
Abstract
While defining the environment for human embryonic stem cell (hESC) culture on 2-dimensional (2D) surfaces has made rapid progress, the industrial-scale implementation of this technology will benefit from translating this knowledge into a 3-dimensional (3D) system, thus enabling better control, automation, and volumetric scale-up in bioreactors. The current study describes a system with defined conditions that are capable of supporting the long-term 2D culture of hESCs and the transposing of these conditions to 3D microcarrier (MC) cultures. Vitronectin (VN) and laminin (LN) were chosen as matrices for the long-term propagation of hESCs in a defined culture medium (STEMPRO®) for conventional 2D culture. Adsorption of these proteins onto 2D tissue culture polystyrene (TCPS) indicated that surface density saturation of 510 and 850 ng/cm2 for VN and LN, respectively, was attained above 20 μg/mL deposition solution concentration. Adsorption of these proteins onto spherical (97±10 μm), polystyrene MC followed a similar trend and coating surface densities of 450 and 650 ng/cm2 for VN and LN, respectively, were used to support hESC propagation. The long-term expansion of hESCs was equally successful on TCPS and MC, with consistently high expression (>90%) of pluripotent markers (OCT-4, MAB-84, and TRA-1-60) over 20 passages and maintenance of karyotypic normality. The average fold increase in cell numbers on VN-coated MC per serial passage was 8.5±1.0, which was similar to LN-coated MC (8.5±0.9). Embryoid body differentiation assays and teratoma formation confirmed that hESCs retained the ability to differentiate into lineages of all 3 germ layers, thus demonstrating the first translation to a fully defined MC-based environment for the expansion of hESCs.
Introduction
I
To date, much progress has already been made in the formulation of defined serum-free and feeder-free media for long-term hESC culture [5,6]. The commercially available options include STEMPRO® and mTeSR®1, both of which are widely used in hESC research [7,8]. Nevertheless, manufacturers of STEMPRO, mTeSR1, and other commercially available culture media invariably recommend the use of nondefined extracellular matrix (ECM) extracts, such as Matrigel™ or Geltrex™ as substrata for routine hESC culture. These nondefined matrices represent a major obstacle for potential uses of hESC in clinical applications, because a nondefined ECM like Matrigel (being a blend of proteins extracted from a murine sarcoma cell line [9]) exhibits a high degree of interbatch variability, in addition to being a potential source of xenogenic contamination. Several recent studies have identified and developed defined feeder-free substrata, capable of long-term hESC culture. Examples are laminin (LN), vitronectin (VN), fibronectin, and various peptides [10 –14], as well as characterizing the properties of Matrigel [15]. There is a concomitant need for the rigorous characterization of pluripotency and phenotypic stability of hESCs issued from long-term cultures on a defined substratum in defined medium. While a recent study demonstrated that hESCs can maintain their pluripotent state in a defined, serum-free medium (mTeSR1) on a defined substratum, VN, on tissue culture polystyrene (TCPS) [16], only one ECM protein was examined. The current study investigates 2 different ECM protein matrices VN and LN on TCPS, over a long duration of culture (20 serial passages) in a second defined medium (STEMPRO).
The industrial scale-up of hESC culture relies on leveraging successful 2-dimensional (2D) culture into a 3-dimensional (3D) culture environment, where microcarriers (MCs) offer large surface areas for hESC adhesion in bioreactors [17]. While the cultivation of hESCs on MC is a relatively new development in the stem cell field, the culture of cells on MC is a mature technology in the pharmaceutical industry. Several studies have demonstrated the advantages of hESC culture on MC [17 –21], which include higher cell titres being attained for a given volume of culture medium with MC, and obviating the need for enzymatic dissociation of cells during serial passage [17]. Moreover, it was shown that hESCs cultured on MC can be differentiated into a number of specific lineages, such as cardiomyocytes [17,19,22], endoderm [23], or neuronal and pancreatic cells [19].
A recent study demonstrated that LN-coated cellulose and cross-linked dextran MCs could support the long-term, stable propagation of pluripotent hESCs in conditioned medium [24]. While this prior study did not quantify LN surface density on MCs, the current study quantifies adsorbed VN and LN ECM proteins and extends the comparison of these matrices for the long-term propagation of hESCs in 2D and 3D culture environments in defined, serum-free medium.
Materials and Methods
Cells, culture media, MCs, ECM proteins, and reagents
The HES-3 (ES Cell International, Inc.) and H7 (WiCell Research Institute, Inc.) hESC lines were routinely maintained on Matrigel-coated TCPS plates using mouse embryonic fibroblasts conditioned medium (MEF-CM), as described in prior articles [25,26]. Unless otherwise stated, all culture media and supplements were purchased from Invitrogen and all reagents and chemicals were purchased from Sigma-Aldrich. All lab-ware consumables were sourced from Nunc. Polystyrene beads (Cat No. 7602B) with an average diameter of 97±10 μm, purchased from Thermo-Fisher Scientific, were utilized as MCs for hESC culture in this study. Human plasma VN (Cat No. CC080) >90% purity was purchased from Millipore and >95% purity mouse LN (Cat No. 23017-015), derived from Engelbreth-Holm-Swam sarcoma, was purchased from Invitrogen, in order to maintain consistency with our previous studies [16,24].
Coating TCPS and polystyrene MC with LN and VN
TCPS surfaces were coated with human plasma–purified VN and natural mouse LN to saturating surface densities (10 μg/mL and >20 μg/mL, respectively, Fig. 1) as described in Yap et al. [16]. Briefly, VN and LN solutions were prepared by diluting 1 mg/mL stock VN and LN solutions with sterile phosphate-buffered saline (PBS; Sigma P5493) to the required concentrations, which were used to coat TCPS organ culture dishes (OCDs; Becton Dickinson Biosciences) by incubating 300 μL of solution for 15 h at 4°C. OCDs were rinsed with PBS before using them as substrates for stem cell culture. The circular base of the OCDs presented a nominal cell culture area of 2.45 cm2.
