
Abstract
Moving stem cells from bench to bedside has been a challenging task. Undermining this task is comprehending and optimizing the underlying regulatory mechanisms that drive differentiation of stem cells into desired cell and tissue types. Here we present evidence that ribosomal S6 kinase (S6K) is among the proteins upregulated as embryonic stem cells (ESCs) and human induced pluripotent stem cells differentiate into beating cardiomyocytes. We hypothesized that S6K plays a pivotal role in cardiomyogenesis, primarily because it regulates the translation of 3 cardiac-involved genes recently shown to have 5′ terminal oligopyrimidine (5′TOP) sequences: connexin 43 (Cx43), desmoplakin (Dsp), and phosphatase and tensin homolog (PTEN). Along with another independent laboratory, we confirmed that S6K is indeed upregulated in beating ESC-derived cardiomyocytes compared to the surrounding nonbeating, differentiated cells. S6K short interfering RNA-transfected stem cell cultures indicate that inhibition of S6K strongly hinders development of cardiomyocyte beating and translation of Cx43, Dsp, and PTEN; these cardiac 5′TOP mRNAs were only properly translated in cells with S6K, supporting our hypothesis. An unexpected discovery took the role of S6K one step further: S6K-knockdown stem cell cultures developed significantly more neurons than seen in embryoid bodies subjected to a typical cardiac differentiation protocol. These results introduced the novel idea that in addition to its critical cardiac roles, S6K may be a significant factor that prevents stem cells from pursuing a neuronal pathway. Overall, results have indicated the necessity of S6K for normal stem cell cardiomyogenesis, as well as lowered S6K expression for stem cell neurogenesis.
Introduction
U
Regulated upstream by the mammalian target of rapamycin (mTOR) [6,8 –11] and phosphoinositide-3 kinase (PI3K) [13], S6K in turn has important roles in a number of downstream pathways. mTOR inhibition and subsequent inhibition of S6K reduces activation of elongation initiation factor 4B (eIF4B) and phosphorylation of ribosomal protein S6 (rpS6), a substrate of S6K [14]. mTor/S6K is important for initiating translation and appears to be essential for cells to function normally [15]. Perhaps more importantly, inhibition of mTOR has been found to repress a subgroup of mRNAs that include those with 5′ terminal oligopyrimidine (5′TOP) sequences, which function through their secondary structure [14]. Evidence suggests that S6K is an intermediary kinase responsible for this process [7,16,17].
While a single S6K gene is present in some genera such as dS6K in Drosophila, in mammals there are 2 genes—p70 S6K (S6K1) and S6K2—which code for homologous S6Ks [10,18]. When sequentially phosphorylated, S6K1 is activated resulting in the shift of 5′TOP mRNAs into polysomes. This shift may be instigated by an increased rate of binding to ribosomes, caused, in part, by promoting general translation initiation by eIF4B [19]. An S6K1-activated allele also protects 5′TOP mRNAs from rapamycin, whose inhibitory effect on mTOR represses 5′TOP mRNA translation [20] and, with short-term treatment, promotes expansion—as opposed to differentiation—of pluripotent cells [12]. Both the shift into polysomes and the protection conferred by the S6K1-activated allele act to enhance 5′TOP mRNA translation.
The 5′TOP mRNA subgroup has been known for some time to include ribosomal proteins and elongation factors [7,19]. Interestingly, a 2008 study by Yamashita et al. [21] showed that the 5′TOP subgroup also included 3 genes involved in cardiac development. These genes were desmoplakin (Dsp), an essential protein for desmosomes [22,23]; connexin 43 (Cx43), a gap junction protein necessary for heart tube development [24] and hematopoiesis [25]; and phosphatase and tensin homolog (PTEN). The knockout phenotypes for each of these genes possess severe cardiac developmental defects [26 –28].
Based on the above information, we hypothesized that S6K regulates differentiation of stem cells into beating cardiomyocytes by promoting translation of cardiac 5′TOP mRNAs. We found that knocking down S6K significantly delayed cardiac differentiation and, surprisingly, also increased incidence of neural differentiation.
Materials and Methods
ESC culture and cardiomyocyte production
Mouse CCE ESCs were maintained and differentiated similarly to our previously published protocols [1 –5]. Briefly, ESCs were grown on 0.1% gelatin in 1,000 U/mL leukemia inhibitory factor (LIF). Differentiation was induced by removing LIF and converting colonies of ESCs into embryoid bodies (EBs) [3,5]. Briefly, for EB production, ES colonies were picked using a pipette tip and transferred to Petri dishes designed for growing cells in suspension. These EBs were incubated in DMEM with 15% fetal bovine serum for 2 days in the absence of LIF. At the end of incubation day 2, individual EBs were placed into a single well of a 24-well plate, where they adhered and spread. Rhythmic beating of EBs, indicating cardiac muscle differentiation, was monitored by daily inspection of cultures using phase microscopy. In our hands, beating usually commences ∼day 9 after removal of LIF.
