
Abstract
Avian species are important model animals for developmental biology and disease research. However, unlike in mice, where clonal lines of pluripotent stem cells have enabled researchers to study mammalian gene function, clonal and highly proliferative pluripotent avian cell lines have been an elusive goal. Here we demonstrate the generation of avian induced pluripotent stem cells (iPSCs), the first nonmammalian iPSCs, which were clonally isolated and propagated, important attributes not attained in embryo-sourced avian cells. This was accomplished using human pluripotency genes rather than avian genes, indicating that the process in which mammalian and nonmammalian cells are reprogrammed is a conserved process. Quail iPSCs (qiPSCs) were capable of forming all 3 germ layers in vitro and were directly differentiated in culture into astrocytes, oligodendrocytes, and neurons. Ultimately, qiPSCs were capable of generating live chimeric birds and incorporated into tissues from all 3 germ layers, extraembryonic tissues, and potentially the germline. These chimera competent qiPSCs and in vitro differentiated cells offer insight into the conserved nature of reprogramming and genetic tools that were only previously available in mammals.
Introduction
I
Although avian ESC and primordial germ cell (PGC) lines have been established [11,12], they have not been used in gene targeting studies, mostly likely because they have not been clonally isolated nor are they highly proliferative in extended cultures. Avian ESCs and PGCs have often demonstrated a significant decrease in their potential to form chimeric animals after extended culture, typically <10 passages, further reducing their potential in in vivo gene target studies. Additionally, many of these avian lines have not been shown to robustly undergo directed in vitro differentiation into multiple lineages with phenotypic characteristics similar to mammalian pluripotent cells. Given some of the issues that plague the development of avian ESCs, the probability that avian iPSCs could be generated and show diverse differentiation potential in vitro and in vivo seemed unlikely. Generating nonmammalian iPSCs poses additional challenges and difficulties. Intuitively, species-specific or at least species-related reprogramming factors maybe required for proper reprogramming. However, little is known of the required reprogramming factors in phylogenetically diverse species. Previous studies showed that iPSC could be derived from human, mouse, pig, primate, and even rhinoceros fibroblast using the human reprogramming factors [5,6,13,14]. These results indicated the widely conserved reprogramming process among species and the possibility of deriving iPSCs from phylogenetically diverse species such as avian.
Avian embryonic models have a long history of providing critical new insights into developmental biology including organ function [15,16], disease progression (e.g., Pompe disease) [17], eye disorders [18], and many others [19,20]. The advantage of avian species is their relative size and ease of access to the embryo for manipulation. Cells and tissues, including whole sections of the spinal column, can be transplanted into the avian embryo and can be monitored in real time during development [21]. This is not possible in mammalian species. Moreover, the quail-chicken chimera is an attractive and widely used model for developmental patterning and cell fate studies given that cells can be readily tracked in this model [22,23]. The quail also has a short generation interval (3–4 generations per year) [24], facilitating genetic selection studies and experiments requiring multiple generational observations [25]. Robust clonal and feeder-free iPSC lines capable of generating lineage-committed avian cells offer new opportunities to manipulate and study developmental process both in vitro and in vivo.
In this report we demonstrate for the first time in a nonmammalian species the successful generation of a highly proliferative quail iPSC (qiPSC) population amenable to genetic manipulation, that undergoes advanced directed neural differentiation in vitro and is capable of generating live chimeric offspring. The extensive contribution of qiPSCs to chimeric animals even at late passages demonstrates that these cells are true iPSCs. These first iPSCs in a nonmammalian species demonstrate the highly conserved nature of the reprogramming genes and provide for future mechanistic developmental studies in well-characterized avian models.
Materials and Methods
Cell culture and transduction
Quail embryonic fibroblasts (QEFs) were isolated from day-11 embryos and cultured in fibroblast medium [Dulbecco's modified Eagle's medium (DMEM) high glucose (Hyclone) with 10% fetal bovine serum (Hyclone), 4 mM L-glutamine (Gibco), and 50 U/mL penicillin and 50 μg/mL streptomycin (Gibco)] in 5% CO2 at 37°C. Cells were split using 0.05% trypsin (Gibco) upon reaching confluence. For transduction, a total of 150,000 QEF cells was plated in 1 well of a 12-well plate. After 24 h, QEFs underwent lentiviral transduction utilizing the viPS kit (Thermo Scientific) with viruses containing the human stem cell genes POU5F1, NANOG, SOX2, LIN28, KLF4, and C-MYC under the promoter of human elongation factor-1 alpha (EF1-α). Transduction was performed in the presence of 1× TransDux (System Biosciences). The QEFs were trypsinized 24 h after transduction and passaged onto inactivated feeder cells in ESC expansion medium [DMEM/F12 (Gibco), supplemented with 20% knockout serum replacement (KSR; Gibco), 2 mM L-glutamine (Gibco), 0.1 mM nonessential amino acids (Gibco), 50 U/mL penicillin/50 μg/mL streptomycin (Gibco), 0.1 mM β-mercaptoethanol (Sigma-Aldrich), and 10 ng/mL basic fibroblast growth factor (bFGF; Sigma-Aldrich and R&D Systems)]. The qiPSCs were manually harvested and plated on Matrigel (BD Biosciences; diluted 1:100 in DMEM/F12) coated dishes in mTeSR1 medium (Stemcell Technologies). The qiPSCs were mechanically dissociated using a glass Pasteur pipette every 4–5 days. For clonal expansion, qiPSCs were transduced with green fluorescent protein (GFP) viral vector and single cells were FACS sorted into individual wells of a 96-well plate.