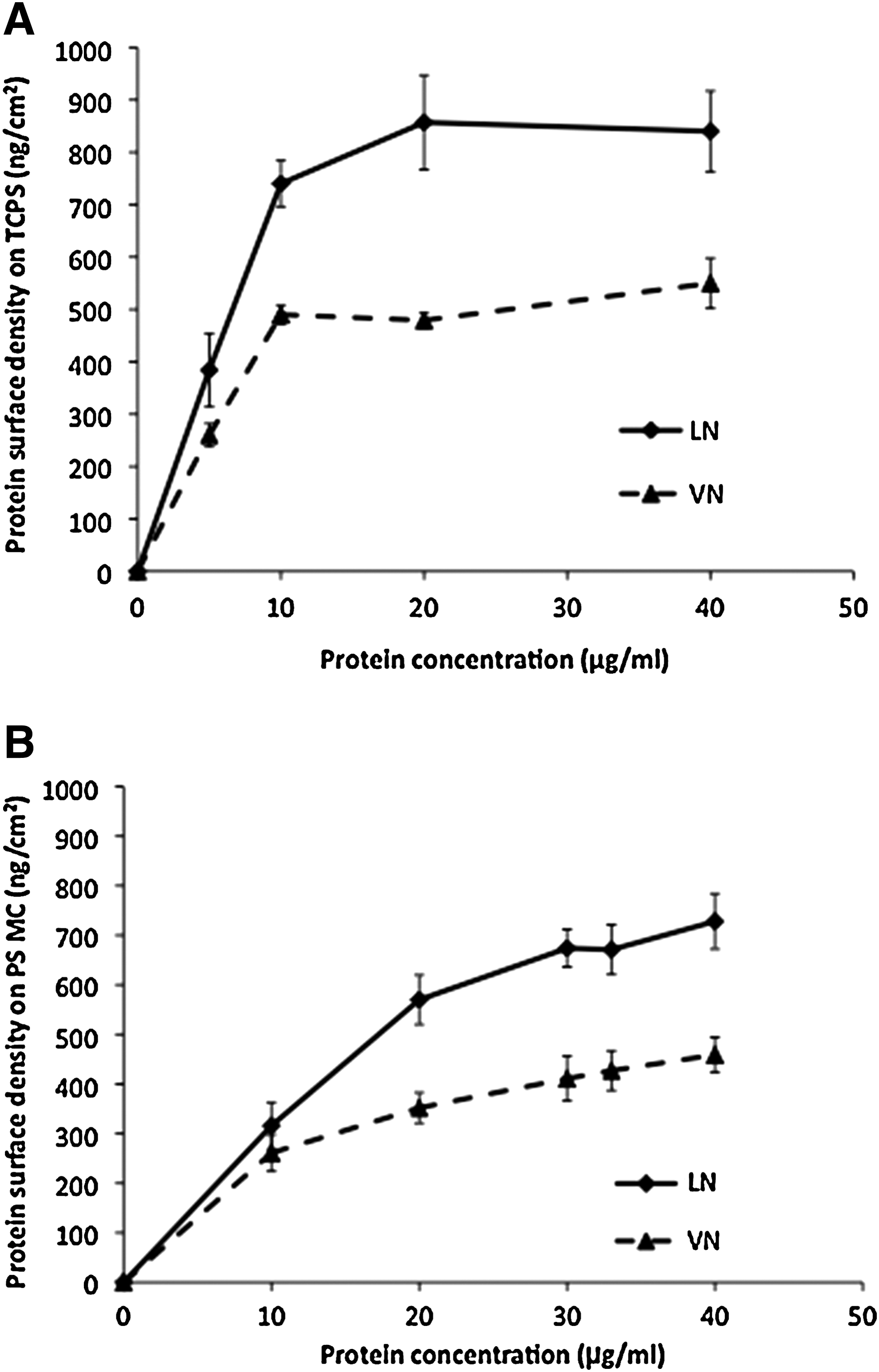
Adsorption of laminin (LN) and vitronectin (VN) on
Spherical MC, made from polystyrene (PS) cross-linked with 4%–8% divinylbenzene, was received as a 100 mg/mL aqueous suspension. These were washed 6 times with distilled water, followed by 5 times with absolute ethanol, and finally rinsed 3 times with distilled water, followed by 3 times in PBS. This MC suspension in PBS was sterilized by gamma irradiation (10 min, 10 kGray/h) exposure to a 60Co irradiator (Gammacell 220 Excel, Canada). For coating with protein, a 200 μL aliquot of 100 mg/mL MC suspension was placed in one well of a 24-well TCPS plate (Becton Dickinson Biosciences). To this, 380 μL PBS and 20 μL of 1 mg/mL stock VN or LN solution (final solution concentration of 33 μg/mL) were added. Following 15-h incubation at 4°C, the MC were briefly rinsed in PBS and immediately seeded with hESCs.
Surface characterization: quantification of VN and LN adsorbed on TCPS and MC by Bradford assay and Ponceau S
VN and LN adsorbed to TCPS were quantified by depletion from their depositing solution, whose concentration was measured using a modified Bradford assay [27,28]. TCPS substrates were incubated in solution concentrations of 0, 5, 10, 20, and 40 μg/mL for 15 h at 4°C, as described previously. After coating, the supernatant for each condition was measured and the protein surface density deposited on TCPS was calculated as described in Yap et al. [16].
