
Abstract
Stromal-derived factor (SDF)-1 is the main regulating factor for trafficking/homing of hematopoietic stem cells (HSC) to the bone marrow (BM). It is possible that this chemokine may also play a fundamental role in regulating the migration of HSC to several organs during extramedullary hematopoiesis. Because liver sinusoidal endothelial cells (LSEC) constitute an extramedullary niche for HSC, it is possible that these cells represent one of the main cellular sources of SDF-1 at the liver. Here, we show that LSEC express SDF-1 at the mRNA and protein level. Biological assays showed that conditioned medium from LSEC (LSEC-CM) stimulated the migration of BM progenitor lineage-negative (BM/Lin−) cells. This effect was significantly reduced by AMD3100, indicating that the SDF-1/CXCR4 axis is involved in the stimulatory migrating effect induced by LSEC-CM. Early localization of HSC in SDF-1–expressing LSEC microenvironment together with increased levels of this chemokine in hepatic homogenates was found in an experimental model of liver extramedullary hematopoiesis. Flow cytometry studies showed that LSEC express the CXCR4 receptor. Functional assays showed that activation of this receptor by SDF-1 stimulated the migration of LSEC and increased the expression of PECAM-1. Our findings suggest that LSEC through the production of SDF-1 may constitute a fundamental niche for regulation of HSC migration to the liver. To our knowledge, this is the first report showing that LSEC not only express and secrete SDF-1, but also its receptor CXCR4.
Introduction
T
During the adult life SDF-1 regulates not only the trafficking of HSC but also their homing and retention in BM [11]. It is known that HSC are attracted to an SDF-1 gradient generated at the BM [2]. There is evidence that the SDF-1/CXCR4 axis plays a critical role in directing homing/engraftment of HSC into BM after transplantation [15]. Other non-HSC expressing CXCR4 on their surfaces may also follow an SDF-1 gradient in other organs than BM [10,16,17]. Together, SDF-1 appears to be one of the most important chemoattractants for CXCR4+/HSC.
Under certain experimental and clinical conditions, extramedullary hematopoiesis results from migration of HSC from BM to some nonhematopoietic organs [18 –21]. The mechanisms by which circulating HSC are recruited to some organs are poorly understood; however, it may involve local chemotactic factors that promote the recruitment of these cells to them. It is possible that under certain conditions, circulating CXCR4+/HSC are chemoattracted to these organs which may express high levels of SDF-1. It may occur because the expression of SDF-1 in several tissues can be upregulated by various factors, such as proinflammatory molecules [22,23]. Regarding the liver, it is well known that it is one of the main organs where extramedullary hematopoiesis can take place [24]. The expression of SDF-1 has been reported by various liver cells [8,25,26]. Circulating CXCR4+/HSC may respond to an SDF-1 gradient generated at the liver under certain experimental and pathological conditions [26].
Previously, we have shown HSC migration and homing to specific liver sinusoidal endothelial cell (LSEC) microenvironment in an in vivo model of induced hepatic extramedullary hematopoiesis [24]. These findings suggest that LSEC may provide signals to regulate the migration and homing of hematopoietic cells to the liver, during extramedullary hematopoiesis [20,27]. In vitro, we have shown that cloned LSEC support the multilineage differentiation of murine hematopoietic cells HSC [19,28]. In this work, we hypothesized that LSEC could be a major source of SDF-1 in the liver, and they may constitute a fundamental niche for regulation of HSC migration and homing to the liver. We tested this hypothesis by examining for the expression and secretion of SDF-1 by LSEC and its possible role in HSC migration. We also examined for the expression of the SDF-1 receptor, CXCR4 in these cells.
Material and Methods
Reagents
Fluorescein isothiocyanate or phycoerythrin (PE)-conjugated rat monoclonal antibodies anti-mouse: CXCR4 (CD184), CD31 (PCAM-1), and CD106 (VCAM-1) were purchased from Becton Dickinson (San Diego). Recombinant murine SDF-1, tumor necrosis factor-α (TNF-α), and interleukin (IL)-6 were purchased from R&D Systems. G-CSF (Neupogen) was from Roche. AMD3100 (CXCR4 inhibitor) and Trizol were obtained from Sigma-Aldrich.
Animals
Female C57BL/6 mice (5–7 weeks old) were used as donors. Mice were obtained from the IVIC Laboratory Animal Center, and maintained on a standard laboratory diet and housed in a controlled environment. All animal experimentation was performed in accordance with institutional guidelines.
LSEC isolation and cloning
The isolation, cloning, and characterization of LSEC have been previously reported [24]. They express the EC markers flk-1, Von Willebrand factor (vWF), CD34, and VCAM-1 [24]. LSEC were maintained in Dulbecco's modified Eagle's medium (DMEM; Gibco)–10% fetal bovine serum (FBS; Sigma) and used for experiments after the cells reached 70% confluence.
Detection of SDF-1 transcript in LSEC by reverse transcriptase–polymerase chain reaction
Total RNA was isolated from monolayers of LSEC clones using a commercial reagent (TRIZOL). One microgram of the total RNA was reverse-transcribed in a total volume of 20 μL in buffer containing 250 mmol/L Tris–HCl (pH 8.3), 250 mmol/L KCl, 50 mmol/L MgCl2, 50 mmol/L dithiothreitol, 2.5 mmol spermidine, 10 mmol/L deoxynucleotide mixture, 100 pmol/L random hexamer oligonucleotides, and 15 U AMV reverse transcriptase (Promega). Polymerase chain reaction (PCR) amplification of the cDNA was then performed using specific oligonucleotides for the detection of SDF-1 and β-actin transcripts (Table 1). PCR conditions were 1 min of denaturation at 95°C, annealing at 61°C for 1 min, and extension at 72°C for 1 min for 35 cycles. Analysis of the PCR products was performed by comparing them with the predicted PCR fragment size after ethidium bromide staining of the PCR products separated by electrophoresis in a 1.5% agarose gel.
Production of conditioned media from LSEC
LSEC were grown to 90% confluence, and culture medium was replaced by fresh medium, and incubated for 48 h. After incubation, the conditioned media (LSEC-CM) was removed, centrifuged at 2,000 g for 15 min, and passed through a 0.2 mm filter before use.
Detection of SDF-1 in LSEC conditioned medium by enzyme-linked immunosorbent assay
LSEC-CM was analyzed by a specific enzyme-linked immunosorbent assay (ELISA) for murine SDF-1 (R&D Systems). A standard curve with a SDF-1-positive control was run in each assay. The sensitivity of these ELISAs was <0.015 ng/mL.
Isolation of BM lineage-negative (BM/Lin−) cells
Animals were sacrificed by cervical dislocation, and the femurs were aseptically removed. BM cells were obtained by flushing the femurs with DMEM medium. To obtain a population of primitive hematopoietic progenitors, BM/Lin− cells were isolated by immunomagnetic separation following the manufacturer's recommendations (Dynal Biotech ASA). In brief, a cocktail of lineage antibodies including those against Mac-1, Gr-1, B220, Ter119, and CD3 (BD) was submitted to immunomagnetic cell separation. Cellular negative fraction (BM/Lin−) was obtained and used for further assays.