Alkaline phosphatase and periodic acid Schiff's staining
Alkaline phosphatase (AP) staining was carried out with VECTOR Red Alkaline Phosphatase Substrate Kit (Vector Laboratories) according to the manufacturer's instructions. Periodic acid Schiff (PAS) staining was performed by 4% paraformaldehyde fixation for 5 min. The PAS (Sigma-Aldrich) was added to the plate and incubated at room temperature for 5 min, followed by phosphate-buffered saline (PBS; Hyclone) washes for 3 times. Schiff's reagent (Sigma-Aldrich) was added and incubated at room temperature for 15 min, followed by 3 washes with PBS and then observation.
Immunocytochemistry
Protocol for immunostaining followed the methods previously reported [6]. Primary antibodies used were POU5F1 (R&D Systems), SOX2 (R&D Systems), TUJ1 (Neuromics), alpha smooth muscle actin (αSMA) (Santa Cruz), SOX17 (Santa Cruz), SSEA4 (Developmental Studies Hybridoma Bank), TRA-1-60 (Chemicon), TRA-1-81 (Chemicon), Hu C/D+ (Invitrogen), MAP2+ (Millipore), GFAP (Chemicon), and O4 (Chemicon). Secondary antibodies used in this study were all from Invitrogen. Cell observations and images were captured on Ix81 with Disc-Spinning Unit (Olympus) using Slide Book Software (Intelligent Imaging Innovations).
Proliferation and telomerase activity
Proliferation assay was performed by manual counts (n=3) at 12, 24, 36, and 48 h after plating. Population doubling time was determined using an exponential regression curve fitting (
Embryoid body formation and differentiation
Embryoid bodies (EBs) were formed by plating 2.0×106 qiPSCs in mTeSR1 medium and 0.1 mM Y-27632 ROCK inhibitor (Calbiochem) in an AggreWell plate (Stemcell Technologies). After 24 h, aggregates were harvested and maintained in mTeSR1 medium for 7 days. Differentiation was assessed by reverse transcription polymerase chain reaction (RT-PCR) using the primers in Supplementary Table S1 (Supplementary Data are available online at
Neural differentiation
To induce neural differentiation, qiPSCs were sequentially cultured in neural derivation medium [DMEM/F12 supplemented with 200 mM L-glutamine, 4 ng/mL bFGF, and 1×N2 (Gibco)] for 12 days, proliferation medium [AB2 (Aruna) medium supplemented with 200 mM L-glutamine, 1×aruna neural supplement (ANS), and 20 ng/mL bFGF] for 7 days, and then in differentiation medium (AB2 medium supplemented with 200 mM L-glutamine, 1×ANS, and 10 ng/mL leukemia inhibitory factor [LIF]) continuously.
Production of chimera
Stage X White Leghorn chicken embryos were used to produce chimeras. Egg shells were removed with a Dremel rotary tool to make an injection window (Supplementary Fig. S2A). The qiPSCs were introduced into the subgerminal cavity using a glass micropipette (Supplementary Fig. S2B) with pressure controlled microinjector (Parker Automation). Cells were prepared at concentration of 10,000,000 per mL and injected 1 μL per embryo, which should be approximately 10,000 cells. The window was sealed by hot glue (Supplementary Fig. S2C) after injection and eggs were incubated at 37.8°C.
RNA isolation, DNA isolation, PCR, and sequencing
RNA was isolated using RNeasy QIAprep Spin miniprep Kit (Qiagen) per manufacturer's instructions. Genomic DNA was removed using gDNA eliminator columns (Qiagen). mRNA extractions were transcribed into cDNA using iScript cDNA Synthesis kit (Bio-Rad Laboratories). DNA was isolated using DNeasy kit (Qiagen) following the manufacturer's instructions. Primers used in PCR and RT-PCR are listed in Table S2. Sequencing verification of hPOU5F1 was performed by extracting DNA from agarose gels after electrophoresis and sequencing. The resulted sequence was compared by Blast in the NCBI database to both human and chicken genomes.