VN and LN adsorbed to the surfaces of MC were quantified in a similar manner. Two hundred microliter aliquots of 100 mg/mL MC suspension was diluted with 400, 394, 388, 382, 380, and 376 μL PBS, to which were added 0, 6, 12, 18, 20, and 24 μL of 1 mg/mL stock VN or LN solution, respectively, in separate wells of a 24-well plate, for a total volume of 0.6 mL in each well. After incubation for 15 h at 4°C in the initial protein solution concentrations of 0, 10, 20, 30, 33, and 40 μg/mL, the final protein concentrations in the supernatants were quantified by the Bradford protein assay, thus yielding the total adsorbed protein mass. To differentiate protein adsorbed on the surface of the well from that adsorbed on the PS MC, both were stained by Ponceau S, by adding 0.8 mL of Ponceau S staining solution (Sigma-Aldrich) to each well containing protein-coated PS MC and incubated for 15 h at room temperature. After rinsing 5 times with 10% (v/v) acetic acid and 3 times with water, the 20 mg PS MC samples were placed in individual 1.5 mL Eppendorf tubes (Greiner Bio-one GmbH), and their Ponceau S stain was desorbed by incubating in 450 μL of 0.1 M NaOH for 20 min under gentle agitation. The Ponceau S stain adsorbed on the container walls was similarly desorbed by incubating in 450 μL of 0.1 M NaOH for 20 min under gentle agitation. Samples were run in duplicates, yielding four 200 μL aliquots per protein concentration for the PS MC and the container, separately. Each sample was placed in a flat-bottomed 96-well plate and neutralized with 15 μL of 50% acetic acid (J. T. Baker). Colorimetric absorption at 515 nm was used to quantify the Ponceau S stain (Supplementary Fig. S1A, B; Supplementary Data are available online at
2D culture of hESCs on LN- and VN-coated TCPS, with either conditioned medium or STEMPRO medium
hESCs were cultured on LN- or VN-coated OCD with either MEF-CM [26] or in STEMPRO (Invitrogen, Inc.) at 37°C and 5% CO2 for 7 days with daily change of MEF-CM or STEMPRO. For MEF-CM cultures, passaging was carried out by enzymatic dissociation of hESC colonies with collagenase IV (5 min at 37°C) at a passage ratio of 1:5 (200,000 cells per OCD). For STEMPRO cultures, passaging was carried out by enzymatic dissociation of hESC colonies with Accutase (3 min at 37°C), at a passage ratio of 1:20 (50,000 cells per OCD). Immediately before passaging hESC cultures were observed under a light stereomicroscope and colonies that appeared differentiated were removed by manual scarping and pipetting. For growth kinetics (after more than 20 passages under all 4 conditions), nuclei were counted daily for 7 days using a Nucleocounter® (Chemometec, Inc.) [17,18]. The seeding density in MEF-CM was twice that in STEMPRO (corresponding to 100,000 vs. 50,000 cells per OCD), since the same seeding density used in STEMPRO, gave suboptimal hESC proliferation rates in MEF-CM (data not shown). Fold expansion was calculated by dividing the cell density at day 7 by the inoculum cell density at day 0.
3D culture of hESCs on LN- and VN-coated PS MC with STEMPRO medium
For the initial transition from 2D to 3D culture, hESCs grown on Matrigel in MEF-CM were cultured in STEMPRO for at least one passage. Following enzymatic dissociation into small cell clumps with Accutase (3 min), the cells were seeded at 5×105 cells per well onto 20 mg of LN- or VN-coated PS MC (8 mg/mL), settled on the base of nonadherent 24-well culture plates. After 24-h incubation, 40 mg of the hESC-seeded PS MC (taken from 2 wells of the 24-well plate) was transferred into 5 mL of fresh culture medium and placed in one well of an ultralow attachment 6-well plate (Corning Cat No. 3471). Medium was refreshed daily (80%) and cultures were passaged after 7 days. Subsequent passaging relied on mechanical dissociation through gentle pipetting, producing relatively large-sized clumps of hESCs cultured on MC. These were seeded onto fresh LN- or VN-coated PS MC at a seeding density of 2×105 cells/mL per 40 mg of new, coated PS MC in 5 mL of medium, placed in a single well of an ultralow attachment 6-well plate.
Immediately after seeding, the culture plates were temporarily placed on an orbital shaker at 110 rpm in a 37°C/5% CO2 incubator for 2 h to promote contact between hESCs and MC. They were then cultivated under static conditions for 7 days before passaging. At each passage, cell numbers were measured by counting nuclei and the percentage of cells expressing pluripotent markers was assessed by flow cytometry [17,18]. Growth kinetics were measured for 2.5×105 hESCs seeded onto 10 mg of LN- or VN-coated PS MC in 1.5 mL of STEMPRO in triplicate wells of an ultralow attachment 24-well plate, with cell numbers measured daily by counting nuclei. Due to the 3D culture format, the growth kinetics data were more appropriately presented as cell titer per unit volume, rather than cell density per unit area for MC. Fold expansion was calculated by dividing the cell density at day 7 by the inoculum cell density at day 0.
Flow cytometry analyses of pluripotent markers OCT-4, TRA-1-60, and MAB-84
Expression levels of the intracellular transcription factor OCT-4 (Santa Cruz) and extracellular antigens MAB-84 [29] and TRA-1-60 (Millipore) were assessed by immunofluorescence using flow cytometry, as described previously [17,24].
Immunocytochemical staining for expression of pluripotent markers
Aggregates of hESCs on LN- and VN-coated MC were plated on corresponding LN- or VN-coated OCD and cultured for 2 days before fixing the cells with 4% paraformaldehyde, prior to staining with 4′,6-diamidino-2-phenylindole (DAPI) and mouse primary antibodies for TRA-1-60 or OCT-4 [24]. Alexa-fluor® 488- and 594-conjugated F(ab’)2 fragments of goat anti-mouse IgG (Invitrogen) were used as secondary antibodies. Immunofluorescence was visualized using an Axiovert 200 M fluorescence microscope (Carl Zeiss).
Embryoid body differentiation assay
Spontaneous differentiation of hESC MC cultures in vitro was carried out by embryoid body (EB) formation, following the protocol described by Chin et al. [25]. Briefly, following 7 days of differentiation, mechanically dissociated EBs were replated onto a gelatin-coated 6-cm tissue culture plate, on which they were cultured for 14 days. RNA was harvested from differentiated cells using an RNA extraction kit with DNase treatment (RNeasy Mini Kit, Cat No. 74104; Qiagen GmbH). cDNA, synthesized using Superscript II Reverse Transcriptase (Invitrogen), was used in subsequent quantitative reverse transcriptase–polymerase chain reaction (RT-PCR) containing Power SYBR Green PCR Master Mix (Applied Biosystems) with primers of the following genes, OCT-4, NANOG, AFP, GATA6, Hand1, Nkx2.5, PAX6, SOX1, and GAPDH (housekeeping gene), as previously described [24]. The fold change (relative expression unit) of each gene in EBs was referenced against the same gene prior to differentiation of hESCs. Immunostaining was carried out according to Chan et al. [30], to identify cells from the 3 primary germ layers. Briefly, differentiated hESCs were fixed with 4% paraformaldehyde for 15 min and blocked for 2 h in PBS buffer containing 0.1% Triton X-100, 10% goat serum, and 1% bovine serum albumin (BSA). The primary antibody was diluted in 1% BSA/PBS at the following concentrations: 1:400 for α-smooth muscle actin (SMA) (Sigma-Aldrich, Inc., Cat No. A5228), 1:1,000 for β-III tubulin (Millipore, Inc., Cat No. MAB1637), and 1:250 for α-fetoprotein (AFP) (Sigma-Aldrich, Inc., Cat No. A8452). Cells were then washed in 1% BSA/PBS and incubated in the dark with fluorescein isothiocyanate–conjugated secondary antibodies for 2 h at room temperature. Following another wash with 1% BSA/PBS, a fluorescent mounting medium with DAPI (Vectashield Cat No. H-1200) was added to cover the cells and incubated for 1 h before imaging immunofluorescence with an Axiovert 200M fluorescence microscope (Carl Zeiss).