Beating areas were isolated from nonbeating areas following our previously published procedure in Refs. [1
–3,5]. In brief, beating areas were isolated using a microscalpel (Fine Science Tools). Any nonbeating cells associated with the beating area were trimmed away. Ten beating and nonbeating areas of similar size were separated and briefly washed in DEPC-treated (RNAse-free) phosphate buffered saline (PBS). Subsequent EBs were analyzed for cardiomyocyte (or alternative tissue) formation by reverse transcriptase-polymerase chain reaction, DIC microscopy, immunofluorescence, and Western blot, or prepared in Kinexus™ sample buffer, frozen, and sent to Kinexus, Inc., for kinase expression analysis. An explanation of how they analyze each protein can be found at
Human induced pluripotent stem cell maintenance and differentiation
Human induced pluripotent stem cells (hiPSCs) were acquired from WiCell. They were maintained according to their protocols. Differentiation was induced by following the protocol by [29].
Antibodies
Anti-S6K was purchased from Abcam. Cx43, PTEN, and β-tubulin (ubiquitously expressed) antibodies used in Western blot Figs. 1, 3, and 6T were purchased from Cell Signaling, Inc. Alexa Fluor® Labeled, neuronal class III β-tubulin (TUJ1) monoclonal antibody used for Fig. 6C–R was purchased from Covance, Inc. Anti-actin was purchased from BioLegend. Anti-cardiac troponin T was purchased from Neomarkers. Anti-Dsp was purchased from American Research Products. Antibodies for Nkx2.5 and Gata4 were purchased from Santa Cruz.
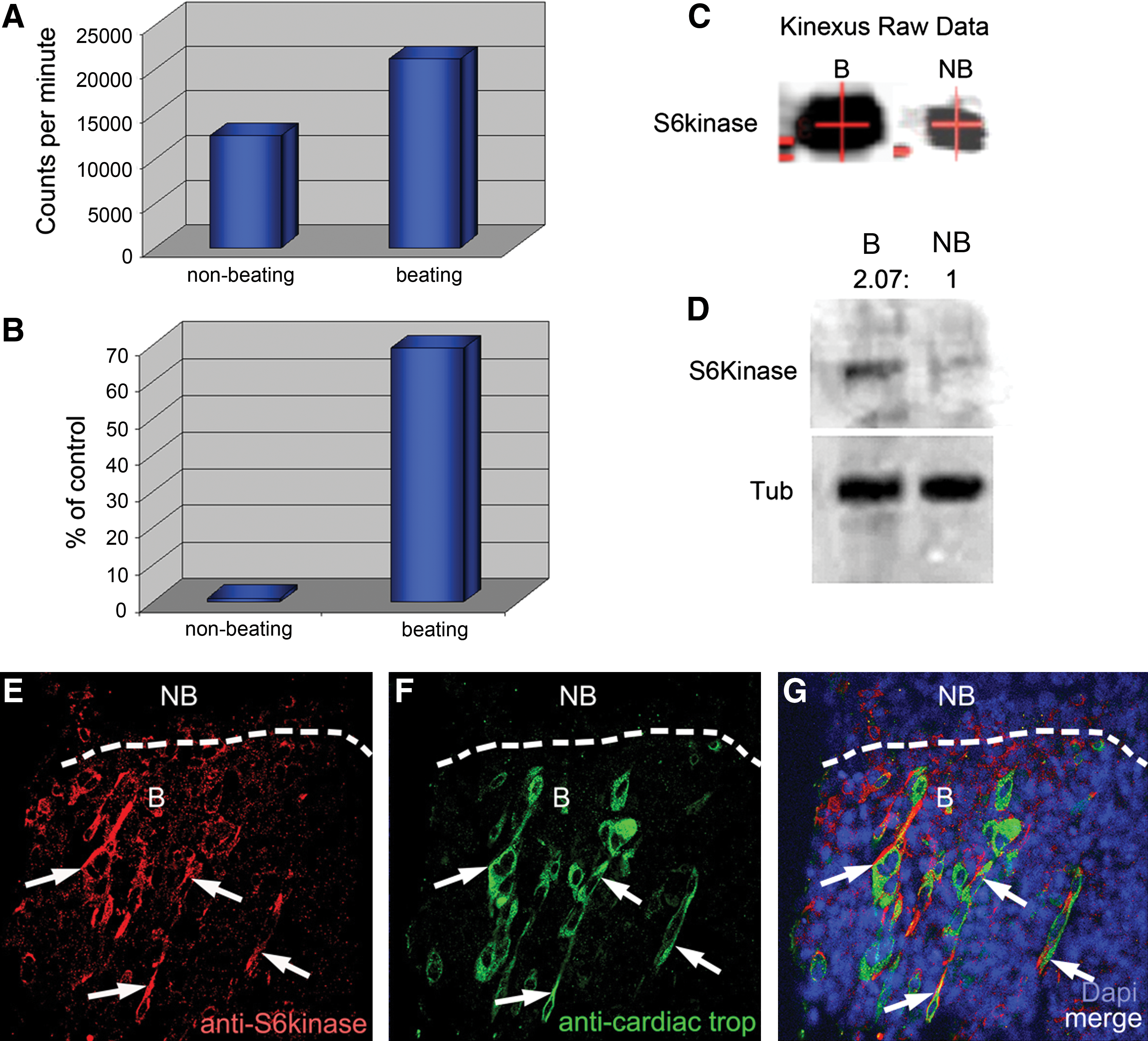
Comparison of nonbeating versus beating areas in ESC-derived cardiomyocytes. Graphs show raw and normalized data taken directly from the Kinexus protein kinase screen. Kinexus, Inc. (
Confocal microscopy and cell counting
Beating and nonbeating areas were surgically spliced out of EBs using microdissection and prepared for confocal microscopy [1,2]. The protocol for confocal microscopy was performed similarly to [1 –5] on these areas. Primary antibodies were diluted 1:50 and secondary AlexaFluor antibodies purchased from Invitrogen, Inc. were diluted 1:800. An Olympus Fluoroview 500 Laser Scanning Microscope (Olympus America, Inc.) was used to image the samples.