Cell migration assay
We evaluated cell migration by using semi-porous polyester membrane (Transwell®, 24 well cell clusters; Corning, Inc.) of 5 μm and 8 μm pore sizes for BM/Lin− cells and for LSEC, respectively. BM/Lin− cells (1×104 cells) or LSEC (1×104 cells) were placed into the top of each transwell filter (100 μL serum-free medium). In some experiments BM/Lin− cells were pretreated with AMD3100 (25 μg/mL) for 45 min at 37°C, and after placed into the top of each transwell filter. Each Transwell containing the cells was carefully placed into 24 wells containing 400 μL serum-free medium alone or with either 20% LSEC-CM or SDF-1 (100 ng/mL) and incubated for 4 h at 37°C. After incubation, each Transwell was carefully removed and the plate was centrifuged, supernatants were discarded, and cells contained in the bottom of each well were fixed, stained with Wright-Giemsa (Himedia Labs), and counted in 9 high powered fields (hpf) per well by using an inverted microscope. Duplicate wells were used for each sample to give a total count of 18 hpf.
Induction of liver extramedullary hematopoiesis
Liver extramedullary hematopoiesis can be induced under several experimental conditions, and is the result of mobilization of BM HSC to this organ [24]. In this work we induced liver extramedullary hematopoiesis by injecting (i.p) a single dose of G-CSF (300 μg/Kg) in 0.1 mL phosphate-buffered saline (PBS). As control mice were given 0.1 mL of PBS. After 24 h, mice were sacrificed by cervical dislocation, and livers aseptically removed, fixed with 10% formalin, embedded in paraffin, sectioned into 3 μm slices, and stained with hematoxilin-eosin (H&E).
Immunohistochemical detection of SDF-1 and c-kit expression
Two-micrometer sections of paraffin-embedded mouse liver tissue were used for the detection of SDF-1 and c-kit. The sections were deparaffinized and incubated for 40 min with the antibody against SDF-1 (R&D Systems) or c-kit (Becton Dickinson), followed by incubation with biotin-labeled rabbit anti-mouse IgG (Dako Corp.) or biotin-labeled goat anti-rat IgG (Becton Dickinson), respectively, for 30 min at room temperature. The sections were then incubated with ABC complex (Vector Laboratories Inc.) and developed with DAB as substrate. They were counterstained in hematoxilin and covered with coverslips. As control, liver sections obtained from PBS injected mice were used.
Detection of SDF-1 in liver homogenates
Liver from mice injected with G-CSF and controls were minced and then homogenized by disruption between sterile glass slides in DMEM. The liver suspension was passed through nylon mesh, and centrifuged at 700 g for 15 min. The supernatants were collected and passed through a 0.2 μm filter before use. Liver homogenates were analyzed by ELISA for murine SDF-1.
Flow cytometry analysis of CXCR4 expression on LSEC
We evaluated the expression of CXCR4 on the surface of LSEC by flow cytometry. For this purpose, LSEC were grown to 90% confluence, the supernatant removed, and then the cells were harvested and incubated with PE-anti-CXCR4 mAb. Simultaneous negative control staining reactions were performed by incubating the cells with the PE labeled hamster IgG isotype. In some experiments, the effect of proinflammatory cytokines on CXCR4 expression by LSEC was evaluated by treating these cells with TNF-α (10 ng/mL) and IL-6 (20 ng/mL) for 18 h in serum free conditions. Data collection and analysis of the fluorescent intensities were made using a FACScalibur (Becton Dickinson). Ten thousand events were acquired and analyzed using the Cell Quest software program.
Effect of SDF-1 on PCAM-1 expression on LSEC
The effect of SDF-1 on the expression of PECAM-1 and VCAM-1 by LSEC was examined. For this purpose, LSEC were grown to 70% confluence, the supernatant removed, and then the cells were cultured in serum-free conditions alone or containing SDF-1 at 100 μg/mL for 18 h at 37°C in a 5% CO2 atmosphere. After incubation, cells were harvested and stained for analysis of PCAM-1 expression by flow cytometry, as described above.
In vitro adhesion assay of BM/Lin− cells to LSEC monolayers
BM/Lin− cells were (10×4 cells/well) were added onto untreated or SDF-1 treated LSEC monolayers. After incubation for 4 h at 37°C, the nonadherent cells were washed off, and the cell culture containing BM/Lin− cells adhered to LSEC monolayers were fixed. Nine fields from duplicate wells were photographed and rounded cells on LSEC monolayers were counted under an inverted microscope.
Statistical analysis
Results are reported as mean±standard error from duplicate or triplicate dishes in all assays. We tested the data from the experiments for statistical significance using the Student's t-test. A value of P<0.05 was considered significant. All experiments were repeated at least 3 times and have documented reproducibility.
Results
Isolation and cloning of LSEC
Isolation, cloning, characterization and culture of LSEC have been previously reported [24]. LSEC express the following EC markers: vWF, flk-1 (VEGF-R2) and CD34 (data not shown).
Expression of SDF-1 by LSEC
Based on results showing that LSEC constitute a liver hematopoietic niche, during extramedullary hematopoiesis [24], we investigated whether these cells express SDF-1. For this purpose, we examined the expression of SDF-1 in LSEC using reverse transcriptase-PCR. A cDNA band corresponding to the predicted size for SDF-1 message was obtained from RNA isolated from LSEC (Fig. 1A), demonstrating that these cells constitutively express SDF-1 transcripts. Similarly, SDF-1 protein was detected by ELISA in LSEC-CM (Fig. 1B). Together, these results show that LSEC express SDF-1 at the RNA and protein level.