Results
qiPSCs display morphological characteristics consistent with a pluripotent cell type
The generation of qiPSCs was initiated by testing the lentiviral transduction efficiency of isolated QEF (Fig. 1A) with an eGFP reporter construct using both GeneJammer and TransDux transduction reagents. A 20 multiplicity of infection (MOI) transduction with Transdux resulted in the highest efficiency with 40.5% green fluorescent protein (GFP)–positive cells (Supplementary Fig. S1). QEFs were then transduced with the 6 human pluripotency genes hPOU5F1, hNANOG, hSOX2, hLIN28, hC-MYC, and hKLF4 driven by the EF1-α promoter with each construct in individual lentiviral vectors. After 24 h, cells were replated on feeder cells in stem cell expansion medium.
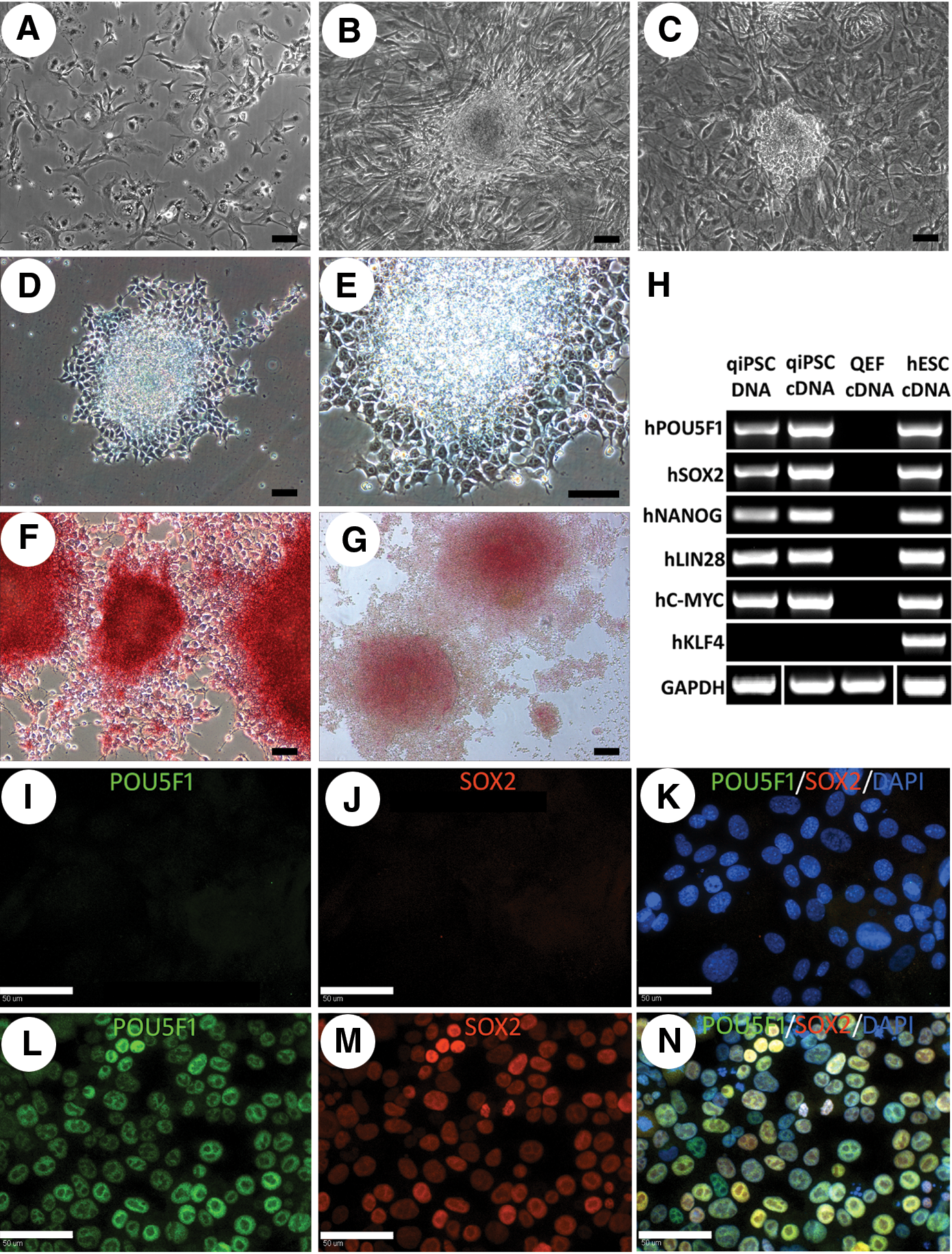
Derivation of qiPSCs from QEFs. QEFs prior to addition of reprogramming factors
Colonies began to emerge 6 days after transduction with irregular-shaped borders and fibroblast-like cell morphology (Fig. 1B). These initial colonies failed to proliferate and expand, indicating that these colonies were not fully reprogrammed. Potential qiPSCs were observed around 17 days after transduction and grew as compact colonies (Fig. 1C). The compact colonies were mechanically picked and initially replated on feeder plates in stem cell expansion medium. However, replated cells failed to proliferate and appeared apoptotic. Additional colonies were collected and replated on Matrigel-coated plates in mTeSR1 stem cell medium. This system supported the growth and expansion of colonies and subsequent qiPSC expansion was performed using this system.