Flow cytometry
Mouse embryonic stem cells (mESCs) were differentiated according to [1]. Cells were washed in PBS, trypsinized (0.05%), and fixed in 2% paraformaldehyde. Primary antibodies used were either cardiac troponin, or Cx43. Secondary antibody used was AlexaFluor 488 antibody (Invitrogen, Inc.) against the appropriate primary antibody. Members of the Georgetown University Flow Cytometry Core Facility acquired the flow cytometry data. Analysis was done in FCS Express 4 Flow Research Edition (De Novo Software).
Short interfering RNA production and transfection
S6K short interfering RNA (siRNA) was ordered from Proligo. S6K siRNA was transfected into the mouse CCE ESCs to specifically inhibit S6K at the mRNA level, preventing translation of the S6K protein. Western blotting confirmed the downregulation of S6K. Addition of 1–3 μL of Qiagen's HiPerfect Transfection Reagent per 300 μL of CCE differentiating media in each well of cells served as a control. siRNA was transfected into EBs on day 6 post removal of LIF from the medium.
Statistical analysis
All experimental trials were repeated 3 times, minimum. Protein expression was measured using histogram analysis software that measured pixel intensity. Each experimental band representing a protein (eg, Dsp Fig. 3) was normalized to the control band (eg, actin). The density of control was measured against S6K siRNA-treated cells. We did not see any marked difference between the outcomes of each blot for each protein. Graphical data displays are plotted as means with error bars representing the standard error of the mean of the trials. Statistical significance was determined by unpaired t-tests.
Results
S6K is upregulated in beating versus nonbeating areas
Comparison of protein levels in beating and nonbeating areas in a proteomic screen (Kinexus, Inc.) showed that S6K concentration was significantly higher (69%) in nascent beating areas than in nonbeating areas (Fig. 1A–C). These results were independently confirmed by our laboratory, using Western blotting (Fig. 1D).
Confocal imaging revealed the cytosolic presence of S6K [10], which showed greater expression in cells within the newly beating areas (1–2 days after the onset of beating) than cells within nonbeating areas (Fig. 1E). Furthermore, areas with high levels of S6K often coincided with areas that developed into beating areas positive for cardiac troponin T (CnnT; Fig. 1F, G).
S6K is necessary for normal cardiomyogenesis
Mouse ESCs are an excellent model system for analyzing cardiomyogenesis because after removal of LIF and plating EBs, beating typically occurs ∼9 days in small areas that can then be quantified [1,2,5]. S6K expression was inhibited with siRNA 3 days before the commencement of beating. S6K-repressed EBs were monitored for beating over a 132-h period and compared to control, untreated EBs, or mock-treated siRNAs. The number of wells containing EBs that developed at least one beating area was significantly less at 72 and 96 h in those transfected with S6K siRNA when compared to those transfected with scrambled siRNA (Fig. 2A). The siRNA wells showed an increase in beating over time due to the degradation of the siRNA [2,30]. EBs at each time point in the graph in Fig. 2A were analyzed by Western blot (Fig. 2B), which showed the knockdown effect of S6K siRNA.