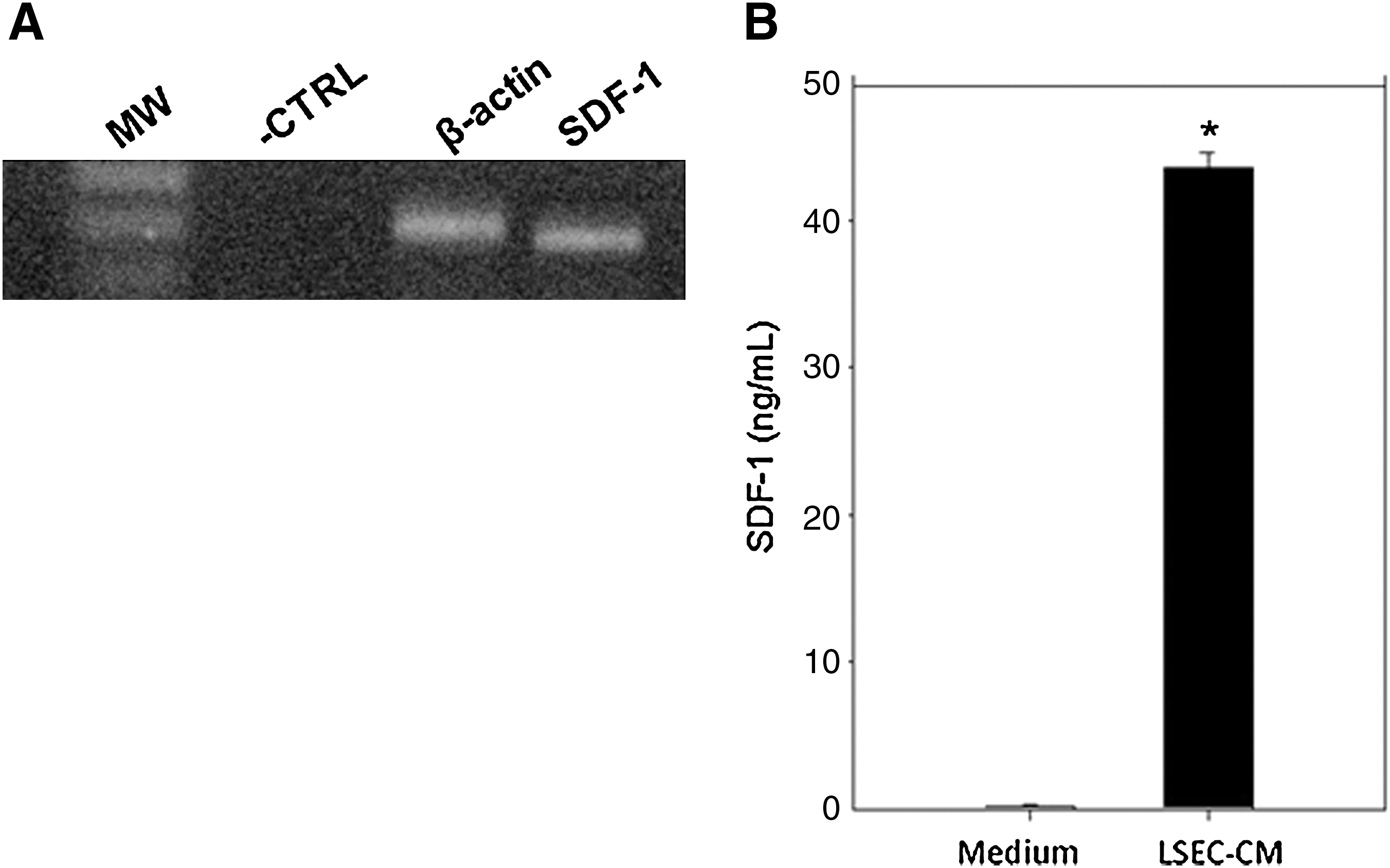
Expression of SDF-1 by LSEC.
Effect of LSEC conditioned media on the migration of BM/Lin- cells
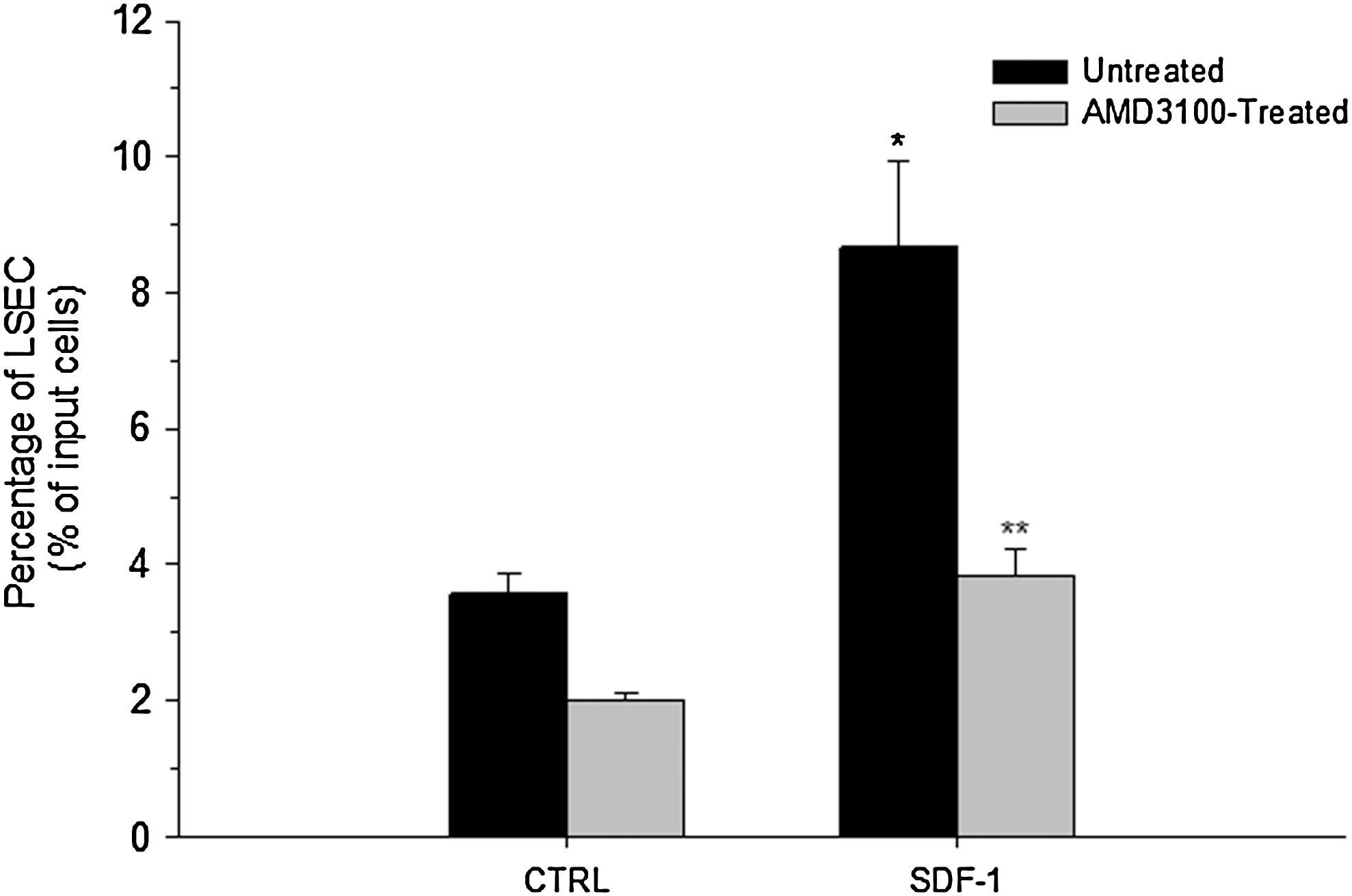
Based on the capacity of LSEC to secrete SDF-1 (Fig. 1B), we investigated the functionality of this chemokine by evaluating the effect of LSEC-CM on the migration of BM/Lin− cells using a transwell system. For this purpose, BM/Lin− cells (1×104 cells) were seeded on the top of transwells and placed in a 24-well plate containing serum-free medium alone (control) or supplemented with 20% LSEC-CM. A significant increase (P<0.05) in the migration of BM/Lin− cells was induced by LSEC-CM, compared with the control (Fig. 2). This effect was similar to that exerted by SDF-1 at 100 ng/mL (Fig. 2). To investigate whether the stimulatory effect of LSEC-CM on BM/Lin− cells migration was due to SDF-1, these cells were incubated with AMD3100 for 1 h, and they were seeded on the top of transwells, as above. AMD3100 inhibited>50% of the migration of BM/Lin− cells induced by LSEC-CM (Fig. 2). The specificity of the CXCR4 blockade by AMD3100 is showed by the significant inhibition (>50%) in the migration of BM/Lin− cells induced by SDF-1 (Fig. 2).