Teratoma formation assay
To confirm the in vivo pluripotency of hESCs cultured on LN and VN, an intramuscular injection of cells was administered to SCID mice and the formed tumors were analyzed 10 weeks postinjection. Briefly, hESCs (cultured under various conditions for 16 passages) were enzymatically dissociated with Accutase. After passing hESCs cultured on MC through a 100 μm filter, the cells were resuspended in PBS and then injected into SCID mouse (5×106 cells per mice), as described previously [26]. After 10 weeks, mice were sacrificed and the tumors dissected, embedded in paraffin, sectioned, and stained with hematoxylin-eosin for histological examination.
Karyotype analysis
To assess chromosomal stability of hESCs cultured in the defined environments for 20 passages, karyotyping of 20 colonies using bromodeoxyuridine/colcemid was performed by the Cytogenetics Laboratory at the Department of Obstetrics and Gynaecology, Kandang Kerbau Women's and Children's Hospital, Singapore, as described previously [17].
Statistical analysis of data
All bar charts and graphs show standard deviations representing at least 3 measurements. Student's t-tests were carried out to determine whether observed differences were statistically significant between different experimental conditions (P<0.05 was considered as statistically significant).
Results
Quantification of LN and VN adsorbed on TCPS and PS MC by Bradford assay
It was important to determine the protein densities of LN and VN on 2D and 3D PS surfaces prior to choosing the coating concentrations for stable, long-term hESC culture. Figure 1A shows the adsorbed VN and LN surface density on TCPS: both increase with concentration of the depositing solution, rapidly reaching a saturation plateau above 10–20 μg/mL, with saturated surface densities for VN and LN on TCPS were quantified at 510±30 and 850±80 ng/cm2, respectively. Earlier work by Yap et al. [16] demonstrated that the threshold depositing solution concentration of VN required to achieve long-term stable hESC propagation is 5 μg/mL, corresponding to an adsorbed protein surface density on TCPS of 250 ng/cm2. In the current study, the concentration of VN coating solution was increased to 10 μg/mL and, in the absence of data identifying the threshold LN surface density required to support stable hESC propagation, the LN coating solution concentration was chosen as 20 μg/mL, which approaches the saturation surface density of adsorbed protein on TCPS.