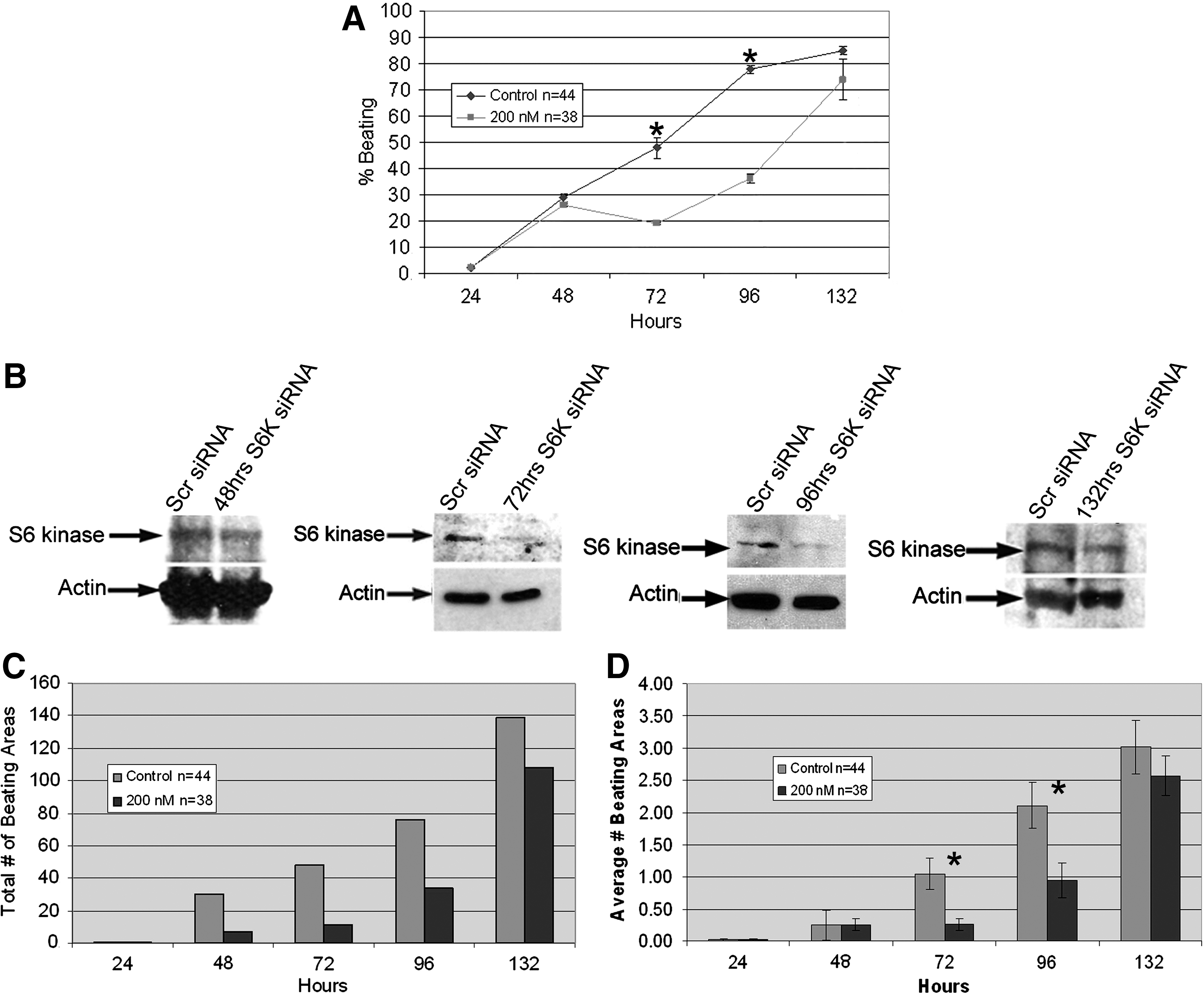
Treatment of ESCs with S6K siRNA strongly hindered differentiation of ESCs into beating areas.
The numbers of beating areas in siRNA and control wells for each experimental trial were also counted. Pooling together the data sums for each time point revealed an obvious difference in number of beating areas between the control groups and the siRNA groups (Fig. 2C). Further analysis of the data made this discrepancy even clearer. Throughout the entire 132 h, the average number of beating areas per control well was always greater than that of the siRNA wells (Fig. 2D). Significance (p<0.05) was reached at 72 and 96 h.
S6K regulates the translation of key cardiac 5′TOP genes
S6K has previously been implicated in the translation of 5′TOP mRNAs [17 –19,31]. To date, of the 1,645 putative 5′TOP mRNAs [21], we have identified 3 to be important for cardiac development: Cx43, Dsp, and PTEN (Fig. 3A). We surmised that if S6K expression was suppressed using siRNA, then the protein products of the three 5′TOP mRNAs involved in cardiac development would also be suppressed. EBs were transfected with control siRNA or siRNA to S6K 6 days after removal from LIF. This is equivalent to ∼3 days before beating normally occurs. Four days after transfection, EBs were accumulated and analyzed by Western blot. Interestingly, knockdown of S6K markedly reduced the expression of all 3 mRNAs with 5′TOP sequences (Fig. 3B); protein levels of both isoforms of Dsp (Dsp I and Dsp II) showed decreased expression levels, as did Cx43 and PTEN. Non-5′TOP miRNA products were assessed to determine cardiomyocyte differentiation. Expression of NkX2.5, cardiac troponin (CnnT), Gata4, and actin and β-tubulin (ubiquitously expressed) were not decreased by S6K siRNA (Fig. 3C–C′′′), suggesting that S6K primarily affects specific components of cardiomyogensis.