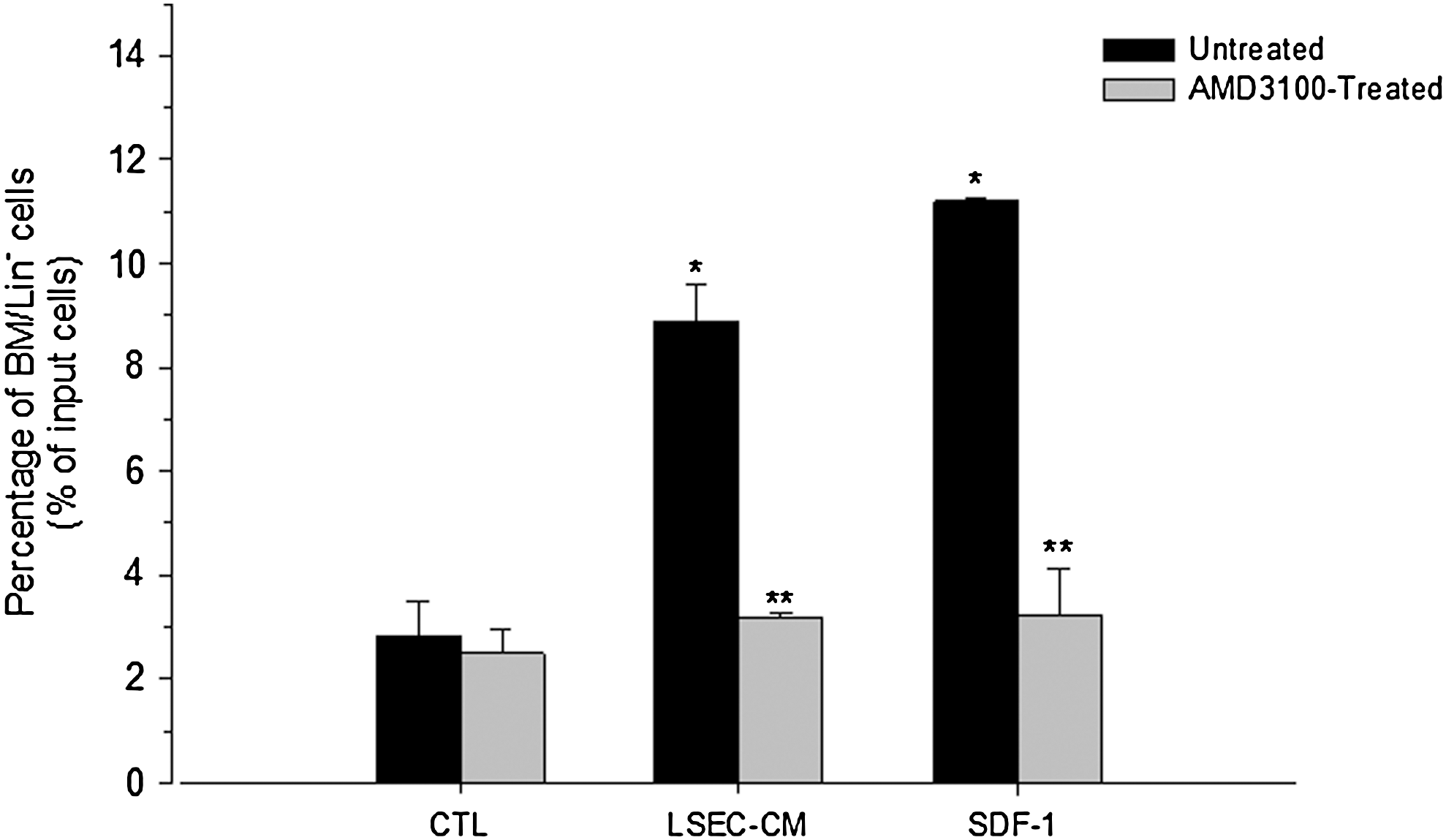
Effect of LSEC-CM on the migration of BM/Lin− cells. Untreated or AMD3100-treated BM/Lin− cells were seeded (10×4 cells/well) on the top of inserts (see material and methods) and placed in a 24-well plate containing serum-free medium alone (control, CTRL) or supplemented with 20% LSEC-CM or SDF-1 (100 ng/mL). After 4 h, the inserts were removed and the plate was centrifuged. Cells present in the bottom of each well were evidenced by Wright-Giemsa staining. The cells were counted in 5 high powered fields (hpf) per well by using an inverted microscope. Duplicate wells were used for each sample to give a total count of 10 hpf. LSEC-CM and SDF-1 induced a statistically significant increases in the migration of BM/Lin− cells as compared with the control (*P<0.05). AMD3100 treatment induced a statistically significant inhibition of the BM/Lin− cell migration induced by LSEC-CM and SDF-1 (**P<0.05). Data are shown as percentage of migrating cells from the input cell number (mean±SE of 3 independent experiments).
Expression of SDF-1 on LSEC microenvironments during liver extramedullary hematopoiesis
Because LSEC constitute an extramedullary niche for HSC (24,28), we investigated whether SDF-1 would be expressed on LSEC microenvironments during liver extramedullary hematopoiesis. For this purpose, we induced liver extramedullary hematopoiesis by injecting mice with G-CSF. Mice were sacrificed after 4 h, and histological evaluation of liver sections was performed. Immunohistochemical examination of liver section from these mice showed that SDF-1 was highly expressed in the LSEC territories (Fig. 3A). Interestingly, hematopoietic-like cells, inside the hepatic sinusoids, were observed in close proximity to the SDF-1–expressing LSEC microenvironment (Fig. 3A) suggesting a possible relationship between the presence of these cells and the expression of SDF-1 by LSEC. High levels of SDF-1 were detected by ELISA in liver homogenates from these animals (Fig. 3B).