Figure 1B shows the adsorbed protein surface density on MC increasing more slowly with the solution concentration than on TCPS (c.f. Fig. 1A.). For the 33 μg/mL solution concentrations used to coat MC for cell culture, deposited VN and LN surface densities are 400±50 and 650±40 ng/cm2, respectively, approaching their saturation values.
hESCs maintain long-term pluripotency in 2D culture on LN- and VN-coated TCPS
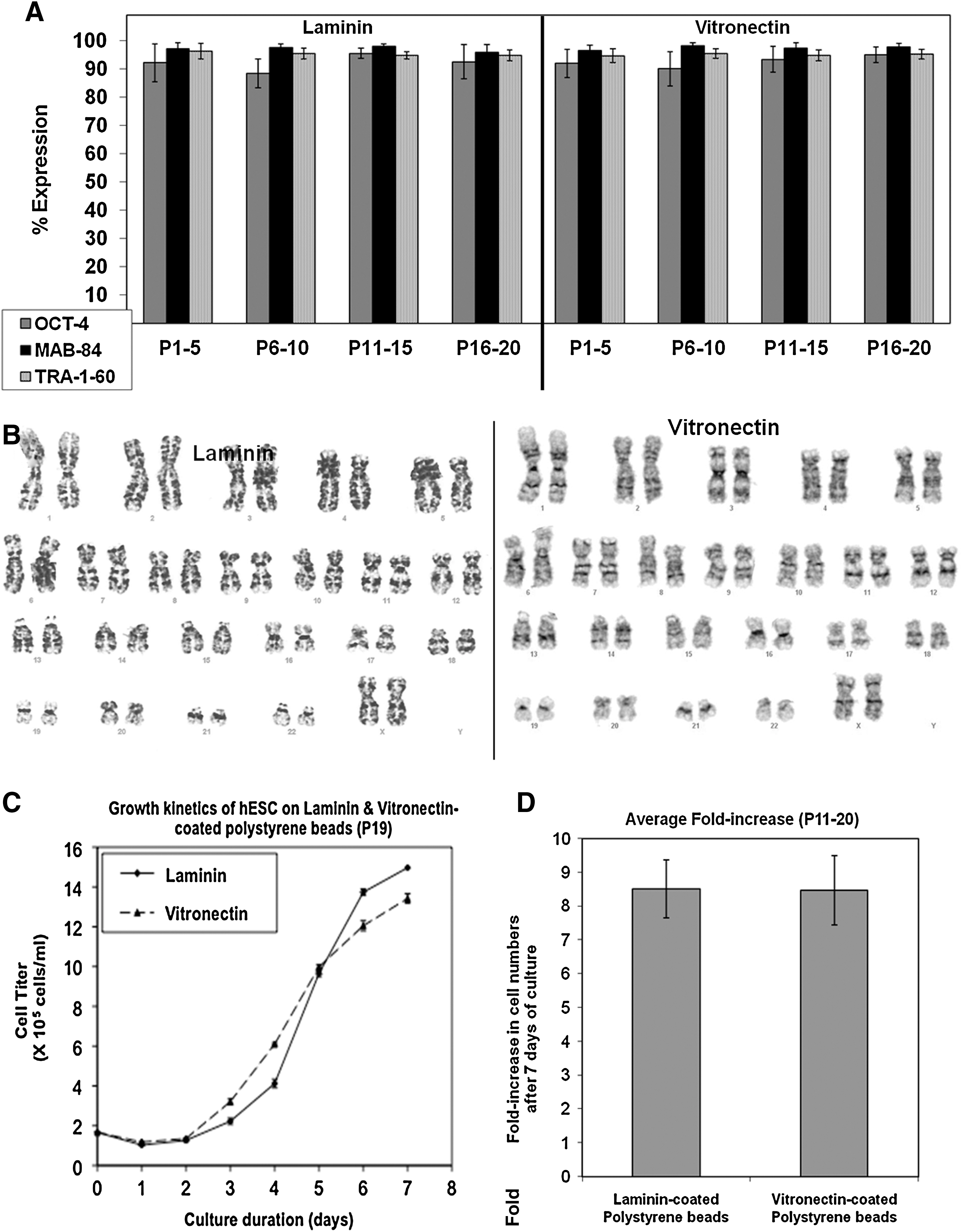
The initial phase of this study evaluated the ability of LN- and VN-coated 2D surfaces to support long-term hESC propagation in MEF-CM and STEMPRO. Figure 2 shows no distinct differences in cell morphology between hESCs cultured on LN and VN, in either MEF-CM or STEMPRO. Flow cytometry analysis revealed consistently high expression levels of pluripotent markers OCT-4, MAB-84, and TRA-1-60 over >20 passages on both LN- and VN-coated TCPS, in either CM or StemPro medium (Figs. 3A and 4A, respectively). These expression levels changed little from their initial values at passage 0: 97.9%, 99.7%, and 96.2% for OCT-4, MAB-84, and TRA-1-60, respectively. hESCs also displayed normal karyotype (46 XX) after 20 passages on either LN- or VN-coated TCPS, in both MEF-CM (Fig. 3B) and STEMPRO (Fig. 4B). hESC growth kinetics were similar when cultured on either LN or VN, notwithstanding differences observed between MEF-CM and STEMPRO culture media (Figs. 3C and 4D, respectively). Cells cultured on MEF-CM began their expansion after one day, while those cultured on STEMPRO exhibited a 4-day lag phase, which may be attributed to differences in enzymatic digestion procedures. For MEF-CM, relatively mild collagenase digestion of hESCs into large clumps was used. In contrast for STEMPRO, Accutase digests hESCs into single cells or small-sized clumps. Hence, the 4-day lag phase for STEMPRO could be generated by single cells or smaller clumps needing to migrate and form larger colonies, prior to undergoing optimal proliferation. For MEF-CM, the relatively large clumps generated by collagenase digestion are already optimal for proliferation, thus explaining the much shorter lag phase of one day.
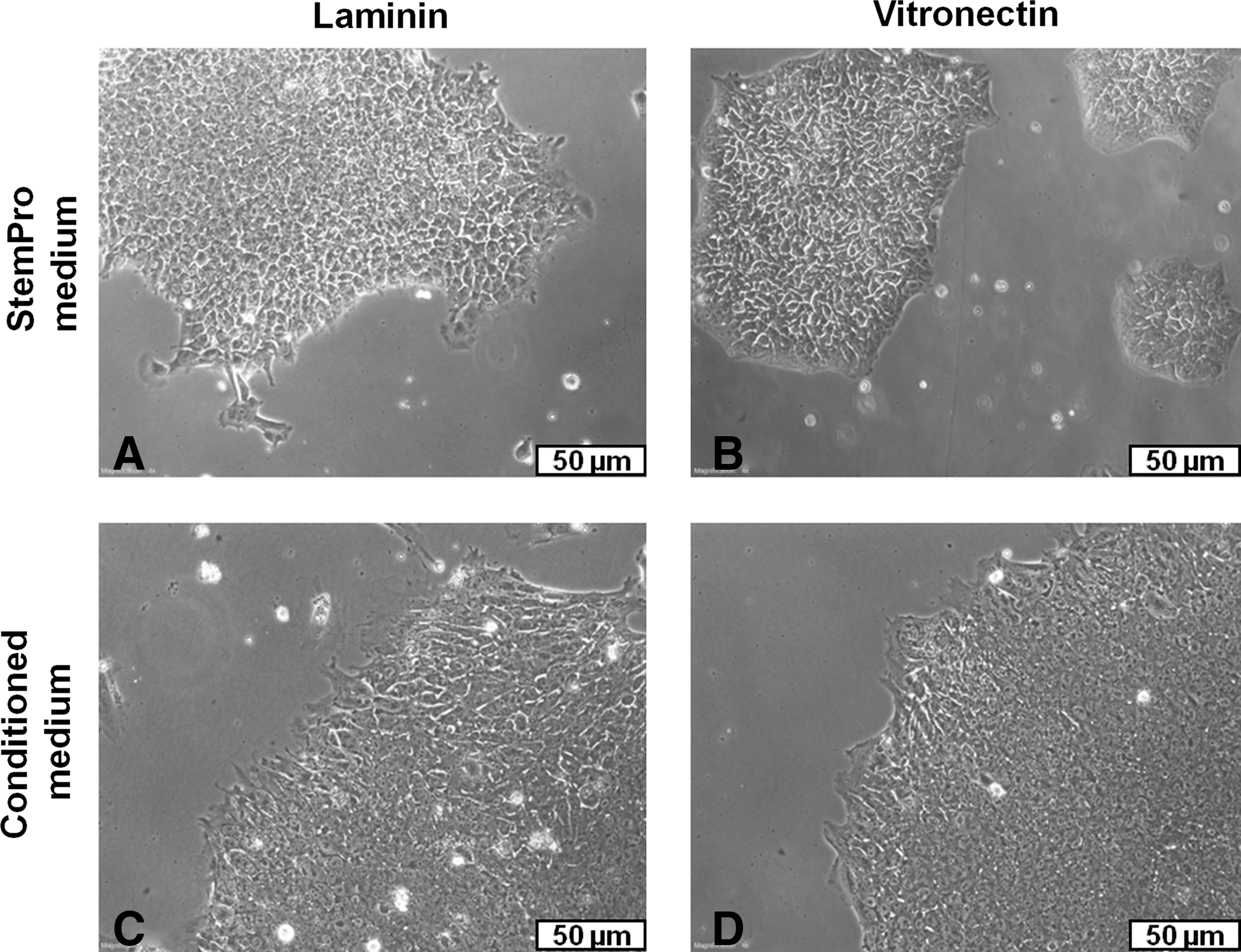
Phase-contrast microscopy images of human embryonic stem cells (hESCs; HES-3) in 2D culture on TCPS,

Long-term 2-dimensional (2D) culture of hESCs (HES-3) propagated in MEF-CM, on LN- and VN-coated TCPS.

Long-term 2D culture of hESCs (HES-3) in STEMPRO on LN- or VN-coated TCPS.