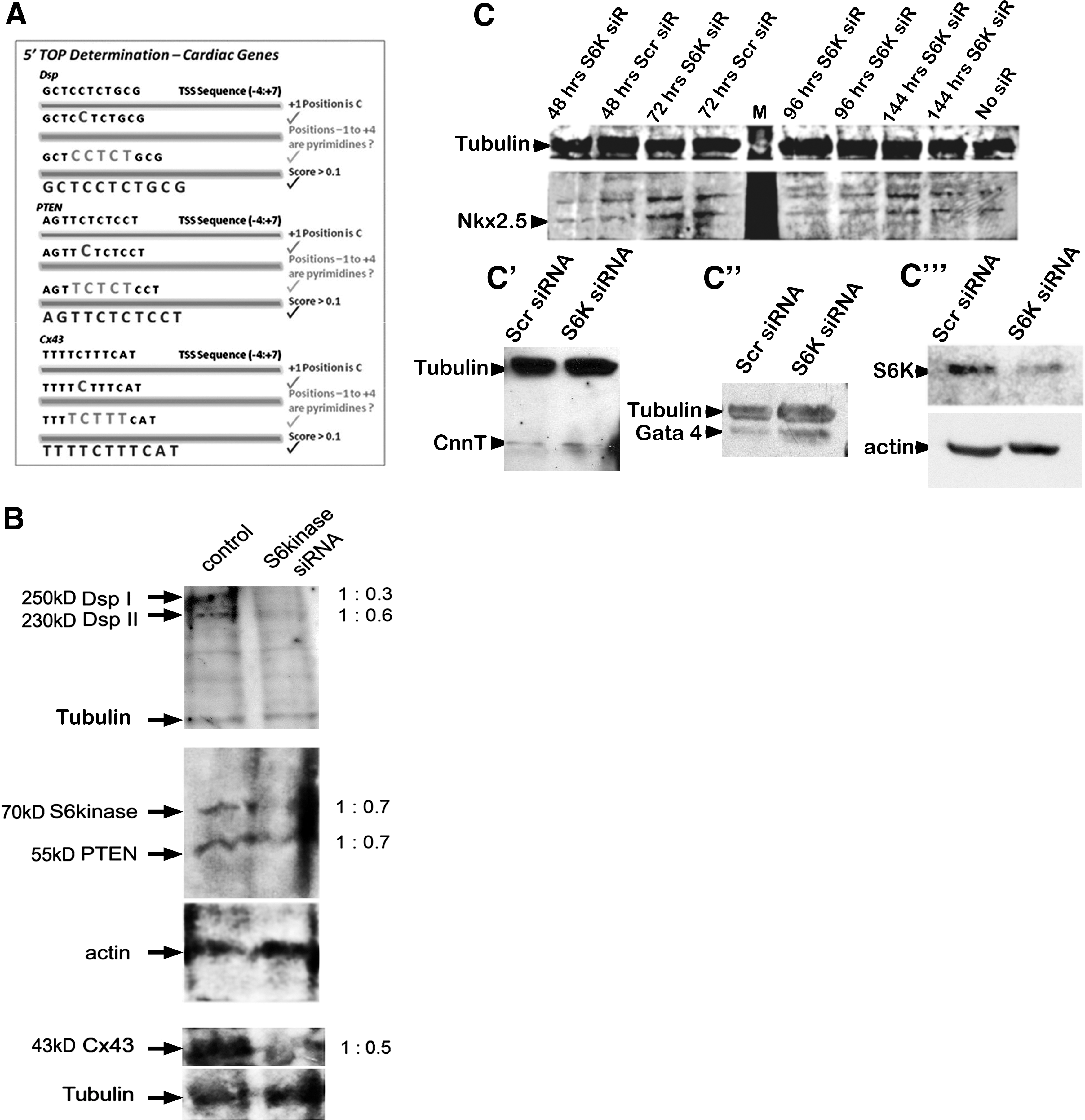
siRNA to S6K results in down-regulation of cardiac genes with 5′TOP sequences.
Subjecting cells to flow cytometry, we were able to quantify the number of mESCs affected by S6K siRNA (Fig. 4). We found that the percentage of cells expressing cardiac troponin was not significantly different between the Mock siRNA and S6K siRNA treatments; however, the number of differentiated mESCs expressing the 5′TOP mRNA product Cx43 was significantly reduced. This provided further evidence that S6K is necessary for normal cardiac development.