Morphologically, qiPSC colonies were highly refractive and at the single-cell level they showed clear cell borders, high nuclear-to-cytoplasm ratio, and prominent nucleoli (Fig. 1D, E). The qiPSCs were strongly positive for AP and PAS staining (Fig. 1F, G). PCR and RT-PCR using human-specific primers revealed that 5 out of 6 pluripotent stem cell factors, hPOU5F1, hSOX2, hNANOG, hLIN28, and hC-MYC, were integrated and expressed in qiPSCs, while hKLF4 was not present (Fig. 1H). Immunocytochemistry revealed that POU5F1 and SOX2 proteins were absent in QEFs (Fig. 1I–K), but positive in qiPSCs (Fig. 1L–N). Immunocytochemistry of qiPSCs showed that cells were negative for the pluripotency markers SSEA4, TRA-1-81, or TRA-1-60.
qiPSCs are highly proliferative, express pluripotent markers, and are capable of clonal expansion after genetic manipulation
Rapid proliferation and high levels of telomerase activity are hallmarks of pluripotent stem cells. To determine the doubling time, plated cells were quantified every 12 h for 48 h. The population doubling time of qiPSCs was 16.6 h, much faster than the QEF parent cell line (36.9 h, P<0.01) (Fig. 2A). The qiPSCs are highly proliferative and are passaged every 4 days. Cells have been maintained for more than 50 passages without loss of the pluripotent phenotype. Telomerase activity revealed a significant (P<0.01) increase of >11-fold from 8.4 total product generated (TPG) in QEFs to 95.3 TPG in qiPSCs (Fig. 2B). Telomerase activity of qiPSCs was comparable to that of the positive control HeLa cell line (50.8 TPG, P=0.07), which indicates the immortality of qiPSCs. Moreover, these cells were capable of clonal expansion after genetic manipulation. The qiPSCs were transduced with the eGFP gene, resulting in a 36.1% GFP+ population (Fig. 2C–E). Single GFP+ cells were FACS sorted into each well of a 96-well plate and colonies were found in 2 wells 9 days after sorting (Fig. 2F, G). Flow cytometry analysis of cells expanded from 1 colony showed that >96% of the cells still expressed GFP after serial subculture (Fig. 2H).

qiPSCs demonstrate rapid proliferation, high levels of telomerase activity, and clonal expansion after genetic manipulation qiPSC doubling time was 16.6 h (n=3), significantly faster than the QEF cells (36.9 h; P<0.01)
EB differentiation of qiPSCs resulted in the formation of all 3 germ layers
To derive EBs, qiPSCs were plated in AggreWell plates for 24 h and then transferred to suspension culture in mTeSR1 medium for differentiation. Six days of suspension culture resulted in round and compact EBs from qiPSCs (Fig. 3A). The EBs were collected for RNA isolation and RT-PCR or replated for additional differentiation for 2 days in stem cell expansion medium without bFGF—the removal of which will drive differentiation (Fig. 3B). Results of RT-PCR showed expression of TUJ1 (ectoderm), PAX6 (ectoderm), Vimentin (endoderm), and Brachyury (mesoderm) in EBs, but not in qiPSCs or QEF cells (Fig. 3C). Immunocytochemistry showed cells positive for TUJ1 (ectoderm, Fig. 3D), SOX17 (endoderm, Fig. 3E), and alpha smooth muscle actin (αSMA, mesoderm, Fig. 3F) in plated EBs. These results indicated that qiPSCs could differentiate into various cell types from all 3 germ layers.