During their expansion phase, similar doubling times were observed for LN and VN of 22.2 and 25.6 h, respectively, on MEF-CM and 21.5 and 20.1 h, respectively, on STEMPRO. Final cell numbers yielded an approximately 16-fold increase on MEF-CM and a 27-fold increase on STEMPRO. hESCs expanded on both ECM proteins demonstrated pluripotency in their ability to form teratomas containing tissues from the ectoderm, mesoderm, and endoderm lineages (Fig. 4C).
hESCs also maintain long-term pluripotency in 3D culture on LN- and VN-coated PS MC
Having demonstrated that both ECMs could support the long-term 2D culture of hESCs in defined media, the study translated these successful culture conditions into 3D with MC coated with 33 μg/mL of LN and VN to support long-term hESC propagation in STEMPRO serum-free medium. As seen in Fig. 5A and B, hESCs cultured on LN- and VN-coated PS MC resulted in the formation of cell-MC aggregates that displayed similar morphology for both LN and VN coatings. Subsequent immunocytochemical staining revealed strong expression of OCT-4 and TRA-1-60 in hESCs cultured on PS MC coated with both LN (Fig. 5C, E) and VN (Fig. 5D, F), with corresponding DAPI stains of the nuclei shown in Fig. 5C1, E1, and Fig. 5D1, F1. This immunostaining data correlates with results of flow cytometry analysis, which showed consistently high expression of the pluripotent markers OCT-4, MAB-84, and TRA-1-60 over 20 passages (Fig. 6A and Supplementary Fig. S2). Karyotypic normality (46 XX) was also maintained after 20 passages on LN- and VN-coated PS MC (Fig. 6B). Growth kinetics of hESCs on LN- and VN-coated PS MC (Fig. 6C) showed similar behavior, with a lag phase of 2 days. During the expansion phase, from day 2 to day 5, the doubling times of hESC populations cultured on LN- and VN-coated PS MC were similar, at 24.6 and 25.0 h, respectively. From P11 to P20, hESCs cultured on LN-coated PS MC displayed an average 8.5±0.9-fold increase in cell numbers per serial passage. This was not significantly different (P>0.05) from the 8.5±1.0 value obtained on VN-coated PS MC (Fig. 6D). Although this demonstrates proof of concept for culturing hESCs on defined MC matrices, there is room for process optimization by tuning the MC dimensions and adjusting the composition of the cell culture medium.

Phase-contrast microscopy images of hESCs in 3D culture on PS MC coated with
Long-term 3D culture of hESCs (HES-3) on LN- and VN-coated PS MC in STEMPRO.
The pluripotency of hESCs expanded on LN- and VN-coated PS MC was further assessed by in vitro EB differentiation (Fig. 7) and in vivo teratoma formation (Fig. 8) assays. After 21 days' differentiation in EBs, quantitative RT-PCR analysis (Fig. 7A, E) showed that hESCs cultured on both LN- and VN-coated PS MC upregulated genes associated with the endoderm (AFP and GATA 6), mesoderm (Hand1 and Nkx 2.5), and ectoderm (Pax 6 and Sox 1) primary germ lineages, while downregulating their pluripotent markers (Nanog and OCT-4). This data was corroborated by positive immunostaining results for markers associated with the 3 embryonic germ layers (AFP, SMA, and β-III tubulin) on both LN (Fig. 7B–D) and VN (Fig. 7F–H) cultures. Dissected tissues from teratomas formed in SCID mice displayed all 3 characteristic germ layers for hESCs cultured on both LN- and VN-coated PS MC (Fig. 8A, B). Further validation was carried out with a second hESC line, H7, which maintained long-term pluripotency while expanding an average of 8.5±1.0-fold each passage on PS MC coated with both ECM proteins (Supplementary Fig. S3).

Spontaneous differentiation of hESCs cultured on LN- and VN-coated PS MC. Quantitative reverse transcriptase–polymerase chain reaction shows upregulation of genes associated with the formation of 3 germ layers and the corresponding downregulation of pluripotent genetic markers, for
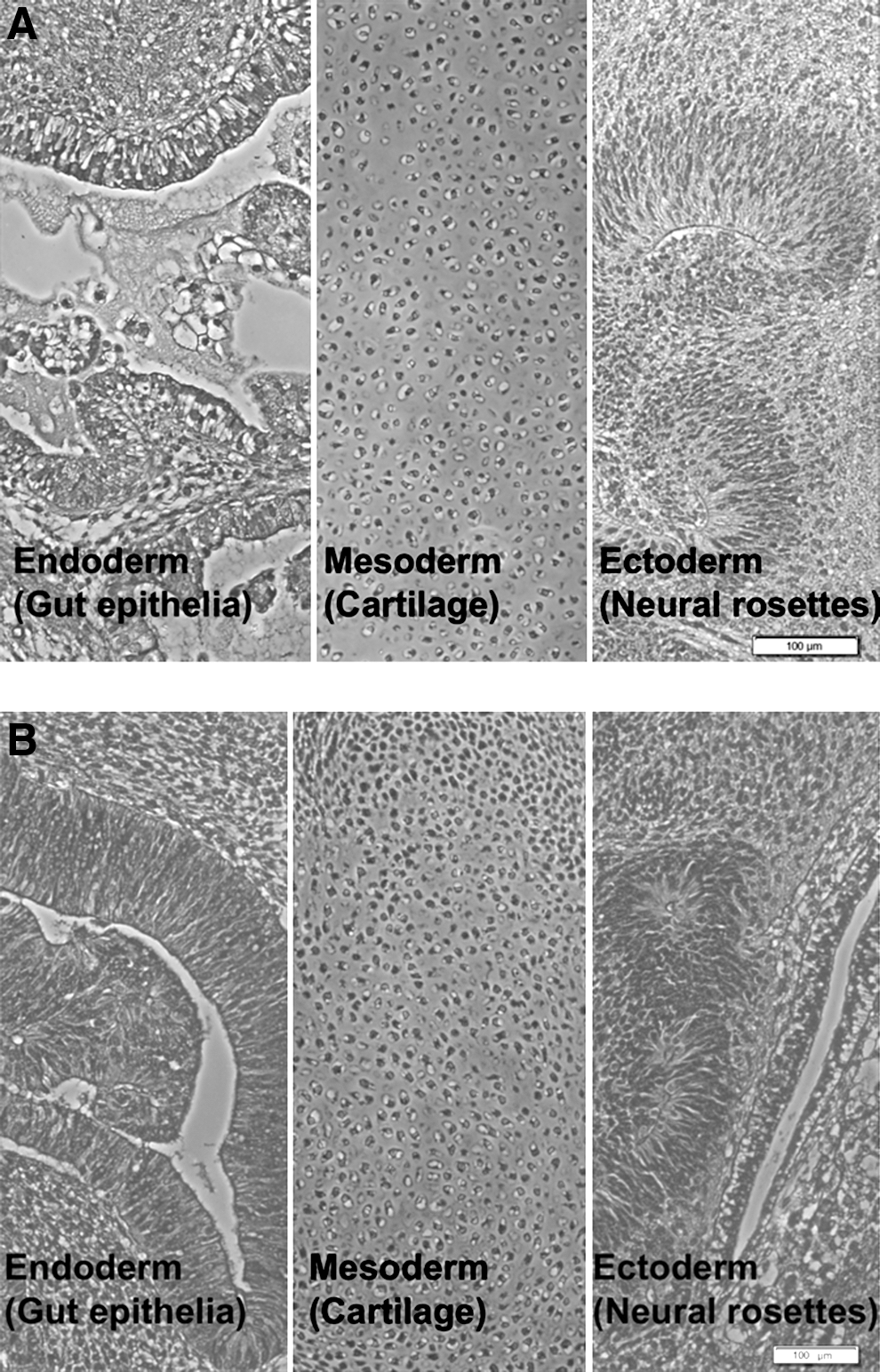
Hematoxylin-eosin staining of teratoma generated by hESCs from P16 in SCID mouse showing the 3 germ layers, gut epithelia (endoderm), cartilage (mesoderm), and neural rosettes (ectoderm) for
Discussion
Considerable progress has been made in recent years toward developing a defined environment for the long-term propagation of pluripotent hESCs [7,8]. Initial hESC culture relied on mitotically inactivated murine embryonic fibroblast feeder cells [31], gradually progressing to human-derived feeders [32], and eventually conditioned medium with Matrigel [33]. The current breakthrough in formulating serum-free, defined culture media, such as STEMPRO and mTeSR1 [7,8], has opened the possibility of replacing the routine use of nondefined ECM extracts, such as Matrigel [9] and Geltrex, with defined cell culture substrates, thus generating an entirely defined hESC expansion environment that is capable of expanding clinical-grade hESCs.