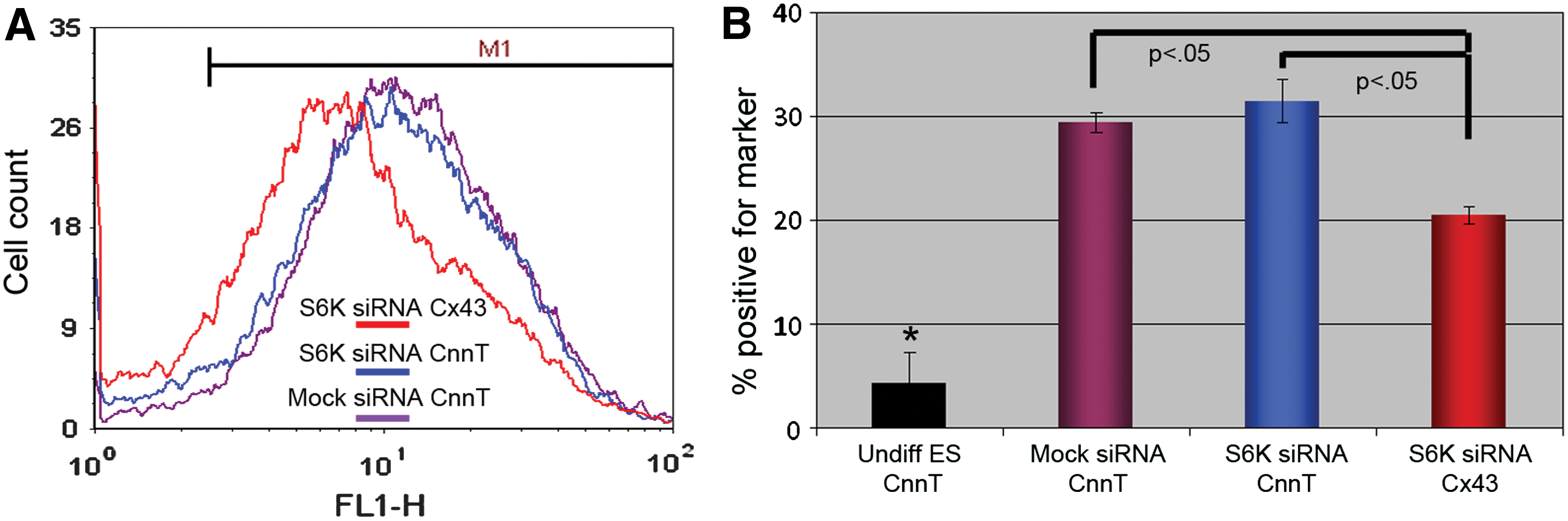
FCM analyses confirm our Western blot, confocal, and morphological data.
Similar to mESCs, suppression of S6K in hiPSCs results in aberrant cardiomyocyte differentiation
To verify the ubiquity, of S6K function during cardiomyogenesis, we transfected S6K siRNA into hiPSCs during differentiation into the cardiac lineage [12]. Four days after transfection, EBs were analyzed morphologically for their capacity to fully differentiate into beating areas. Figure 5 reveals that S6K siRNA markedly reduced the average and total number of beating areas derived from human iPS.
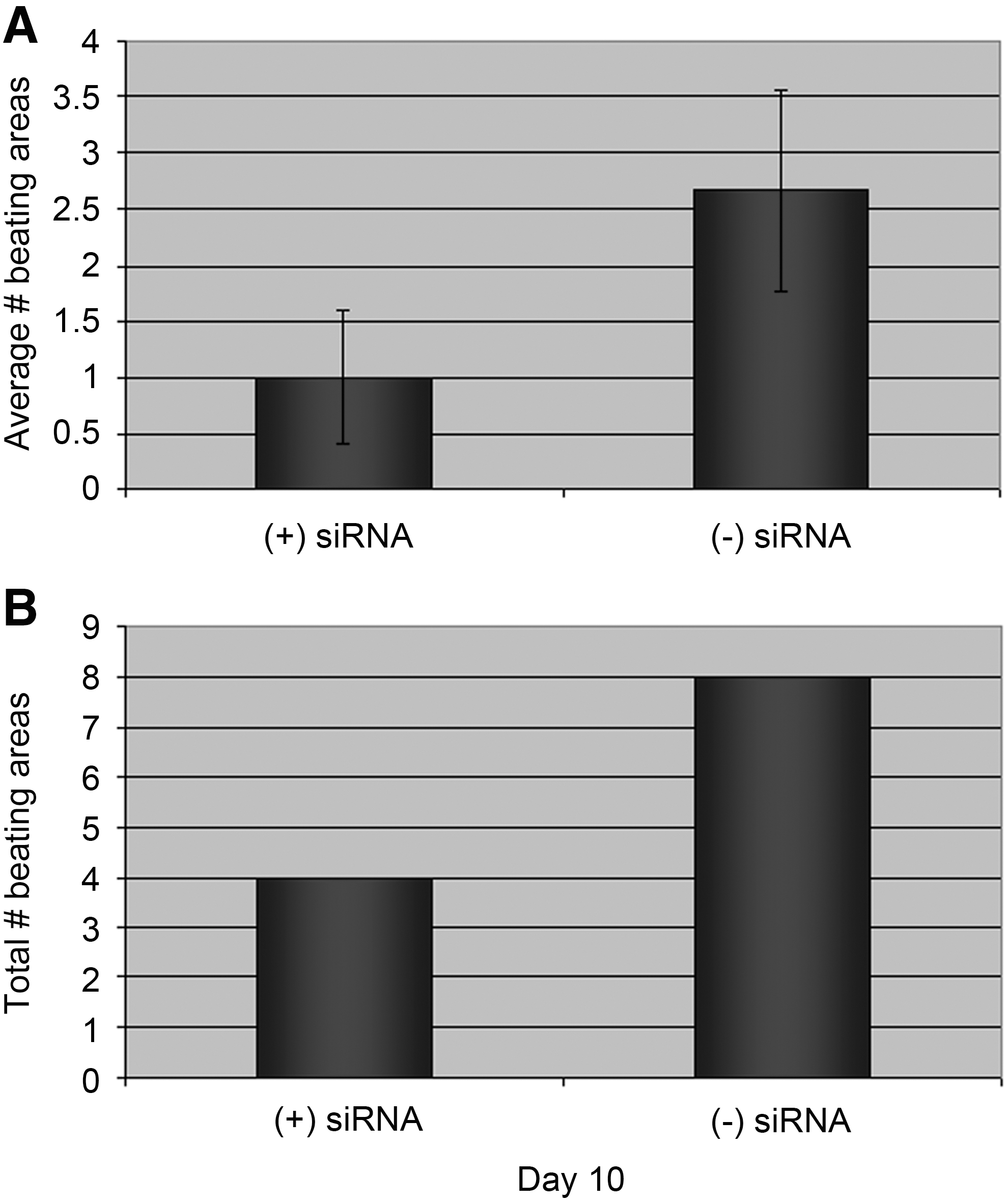
To determine if S6K activity was restricted to mouse cardiac development or was conserved, hiPS cells were subjected to S6K siRNA. The average number
Inhibiting S6K unexpectedly shifts differentiation towards a neural fate
We consistently observed that after S6K siRNA degraded, the number of beating areas would approach, but not draw even with, the number of beating areas within control EBs (Fig. 2). This observation continued in most experiments up to 21 days after transfection of siRNA. Surprisingly, 7–10 days after siRNA transfection, cell morphology within the spreading EBs was noticeably different. High-magnification views revealed that EBs subjected to S6K siRNA repeatedly resulted in areas containing numerous neurons compared to control EB cultures (Fig 6A, B). N.b., no neural-differentiation media was used