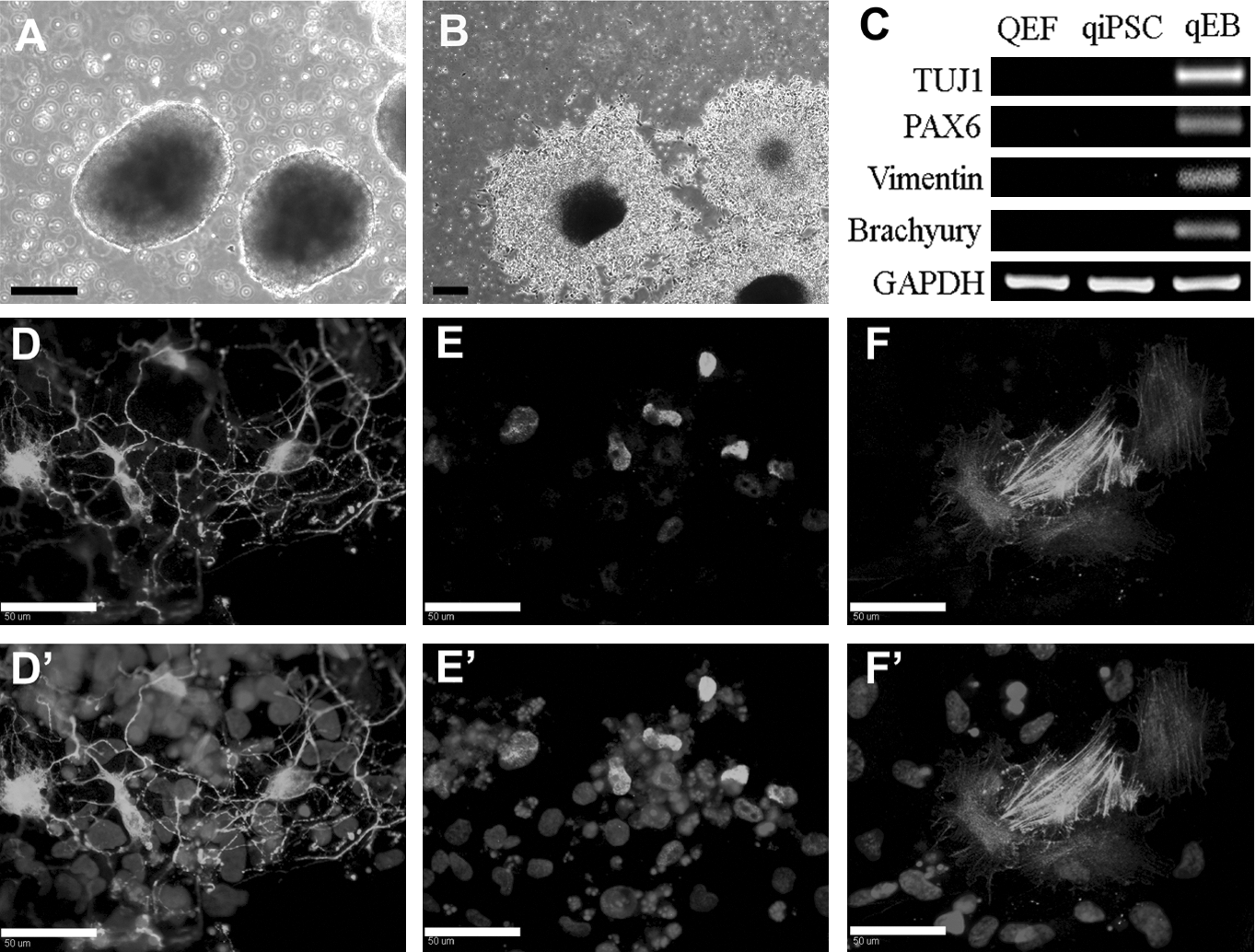
qiPSCs generate EBs that form all 3 germ layers. Compact EBs were formed after 6 days in culture
qiPSC differentiate in vitro into neurons, astrocytes, and oligodendrocytes
To derive neural cells, qiPSCs were subjected to a 3-step neural differentiation process. Cells were initially cultured in neural derivation medium for 12 days, proliferation medium for 7 days, and neural differentiation medium continuously thereafter. Immunostaining showed that these cells were positive for the neural proteins Hu C/D+ and MAP2+ (Fig. 4A, B) after 10 days of differentiation. A significant number of neurite extensions was observed after differentiation. Differentiated qiPSCs were found to be positive for the astrocyte- and oligodendrocyte-associated proteins GFAP and O4 after 23 and 39 days of differentiation, respectively, in neural differentiation medium (Fig. 4C, D). These data demonstrated the neural competence of qiPSC and differentiation into all 3 neural lineages, which has not been seen in any previous avian stem cell lines.