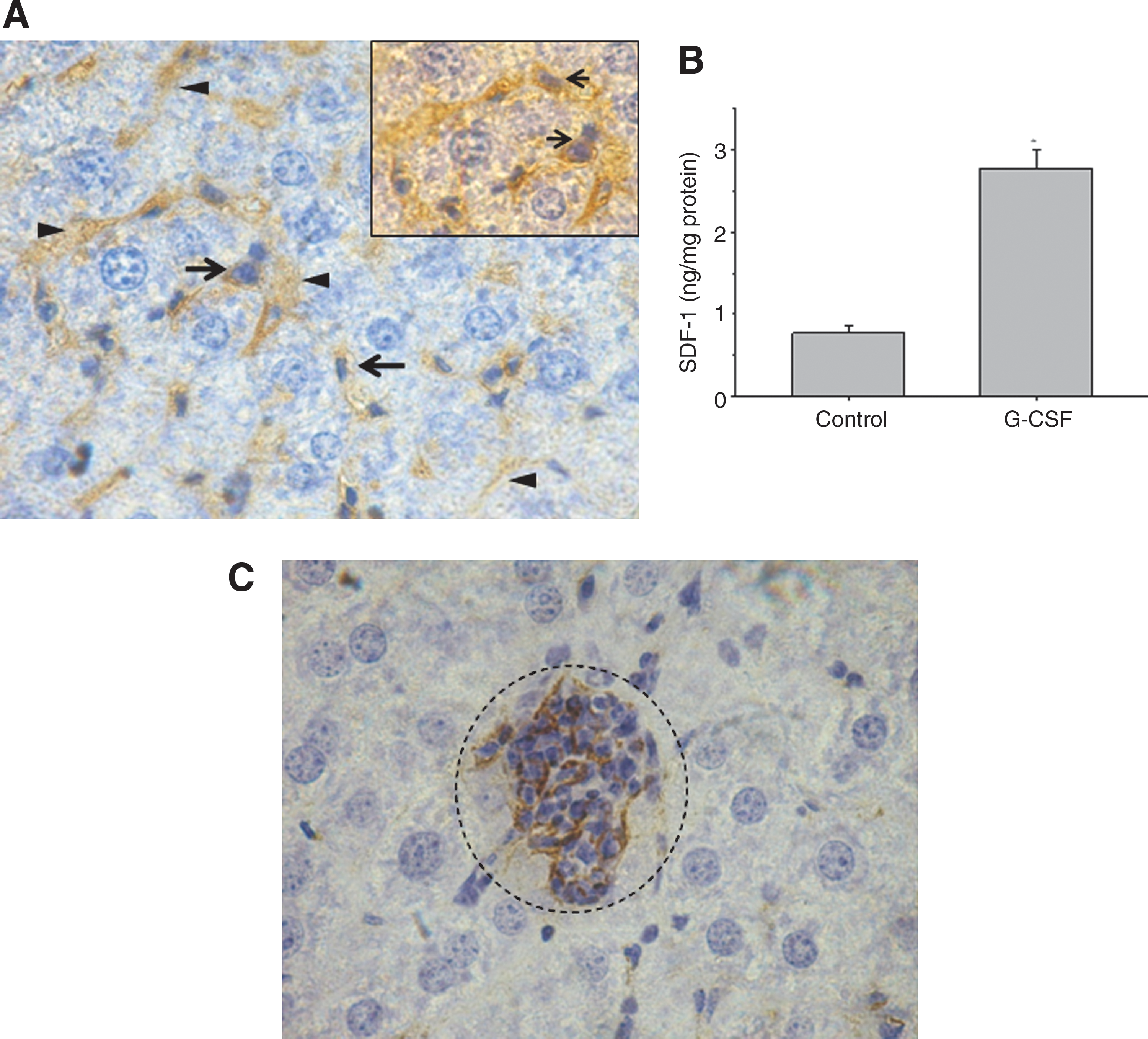
Detection of SDF-1+-LSEC and c-kit+-hematopoietic cells in a liver extramedullary hematopoiesis model. An experimental model of liver extramedullary hematopoiesis was induced by injecting mice with a single intraperitoneal dose of G-CSF. After 24 h mice were sacrificed and evaluation of liver sections was performed.
Detection of HSC in the hepatic sinusoids during liver extramedullary hematopoiesis
The presence of HSC inside the hepatic sinusoids were confirmed by detecting cells expressing the primitive hematopoietic stem cell marker, c-kit, in close contact with LSEC (Fig. 3C).
Expression of CXCR4 on LSEC
Based on evidence showing the expression of CXCR4 on EC, we examined the expression of this receptor on LSEC surface by flow cytometry. LSEC constitutively expressed high levels of CXCR4 (Fig. 4). We evaluated whether the endothelial cell-activating cytokines, IL-6 and TNF-α, could regulate the expression of CXCR4 on LSEC. IL-6 (20 ng/mL), but not TNF-α (10 ng/mL), induced an increase in CXCR4 expression on LSEC, after 18 h of treatment (Fig. 5). Further experiments showed that G-CSF did not regulate the expression of CXCR4 (not shown).
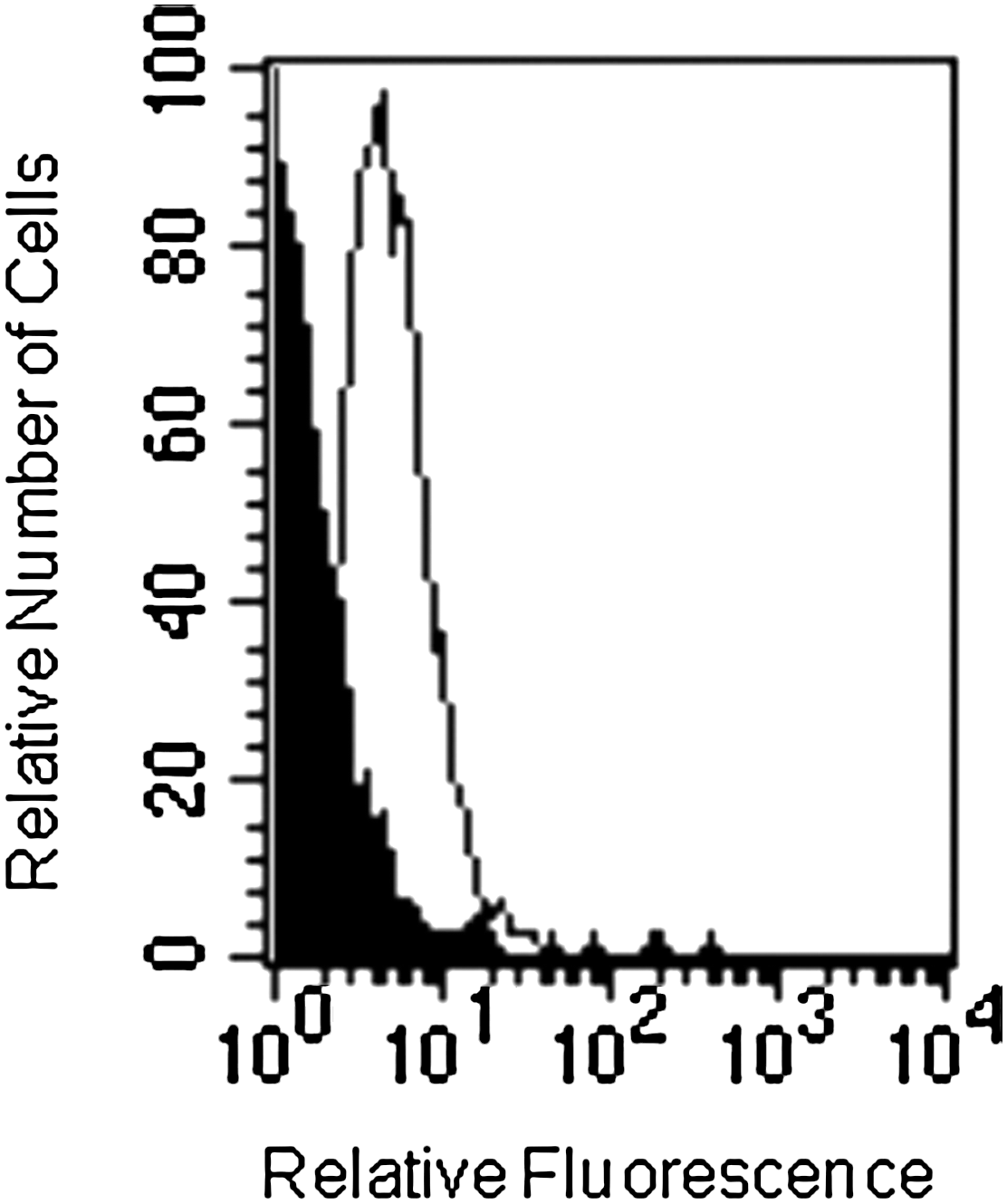
CXCR4 expression on LSEC. Representative flow cytometric histograms showing CXCR4 expression (thin line) on LSEC. Negative controls were stained with the respective isotype (black profile). Results are representative of at least 3 independent experiments, all of which had similar results.