The current study examines the suitability of LN and VN ECM proteins as matrices for the long-term hESC expansion on 2D and 3D substrates. The selection of LN and VN stems from previous studies indicating the suitability of these adhesion-promoting proteins for long-term hESC culture [10,11,34,35]. Although prior studies by this group demonstrated the success of VN in 2D [16] and LN in 3D [24], no head-to-head comparison has been made between these matrices in 2D and 3D hESC cultures. Moreover, the current study is a first evaluation of these ECM matrices in a 3D environment in a defined, serum-free culture medium.
LN is a basement membrane glycoprotein that is known to mediate cell adhesion. Its interactions with polysaccharides [36] and proteins [37], including the activation of specific integrin receptors [38], play a key role in directing cell development, migration, and differentiation [39]. It is formed from the self-assembly of 3 chains into a cruciform structure [40,41] and exists in a number of genetic variants [42]. The current study implements a common form of murine LN (850 kDa), extracted from an Engelbreth-Holm-Swarm sarcoma [43]. Studies have also reported the suitability of human recombinant LN 511 for maintaining the pluripotency of both hESCs [10] and induced pluripotent stem cells (iPSCs) [44].
VN protein (75 kDa), found in both serum and the ECM, similarly mediates cell adhesion and spreading [45]. It has been demonstrated to be capable of supporting the long-term culture of both hESCs [10,16] and iPSCs [6,46]. The current study adsorbed human purified VN on a polymer substrate, as used in prior studies [16,47].
Besides comparing the performance of VN and LN as matrices for long-term hESC propagation in 2D culture, the current study also validates a transition from a 2D to 3D culture environment using similar PS substrates and cell culture media. Currently, the culture of hESCs in a 3D environment on MC is attracting much attention, due to its scale-up potential and ease of automation in bioreactors [17 –21]. Using MC allows for higher cell yields in a given volume of culture medium and bioreactors enable large batch processes to be run in an automated, controlled, and reliable manner. Additionally, the routine enzymatic dissociation of cells during serial passage is no longer required [17], which simplifies the entire culture process. Given that large quantities are required for clinical and nonclinical hESC applications, bioreactor culture with MC presents a viable and industrially scalable process to achieve this end.
Previous results [16] highlight the importance of determining the LN and VN protein surface density on 2D and 3D PS substrates prior to deciding coating solution concentrations suitable for the stable, long-term propagation of hESCs. LN and VN adsorbed on TCPS reach saturation above 10 μg/mL (Fig. 1A), while LN and VN adsorbed on PS MC approach saturation for deposition solution concentrations above 30 μg/mL (Fig. 1B). At 33 μg/mL solution concentration, VN surface density on PS MC exceeds the required threshold of 250 ng/cm2 for supporting long-term hESC expansion, as established by Yap et al. [16]. Similarly, the surface density of LN adsorbed on PS MC from 33 μg/mL is demonstrated by the current study to be capable of supporting long-term expansion of hESCs (Figs. 5 and 6).
The TCPS-adsorbed surface density of LN saturates at 850±80 ng/cm2, which corresponds to a uniform single layer of about 6.1 nm in thickness, calculated by assuming a protein density of 1.4 g/cm3 [48]. This may represent a quasi monolayer of LN molecules, oriented parallel to the substrate, without substantial aggregation. Given that aggregation of LN molecules in solution is generally mediated by divalent cations, either calcium [49,50] or magnesium [51], the absence of these ions from PBS would avoid accumulation of LN coating TCPS or PS MCs. Similarly, the saturated surface density of VN (510±30 ng/cm2) may be approximated to a uniform layer of thickness 3.6 nm, which is slightly less than the thickness of VN molecules aligned parallel to the polymer substrate [52]. The nanometer scale thickness of these ECM protein coatings contrasts sharply with Matrigel, which is reported to consist of a hydrogel film with a thickness of the order of 10 μm [15].
It has been reported that hydrophilic surfaces, with the exception of super-hydrophilic surfaces, generally adsorb more protein than hydrophobic surfaces [53,54]. This is attributed to protein molecules deforming as they bind to hydrophobic surfaces and thus yielding lower surface density of adsorbed proteins [53]. TCPS exhibits a water contact angle of 58°, while the bare PS surface of the MC is hydrophobic and thus presumed to have the wettability of a bare PS film, which approaches 90° [47]. This wettability difference may account at the adsorbed protein surface density on TCPS attaining saturation at lower deposition solution concentrations than on PS MC, as shown in Fig. 1.