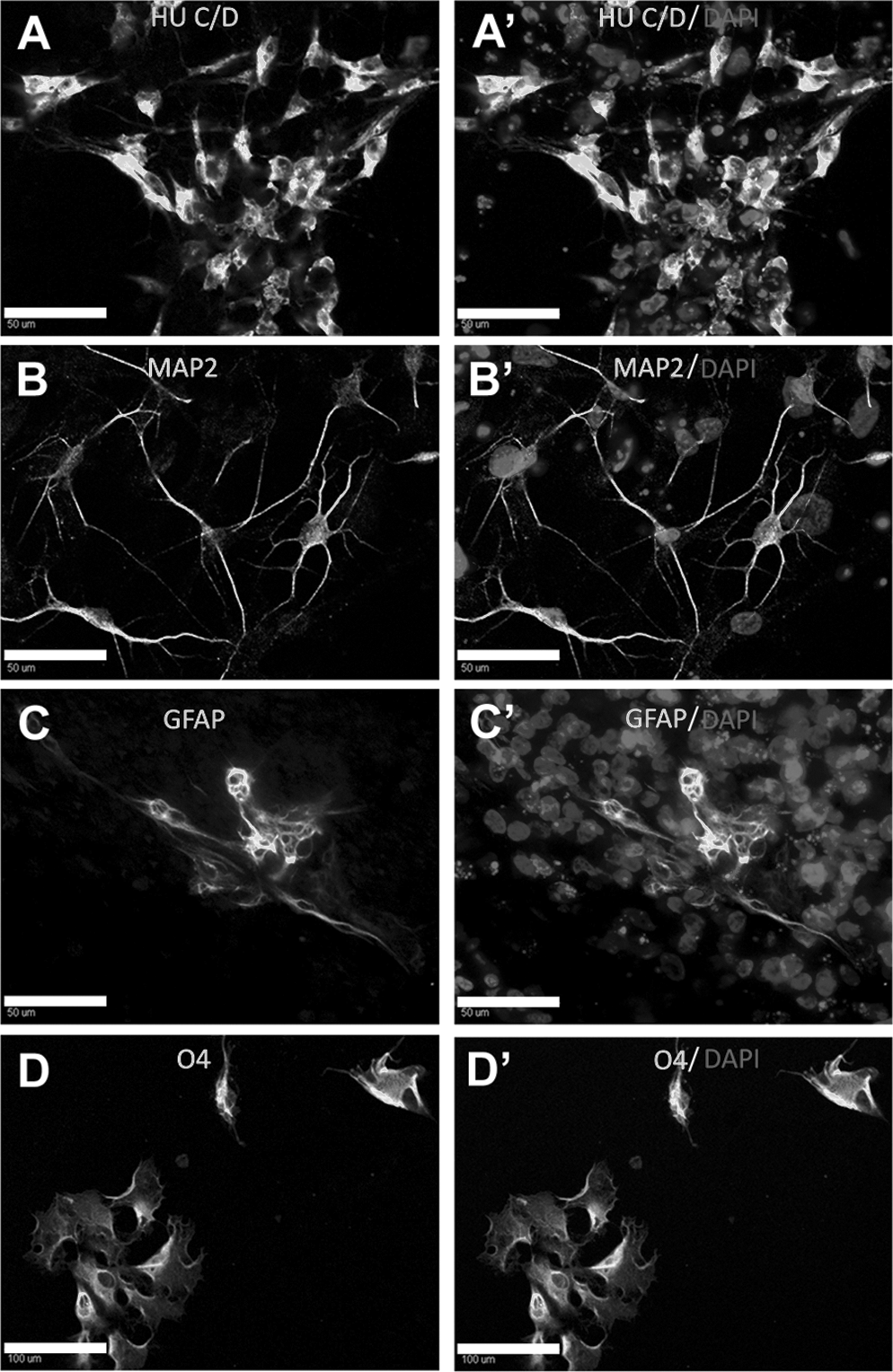
Directed differentiation of qiPSCs to 3 neural lineages. qiPSCs were subjected to a 3-step neural differentiation process, with cells first cultured in neural derivation medium for 12 days, then in proliferation medium for 7 days, followed by continual maintenance in differentiation medium. Neurite extensions could be found after culturing in differentiation medium for 48 days. Neuron-like cells expressing Hu C/D+
Incorporation of qiPSCs into chimeric embryo and generation of live offspring
To generate qiPSC-chicken chimeras, GFP+ qiPSCs at passage 26 were injected into the subgerminal cavity of stage X embryos (Supplementary Fig. S2). Embryos were incubated for 14 or 19 days and were then dissected to determine GFP+ qiPSC incorporation into embryos. GFP+ qiPSCs were incorporated in brain (Fig. 5A), eye (Fig. 5B), trachea/lung (Fig. 5C), heart (Fig. 5D), and yolk sac (Fig. 5F) tissues. PCR was performed for the human POU5F1 gene that was used to reprogram QEFs into iPSCs to further determine qiPSC contribution in chimeric animals. The qiPSCs were present in tissues from the ectoderm (brain, eye, and skin), endoderm (intestine, liver, and lung), mesoderm (muscle and heart), extraembryonic tissue (yolk sac), and the gonad (Fig. 5E, G and Supplementary Table S2). PCR products from qiPSCs from the yolk sac and skin were sequenced to validate that PCR primers were solely expanding the human POU5F1 sequence. Blast of the sequenced DNA amplified from these tissues showed identity of 99% to 100% for human POU5F1 genomic context sequence NC_000006.11, but only 72% to 74% maximum identity for chicken PouV (Oct4 homologue) mRNA NM_001110178.1 and no significant similarity to its genomic context sequence NW_001471503.1. These results indicate that qiPSCs incorporated and contributed to chicken embryonic tissues from all 3 germ layers, extraembryonic tissues, and potentially the germline.