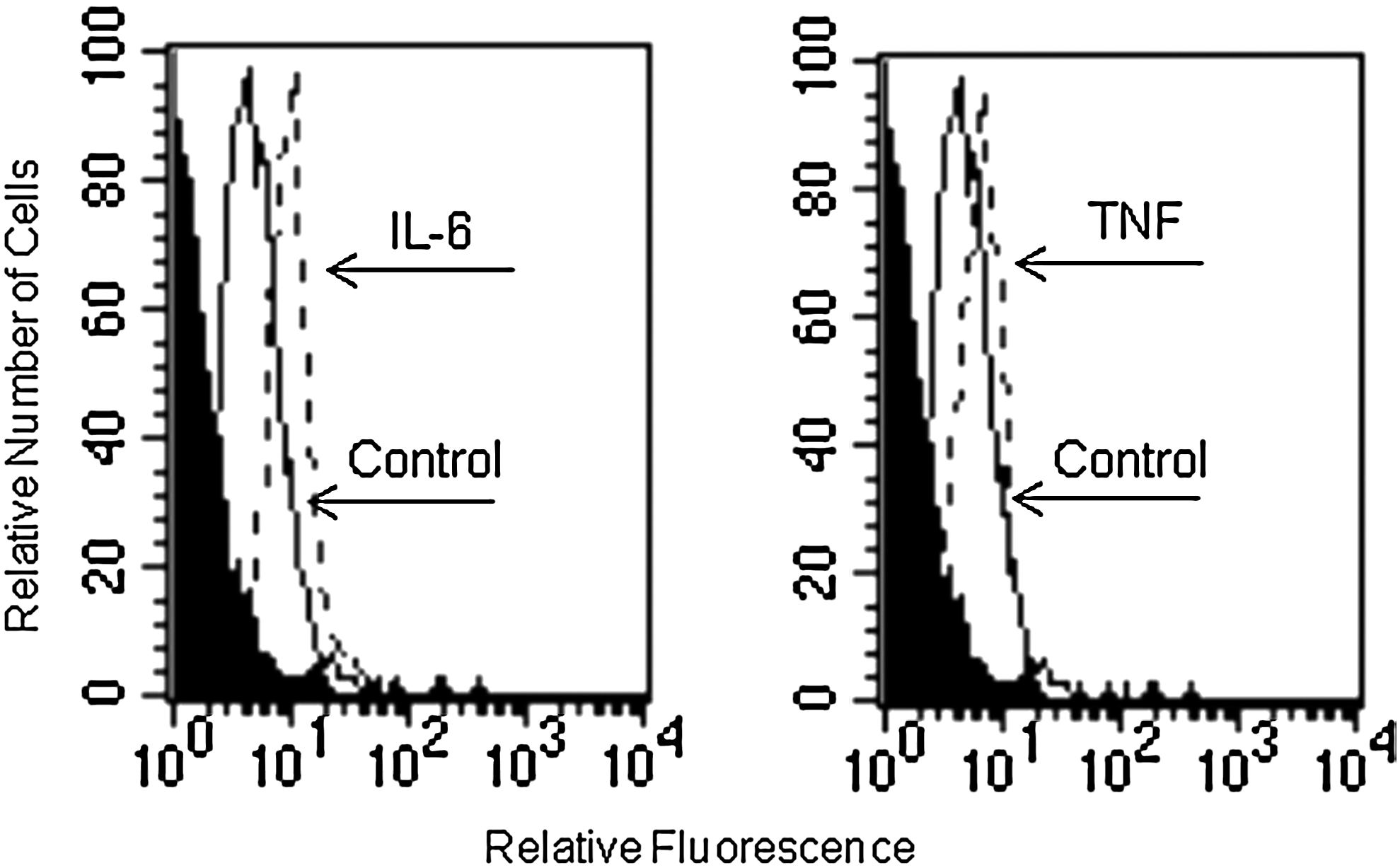
Effect of IL-6 and TNF-α on CXCR4 surface expression in LSEC. LSEC cells were cultured for 18 h with medium alone or in the presence of IL-6 (20 ng/mL) or TNF-α (10 ng/mL), and CXCR4 expression was analyzed by flow cytometry. Negative controls were stained with the respective isotype (black profile). Results are representative of at least 3 independent experiments, all of which had similar results.