The successful long-term propagation of hESCs on either LN or VN, in both 2D and 3D culture conditions (Figs. 4 and 6) yields similar performance parameters for both matrices. Fluorescence-activated cell sorting analysis revealed consistent high expression of pluripotent markers (OCT-4, MAB-84, and TRA-1-60) over 20 passages on both LN and VN. Karyotypic normality was maintained after 20 passages. The growth kinetics of hESCs cultured on VN and LN matrices were similar within 2D and 3D culture environments (Figs. 4D and 6C, respectively). An increase in growth rates was observed for the 3D versus the 2D culture environment (doubling times of 24 and 21 h, respectively). The fold increase in cell numbers, averaged over 10 passages (P11 to P20) in MC culture, did not differ significantly between LN and VN ECM protein matrices (Fig. 6D). Evidence of hESC pluripotency after expansion on both LN and VN was provided by positive results in the teratoma (Figs. 4C and 8) and EB differentiation assays (Fig. 7). Similarly, data for a second hESC line, H7, showed high expression of pluripotent markers and stable fold increase in cell numbers over 6 weeks for both LN- and VN-coated MC (Supplementary Fig. S3). These promising results are of interest when considering that hESC adhesion to LN and VN matrices is reported to be mediated by different subsets of integrin heterodimers expressed on the cell surface. Rodin et al. used antibody blocking to demonstrate that hESC adhesion to LN-coated surfaces is predominantly mediated by the α6β1 integrin heterodimer [44]. In contrast, VN-mediated adhesion of hESCs is mediated by the αVβ5 integrin heterodimer [11] and the proliferation on VN-coated surfaces is also dependent on β1 integrin, despite the β1 integrin not being essential for hESC adhesion on VN [46]. Interestingly, while Matrigel contains LN, it does not contain detectable VN [46].
From growth kinetics (Figs. 3C, 4D, and 6C), the fold increase over 7 days of culture was significantly higher for 2D culture (16-fold in MEF-CM and 27-fold in STEMPRO), when compared with 3D culture (≈8–9-fold). The lower fold-increase for MEF-CM, when compared to STEMPRO, is attributed to the higher seeding ratio, 1:5, which was used to mitigate the suboptimal proliferation rate observed for cells seeded at 1:20 in MEF-CM (data not shown). STEMPRO is optimally formulated for 2D hESC culture, which may explain why hESCs can grow well in STEMPRO at lower seeding densities than in MEF-CM. A prior study revealed higher fold expansion for defined versus conditioned medium [55], which is attributed to the defined culture media formulation being optimized for 2D hESC culture. Matrigel 2D cultures, in comparison, expanded 10-fold and 20-fold in MEF-CM and STEMPRO, respectively (data not shown).
Although data do not indicate whether this yield improvement is retained in the 3D culture environment, this loss in yield may be attributed to the hydrophobicity of the PS MC material. A prior study showed that hydrophobic surface properties can reduce the cell yields in 2D monolayer cultures [47]. The 8–9-fold increase over 7 days of culture also lies within a range observed on various MC made from more hydrophilic materials (Cytodex®, Tosoh®, and DE53®), coated with LN in conditioned medium [24]. The generation of large and compact hESC aggregates with PS MC (Fig. 5A, B) may be another factor contributing to reduced yield, due to limited access to nutrients and oxygen. Future studies will explore how varying the dimensions of the PS MC can affect the size and compactness of the hESC clumps, thus influencing hESC access to nutrients and oxygen, which may regulate their proliferation rate. A previous study indicated that high aspect ratio, rod-shaped MC are optimal for the culture of hESCs in a 3D environment [24]. This is probably because they generate less-compact cellular clumps. It might lend merit to examining rod-shaped PS MC, coated with ECM, for hESC culture in a defined medium.
The shorter 2-day lag phase for 3D culture (Fig. 6C), as compared with the 4 days observed in 2D culture (Fig. 4D), may arise from differences in passaging techniques. Enzymatic dissociation, used in 2D culture, generates either single cells or small cell clusters that need to reattach and spread before expanding. In contrast, the mechanical dissociation used for 3D culture generates hESC clumps that are capable of reattaching more quickly to new MC, hence reducing the lag phase. When Accutase was used to digest hESCs into single cells or small clumps, they attached with low efficiency to the LN- and VN-coated PS MC and grew very poorly, preventing the generation of any meaningful data with enzymatic digestion in 3D culture. By contrast, Accutase-digested single cells and small clumps adhere reasonably well on LN- and VN-coated 2D surfaces, probably due to cell sedimentation by gravity.
Recently, several groups have also developed suspension cultures of hESCs as pluripotent aggregates [56 –59]. Expansion of aggregates ranges from 6-fold between 4 and 7 days [56], to 25-fold over 10 days [58]. However, frequent dissociation to single cells is needed as aggregates larger than 0.5–1 mm tend to become necrotic and die [58,59] and some cell lines are also prone to spontaneous differentiation [56,57,59]. A group led by Skottman has also evaluated and developed their own xeno-free media for 2D culture of hESCs [60,61], while newer xeno-free products such as HEScGRO and Nutristem hESC SF are now in the market. It would be interesting to see whether these media are suitable for 3D MC or aggregate cultures of hESCs.
In summary, the current study demonstrates that LN and VN matrices are viable, generating similar performance for the long-term culture of hESCs in both 2D and 3D culture environments. Since industrial scale-up in bioreactors will require a degree of agitation to enable optimal circulation of oxygen and nutrients, future studies will examine the expansion of hESCs in agitated environments. With earlier work showing that agitation induces spontaneous differentiation of the HES-3 cell line [24,62], upcoming experiments will focus on mitigating this impact, through optimization of the hESC culture environment and minimizing shear stresses on the hESC/MC aggregates. Selection criteria for the MC and ECM will also have to consider maximizing hESC yield versus the economics of the stem cell bioprocess.
Footnotes
Acknowledgments
The authors thank the Joint Council Office of the Agency of Science and Technology (A*STAR) for funding this project and are grateful to Dr. Tan Heng Liang and Jasmin Seow for the teratoma assays and culture of the H7 cell line. We also thank Dr. Alan Lam for his critical review of this manuscript.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.