Chimeric chicken embryos derived from qiPSCs. GFP+ qiPSCs were incorporated into the brain
Two additional rounds of injections were performed to generate chimeric chicks using passage 7 and passage 45 qiPSCs. Early passage qiPSCs were injected into 30 eggs. Three embryos developed to term and hatched after 21 days of incubation, while 9 were unfertilized and 18 embryos failed to hatch. Hatched offspring were not chimeric, but 2 chicks at late stages of embryonic development showed feather chimerism (Fig. 6A, B). Passage 45 qiPSCs were injected into 102 eggs, resulting in 47 live offspring. Two chicks died soon after hatching and the rest of 45 chicks were healthy, but no feather chimerism was observed. Brain, muscle, liver, and gonad tissues were collected from 15 chickens for PCR detection of qiPSC incorporation. Results showed that brain, liver, and gonad samples from 2 individuals were positive for the human POU5F1 reprogramming gene (Fig. 6C). The presence of feather chimerism with early passage cells and incorporation of qiPSCs into tissue with later passages indicate that these cells retain pluripotent characteristics following long-term culture and are still capable of contributing to multiple lineages.

Chimeric chickens derived from qiPSCs. Two chicks produced by low-passage qiPSCs (P7) determined to be at day 14
Discussion
iPSCs have been generated from numerous mammalian species [1–2,5 –10], but never before in a nonmammalian species. Here we report the first nonmammalian iPSCs and paradoxically these avian iPSCs were generated using human reprogramming factors. These chimera competent cells were capable of producing live offspring, achieved previously only in mice, rats, and pigs, and showed significant incorporation into all 3 germ layers, marking their true pluripotent nature [26]. Beyond contribution to chimeric offspring, mouse pluripotent cells are relatively easy to genetically modify and clonally isolate thus facilitating the generation of gene-targeted offspring [27]. The qiPSCs were also genetically manipulated and then clonally selected, as proliferative colonies. These in vitro characteristics will likely lead to site-directed gene insertion and selection of clonal colonies for generating chimeras. The qiPSCs show morphology consistent with previously established pluripotent stem cells at the single-cell level. The qiPSCs are highly positive for the stem cell markers AP, PAS, POU5F1, and SOX2 that have been previously used to characterize avian ESCs and PGCs [28 –32]. These iPSCs are highly proliferative with a doubling time of 16.6 h, similar to iPSCs from mouse [1] and pig [33]. In the chick/quail chimera model, qiPSCs contributed to fetal tissues from all 3 germ layers and extraembryonic tissues and ultimately contributed to tissues in live offspring. Although avian ESCs and PGCs have generated chimeric offspring [29,34], the qiPSCs differ because for the first time an avian stem cell exhibits the robust in vitro proliferative and clonal attributes needed for future gene-targeted birds.
In this study, qiPSCs were generated by transducing QEFs with 6 reprogramming factors and 5 of them (POU5F1, NANOG, LIN28, and C-MYC) were found incorporated into the genomic DNA and expressed in high level, which indicated their essential roles in the reprogramming process. KLF4 was not detected in qiPSCs in DNA or RNA level. In previous reports, KLF4 has been shown to be important in the reprogramming process [1,5,35,36]. However, many groups have shown successful reprogram in cells of multiple species without the KLF4 gene, demonstrating that it is not essential [2,37]. The qiPSCs generated without ectopic KLF4 showed robust pluripotency and were capable of chimera formation, indicating ectopic expression of KLF4 is not required in cell reprogramming in avian species. The successful use of human reprogramming factors to generate avian iPSCs suggests that direct reprogramming mechanisms are widely conserved among species. In the chicken, the cPouV and cNanog genes were found to be key factors in the maintenance of chicken pluripotency [38]. However, the homology of reprogramming genes between avian and human genomes is relatively low when comparing chicken—the only avian species with sequence data—and humans with homology ranging from ∼53% to 81% for POU5F1, NANOG, LIN28, C-MYC, and KLF4. SOX2 is the lone exception with homology of 94% between chicken and human genes. A previous report comparing the embryonic pluripotency gene-regulatory networks between mouse and chicken revealed that the mouse core pluripotency network is mediated largely by genomic sequence elements that are not conserved within the chicken [39]. This report suggests that species-specific or at least species-related reprogramming factors would be required for proper reprogramming of avian cells. However, based on the diverse number of species (mouse, human, pig, primate, and rhinoceros) [5,6,13,14] from which cells have been reprogrammed using human factors and now a nonmammalian species, we hypothesize that the reprogramming process is highly conserved. Therefore, direct reprogramming with transcriptional factors could be a universal strategy for generating iPSC lines in distantly related species. This would provide new species iPSCs for divergent species including species where ESCs are hard to isolate, maintain, and expand, which arguably would include all species other than those of primates and some rodents.