Effect of SDF-1 on LSEC migration
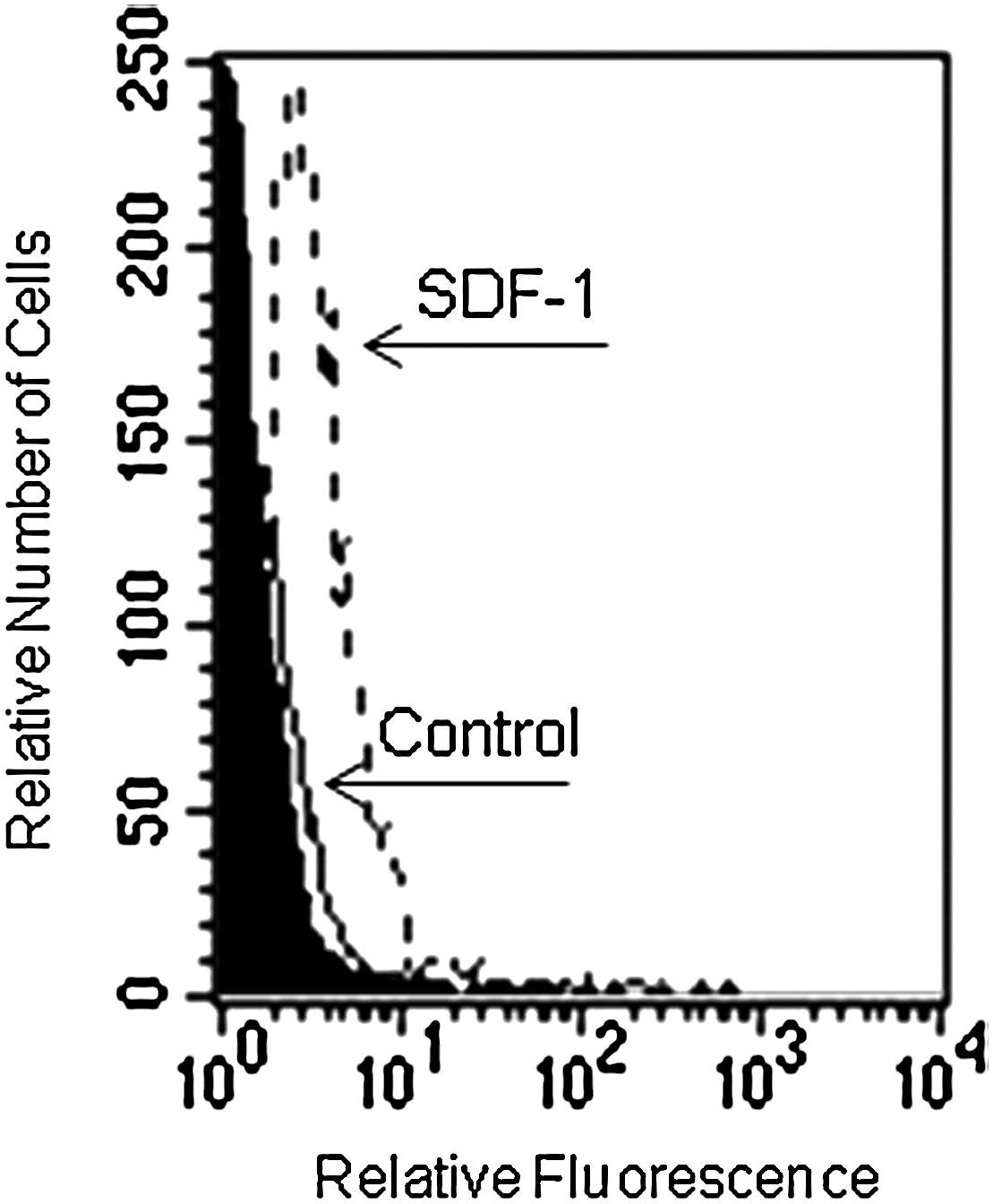
Based on the CXCR4 expression by LSEC, we investigated whether ligation of this receptor with SDF-1 might stimulate some biological functions in these cells. For this purpose, we evaluated the effect of SDF-1 on the migration of LSEC. LSEC were seeded on the top of transwells, and they were placed in a 24 well plate containing serum-free medium with or without SDF-1 at 100 ng/mL for 4 h. SDF-1 induced a significant stimulatory effect on the migration of LSEC, compared with the control (P<0.05) (Fig. 6). Blocking experiments using AMD3100 showed a significant reduction of LSEC migration, indicating that LSEC migration induced by SDF-1 was mediated by the CXCR4 receptor (Fig. 6).
Effect of SDF-1 on LSEC migration. Untreated or AMD3100-treated LSEC cells (10×4 cells/well) were seeded on the top of inserts (see material and methods) and placed in a 24-well plate containing serum-free medium alone (control, CTRL) or containing SDF-1 (100 ng/mL). After 4 h, the inserts were removed and the plate was centrifuged. LSEC present in the bottom of each well were evidenced by Wright Giemsa staining. The cells were counted in 5 high powered fields (hpf) per well by using an inverted microscope. Duplicate wells were used for each sample to give a total count of 10 hpf. SDF-1 induced a statistically significant increases in the migration of LSEC as compared with the control (*P<0.05). AMD3100 treatment induced a statistically significant inhibition of the LSEC migration induced by SDF-1 (**P<0.001). Data are shown as percentage of migrating cells from the input cell number (mean±SE of 3 independent experiments).
Effect of SDF-1 on the expression of PCAM-1 and VCAM-1 on LSEC
Because there are reports showing that SDF-1 may induce EC activation [17], we evaluated the effect of SDF-1 on the expression of PCAM-1 by LSEC. For this purpose, LSEC were treated with SDF-1 at 100 to ng/mL for 24 h, and the expression of PCAM-1 was evaluated by flow cytometry. SDF-1 induced a significant increase in PCAM-1 expression on LSEC surface (Fig. 7).
Effect of SDF-1 on the expression of PECAM-1 and VCAM-1 on LSEC. LSEC were cultured for 18 h in medium alone or containing SDF-1 (100 ng/mL), and then stained with conjugated rat mAb against mouse PCAM-1 or with an isotype control (black profile). Histograms are representative of at least 3 independent experiments, all of which had similar results.
Adhesion of BM/Lin- cells to LSEC monolayers treated with SDF-1
We were interested in evaluating whether the capacity of SDF-1 to up-regulate the expression of adhesion molecules on LSEC might increase the adhesion of BM/Lin- cells to these cells. For this purpose we investigated the adhesion of BM/Lin− cells to LSEC monolayers treated with SDF-1. Treatment of LSEC monolayers with SDF-1 (at 100 ng/mL) by 18 h induced a significant increase in the adhesion of BM/Lin− cells, compared with untreated LSEC monolayers (>4-fold increase over the control) (Fig. 8).

Effect of SDF-1 on the adhesion of BM/Lin− cells to LSEC monolayers. BM/Lin− cells were seeded (10×4 cells) on untreated or SDF-1 treated LSEC monolayers. After 4 h, the nonadherent cells were removed and rounded cells were counted in 9 different fields using a microscope. SDF−1 induced a significant increases in the number of BM/Lin− cells adhered to LSEC monolayers with respect to the control (*P<0.05). Data are shown as percentage of BM/Lin− adhered cells from the input cell number (mean±SE of 3 independent experiments).
Discussion
It is known that under certain clinical and experimental conditions, the liver is one of the main organs where extramedullary hematopoiesis occurs [20,21,24]. Because SDF-1 is the main regulating factor for trafficking/homing of HSC to the BM [1,15], it is possible that this chemokine may also regulate the migration of these cells to the liver during extramedullary hematopoiesis. However, the hepatic cells responsible for the production of this chemokine during extramedullary hematopoiesis are not completely defined. Based on previous results indicating that the LSEC microenvironment constitutes an extramedullary niche for HSC [24], we hypothesized that these cells may provide signals, through the production of SDF-1, to attract circulating HSC to the liver.
In this work, we show that LSEC constitutively express SDF-1 at the RNA level. The fact that SDF-1 was detected by ELISA in conditioned medium from LSEC indicates that these cells not only express it at the mRNA level, but also secrete this chemokine. These results agree with previous reports showing the expression and production of SDF-1 by EC from other sources [11,13,14]. The capacity of LSEC-CM to stimulate the migration of hematopoietic progenitor cells (BM/Lin-) suggest that SDF-1 produced by LSEC is functional. Moreover, the significant inhibition of the migration of BM/Lin- by the CXCR4 antagonist AMD3100, indicates that the effect of LSEC-CM is mediated through the SDF-1/CXCR4 axis.
