ cells appears less prominent in the clusters than in the periluminal layer, thus suggesting that the principal expansion occurs around the cleft and that further proliferation is not needed anymore, or not supported anymore by trophic factors from the “primary” niche, once clusters have moved away.
In addition to the forming clusters, Sox2+ cells also seem to bud from the periluminal layer at some locations and to form “migratory streams” toward the lobe's parenchyma, again more frequently encountered in the neonatal pituitary than in the more mature gland. Migration may involve epithelial-mesenchymal transition (EMT), a process suggested to be implicated in embryonic development of the gland [1,35,36] as well as in the behavior of stem cells in other, postnatal tissues [37,38]. Interestingly, genes involved in the EMT pathway appear higher expressed in the neonatal than the adult SC-SP (data not shown). These observations encourage the idea that periluminal stem/progenitor cells at some places migrate toward the AP, supplementary to the formation of secondary niches by cluster outgrowth. Of further note, the submarginal strip is mostly devoid of Sox2+ cells and contains some Ki67+ as well as hormonal cells, in particular GH+ and αGSU+ cells but virtually no ACTH+ cells. αGSU represents the first pituitary marker in the hierarchy of embryonic development [1,39]. The peculiar absence of ACTH in this zone has very recently been confirmed in a study demonstrating interdigitated ACTH cell networks [40], and may be related to the early (embryonic) development of corticotropes, before the other cell types appear [6,41].
In conclusion, our study demonstrates that the neonatal stem/progenitor cell compartment shows an activated phenotype with regard to abundance, proliferative condition, and stem cell-associated molecular and functional properties, thereby arguing for its participation in the early-postnatal maturation process of the pituitary gland. Our study is expected to elicit renewed interest in the mechanisms of pituitary early-postnatal modeling.
Footnotes
Acknowledgments
This work was supported by grants from the Fund for Scientific Research-Flanders (Belgium) (F.W.O.-Vlaanderen), the Research Fund (BOF Onderzoeksfonds) of the K.U.Leuven, Joint Research of the FWO and MOST (Ministry of Science and Technology, P.R. China), and the National Natural Science Foundation of China. LG is a Research Fellow of the FWO and QF receives a SBA Scholarship (China - K.U.Leuven). The authors are very grateful to Vik Van Duppen (Hematology, K.U.Leuven) for FACS analysis and sorting, to the VIB-MAF (K.U.Leuven) for microarray analysis, and to Yvonne Van Goethem (Lab. Tissue Plasticity) for technical support.
Author Disclosure Statement
This work does not include conflicts of interest of any kind.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.