Quail and quail-chick chimeras have long been used in understanding the development of the nervous system [40 –43]. With the benefit of this avian system, numerous facets of neural development in the brain [15,40] and neural crest [41] have already been deciphered. In the present study, qiPSCs were found to significantly contribute to the brain and eye tissue when injected into stage X chicken embryos in an undifferentiated state. Upon proper signaling in vitro, qiPSCs could differentiate into a neural progenitor (TUJ1+) and all 3 lineages of neural cells—neuronal (Hu C/D+ and MAP2+), astrocyte (GFAP+), and oligodendrocyte (O4+)—in vitro. Despite preliminary neural differentiation of chicken ESCs or embryonic germ cell into neural cell [11,12], the desired diversity in neural developmental competence, forming neuronal both glia lineages, (TUJ1, MAP2, Hu C/D, GFAP, and O4) was not present or reported [44 –46]. Given that we have derived neurons, astrocytes, and oligodendrocytes using processes first developed in human pluripotent cells [47], the repertoire of “tools” and translational potential for embryonic graft studies is greatly enhanced. Temporal and spatial studies investigating the role and interaction among neural cell types during embryonic development are now possible. Pluripotent qiPSC-derived neural cells can be compared with tissues from fetal and adult organisms faster and in relevant but comparatively simple models in contrast to mammalian systems [21].
The methods used here for qiPSCs overcome impediments inherent to avian ESCs and PGCs. Previously quail or chicken PGCs [32,34] and ESCs [29] contributed to chimeras when injected into embryos immediately after collection from the donor embryo or after only a few passages. After serial subculture in this system, qiPSCs still efficiently incorporated into tissues from all 3 germ layers in chimeric embryos at passages 26 and 45. This will further enable complex genetic manipulations like homologous recombination, multiple gene introductions, drug selection, and other strategies that may require extended culture. This significantly increases the value of these cells for future developmental studies. In addition, cultures beyond 50 passages that maintain a short doubling time and a pluripotent phenotype have not been previously reported. Beyond long-term culture, qiPSCs demonstrated the capability of clonal expansion after isolating individual cells in a 96-well format, providing the possibility of targeting genes of interest. Little is known about the maintenance of pluripotency and expansion of avian pluripotent cells. Therefore, we used methods based on the significant body of knowledge for mammalian pluripotent cells [48 –52]. The feeder-free culture system that supported qiPSC cultures for more than 50 passages without loss of the pluripotent phenotype was developed for human pluripotent cells and contains high levels of bFGF [53]. In the future, this system can be used to investigate individual factors and their role in maintaining pluripotency of avian stem cells. The qiPSC gene-targeted avian models will compliment important rodent models in disease and developmental gene function studies [27].
We report here the availability of chimera competent avian iPSCs that will greatly facilitate gene targeting and the insertion of genetic reporters. Future studies that generate cells with gene-specific and multiple promoters, inducers, and conditional expression systems in avian iPSCs are feasible, thus enhancing the research community's capabilities when it comes to directly observing cell migration and embryonic and fetal development in ova [54]. Since these qiPSCs were capable of clonal expansion after genetic modification, targeting genes of interest is potentially possible, which would facilitate research on gene function and signaling pathways underlying the development process in chimeric embryos. Further, avian iPSC derivatives such as neural cells should compliment mammalian cell transplant models for regenerative medicine [55]. In total, this unique source of avian iPSCs and derivative cells will provide biologists with multiple opportunities to enhance and expedite developmentally related discoveries.
Footnotes
Acknowledgments
The authors thank Dr. Jeanna L. Wilson (Department of Poultry Science, University of Georgia) for providing quail embryos, Anand Subramanian (Department of Animal and Dairy Science, University of Georgia) for immunostaining of neural marker MAP2, and Julie Nelson at the Center for Tropical and Emerging Global Diseases Flow Cytometry Facility. This work was jointly supported by the Bill and Melinda Gates Foundation and the Guangxi Scholarship Fund.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.