An important question derived by our in vitro findings, is whether the capacity of LSEC to produce SDF-1 may also serve, in vivo, as an attracting signal for circulating HSC when they HSC are mobilized or under certain pathological conditions. Based on in vivo evidence that the LSEC serve as a specific hematopoietic niche during experimental extramedullary hematopoiesis [24], it is possible that these cells represent the main microenvironment regulating migration and homing of HSC to the liver through the production of SDF-1. This possibility is supported by the early and specific expression of SDF-1 on the LSEC, and not on other hepatic cells, in livers of mice injected with G-CSF. Our results differ from those showing the expression of SDF-1 in the liver by stellate cells and biliary epithelium [8,16,25,26,29]. However, these studies reported the expression of SDF-1 by these cells under inflammatory conditions and in chronic disease. Our results showing c-kit+-HSC, located in close proximity to the SDF-1–expressing LSEC suggest a possible relationship between the presence of HSC and the expression of this chemokine by LSEC. The possible involvement of SDF-1 during extramedullary hematopoiesis is supported, in our work, by the high levels of this chemokine found in liver homogenates from mice injected with G-CSF. Together, these results suggest that during experimental liver extramedullary hematopoiesis, the LSEC may constitute a specific hematopoietic niche due to their capacity to produce SDF-1, which promote the attraction, migration and adhesion of HSC to this organ.
The expression of CXCR4 has been shown in HSC and cells of nonhematogenous origin, such as EC [9,11,29]. Our results show that LSEC constitutively express functional CXCR4 receptor. Ligation of CXCR4 by SDF-1 induced LSEC migration indicating that this receptor is functional in these cells. These results agree with those showing the expression of functional CXCR4 on EC from other sources [30,31]. The constitutive expression of both SDF-1 and CXCR4 by LSEC suggest that this chemokine may play an autocrine role in these cells. An angiogenic effect of SDF-1 has been reported during liver regeneration after hepatic tissue damage [30,32]. It is possible that the SDF-1/CXCR4 axis may exert in vivo a proliferation and migration effect on LSEC. Our in vitro results also show that the expression of CXCR4 is up-regulated by IL-6, suggesting that proinflammatory cytokines may regulate the expression of this receptor on these cells. There is in vivo evidence showing that the expression of CXCR4 may be regulated during inflammatory conditions [33,34]. Regarding the liver, the increased expression of CXCR4 has been reported in inflammatory hepatic diseases [35,36]. Up-regulation of CXCR4 has been also reported in other cells including hematopoietic progenitor cells [9,37].
There is evidence showing that SDF-1 may increase the adhesion of cells bearing CXCR4 [17,37,38]. Likewise, it has been shown that SDF-1 may increase, in vivo and in vitro, the adhesion of inflammatory cells to the endothelium [27,39]. These results can be explained by the capacity of SDF-1 to upregulate the expression of various adhesion molecules on various cells [37,38,40]. Here, we show that SDF-1 increases the expression of PECAM-1 on LSEC, indicating that this chemokine may regulate the expression of adhesion molecules on these cells. It has been well established that PECAM-1 plays a role in regulating cell adhesion, migration and activation in cells of hematopoietic origin. Additionally, there is evidence that PECAM-1 plays a role in the regulation of HSC migration to the chemokine CXCL12 [41]. By upregulating the express ion of PCAM-1, SDF-1 may increase the adhesion of cells to LSEC. This possibility was confirmed by showing an increased adhesion of BM/Lin− to SDF-1-treated LSEC monolayers. It is possible that during liver extramedullary hematopoiesis, SDF-1 produced by LSEC not only may attract circulating CXCR4+/HSC but also could increase the adhesion of these cells to the LSEC microenvironments.
We have previously shown that during liver extramedullary hematopoiesis, hematopoietic foci are present only in the hepatic sinusoids in those zones covered by LSEC [21,24].There is evidence showing that the SDF-1/CXCR4 axis may be implicated in stress-induced stem cells trafficking in vivo, and this mechanism may be implicated in HSC migration into the liver under stress conditions [8,42,43]. The capacity of SDF-1 to induce migration of HSC to the liver has been shown by injecting this chemokine into the liver parenchyma [8]. Here, we show the expression of SDF-1 on LSEC microenvironments during an experimental model of liver extramedullary hematopoiesis. Interestingly, hematopoietic-like cells were located on the LSEC microenvironments expressing SDF-1, which was associated with the presence of high levels of SDF-1 in liver homogenates. These results suggest that SDF-1 produced by LSEC may constitute one of the main signals to attract HSC to the liver during extramedullary hematopoiesis. Based on these results, we propose that LSEC may constitute a SDF-1 positive niche in the liver, which can be activated as a hematopoietic microenvironment during extramedullary hematopoiesis. It may occur through the formation of a gradient of SDF-1 at the LSEC microenvironment, as it has been proposed at the BM microenvironment [8].
Based on our in vivo and in vitro results, we propose a model of liver extramedullary hematopoiesis, which involve a crosstalk between LSEC and circulating HSC (Fig. 9). Increased expression of SDF-1 at the liver could be induced by some cytokines such as G-CSF or under certain pathological conditions. As it has been reported that SDF-1 is involved in homing and retention of HSC in the BM [44], it is possible that circulating CXCR4+/HSC may be attracted to an SDF-1 gradient generated at the liver. It may occur through the production of SDF-1 at the LSEC microenvironment, which may retain circulating CXCR4+/HSC in the LSEC niche. In this model SDF-1 expression by LSEC might constitute one of the first signals to induce liver extramedullary hematopoiesis, and the LSEC microenvironments may constitute the earliest hematopoietic niche at the liver. Moreover, liver extramedullary hematopoiesis would be established at the LSEC microenvironment because these cells may provide not only adhesion but also hematopoietic factors for inducing proliferation and differentiation of HSC [24,28], which may result in liver extramedullary hematopoiesis.

Possible role of LSEC microenvironments in liver extramedullary hematopoiesis. During liver extramedullary hematopoiesis, the expression of SDF-1 by LSEC can result in the attraction and homing of circulating CXCR4+-HSC to the liver.
In conclusion, our results show that LSEC constitutively express both SDF-1 and its receptor CXCR4. Our data provide evidence that in the liver, the LSEC microenvironments may constitute one of the main elements to provide SDF-1 signals to attract circulating HSC during liver extramedullary hematopoiesis. The circulating HSC not only would migrate and localize but also adhere and differentiate in the LSEC microenvironments. The data presented here not only demonstrate that signals transduced through CXCR4 may be involved in inducing migration of LSEC, but also that they can be involved in activating some LSEC biological functions such as upregulation of adhesion molecules.
Footnotes
Acknowledgments
The authors wish to thank Olga Wittig and Dr. Carmen Lopez for technical assistance. This work was supported by IVIC and FONACIT grant G2005000405.
Author Disclosure Statement
Authors declare that they do not have any conflict of interest to disclose.