in these experiments. This result was further confirmed using different S6K siRNA concentrations, which revealed that the shift to a neural lineage was dependent upon the dosage of S6K siRNA (Fig. 6C–R). Quantifying multiple areas from multiple experiments provided further evidence that S6K siRNA pushes ESCs down the neural pathway (Fig. 6S). Finally, Western analyses clearly revealed that suppression of S6K promoted differentiation toward a neural fate as neurofilament M (NF-M) protein was detected as soon as 7 days after S6K siRNA transfection (Fig. 6T). In contrast, 7 days after Mock siRNA, NF-M was below the level of detection in mESCs treated with scrambled siRNA. As a control, given enough time in culture, day 14 post-siRNA treatment (21 days postremoval from LIF), as might be expected, NF-M was eventually detected in scrambled siRNA-treated EBs.

ESCs treated with S6K siRNA and cultured for an extended time form neurons. Neurons were very easy to spot by simple morphological observations using phase-contrast microscopy
In vitro analysis of S6K expression mimics in vivo expression during early development
We surmised that if upregulation of S6K is important for cardiomyogenesis and its down-regulation is necessary for neurogenesis in vitro, then this scenario should be mirrored in vivo. E9.5 day embryos (Fig. 7) revealed that S6K is highly expressed in cardiomyocytes within the developing heart tube when compared to its expression in the neural tube.
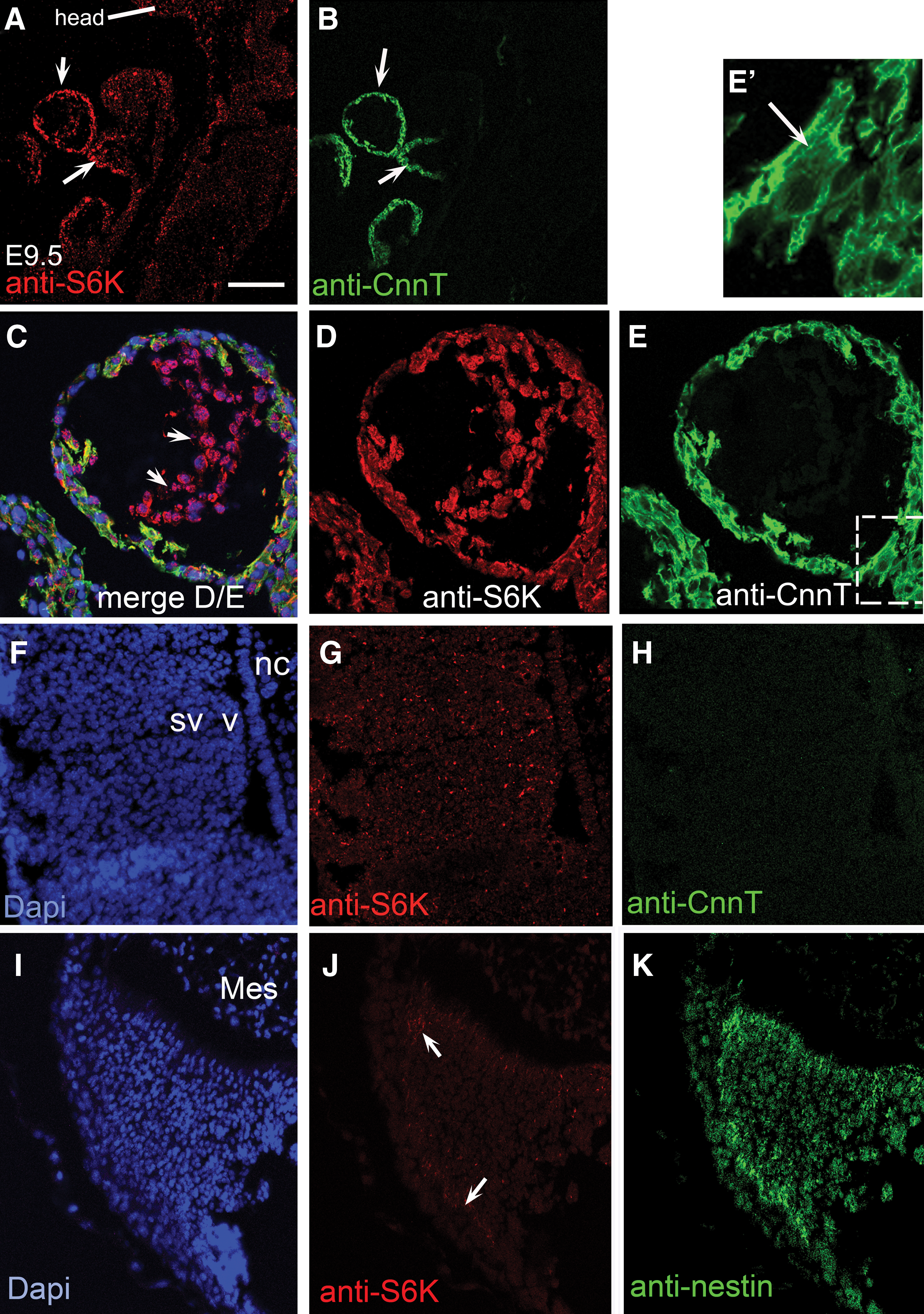
In vivo analysis during development reveals an S6K localization pattern that mimics differentiation in vitro.
Discussion
A number of studies have analyzed the functional necessity of S6K, and confirmed its role in 5′TOP mRNA translation [17,18,31]; however, few thus far have examined the implications of S6K activity in stem cell differentiation [12]. Here, we provide evidence in human and mouse pluripotent cells that during differentiation, S6K is up-regulated in newly beating areas isolated from EBs. Furthermore, inhibiting S6K abrogates development of beating areas in both mouse and human pluripotent cells. Surprisingly, down-regulation of S6K also appears to be important in neuronal developmental.
S6K plays a central role in 5′TOP mRNA translation because it was shown to enable more efficient translation of the mRNAs, including those necessary for generating cardiac development [21]. The S6K knockouts further accentuated the importance of this kinase in vivo. Of the 2 mammalian forms of S6K [10,18], S6K1 is the gene chiefly responsible for regulating cell size in mammals [32 –40]. Single knockouts of S6K2 in mice (S6K2–/– ) resulted in no significant alteration of the normal phenotype [20,41]. The phenotypes of S6K1–/– and S6K1–/–/S6K2–/– mice were much the same—with the exception of significantly higher incidence of lethality in double knockout mice [20] with complications indicative of heart developmental defects. Nonviable mice showed multiple areas of hemorrhaging within their myocardium [20].
Past experimentation has shown that S6K1 is responsible for the 5′TOP mRNA translation-promoting effects [17]. S6K2 is only able to compensate partially in the absence of S6K1 [18]. As a result, it is plausible that S6K1–/–/S6K2–/– mice and their hearts were unable to develop normally because they were without the benefit of p70 S6K1 function, and thus of the translation of cardiac 5′TOP mRNAs Cx43, Dsp, and PTEN.
Of high interest is the alteration of EB phenotype from cardiac to neural 1 week after S6K siRNA treatment. The in vivo analysis supported the in vitro data as S6K expression was markedly lower in the neural tube versus the heart tube. The mechanism driving this shift is not known [42]; however, our work does corroborate with that of D'Aniello et al. [43], who showed that an association between S6K and a G-protein tilts differentiation toward the cardiac lineage. Understanding this mechanism will be a focus of future work.
Discovery of the key mechanisms involved in stem cell differentiation into tissue-specific cell populations is necessary before stem cell therapies can proceed safely. Comprehension of the kinetics of these critical factors will lead to the production of purer cell populations, more effective than shot-in-the-dark approaches heavily reliant on chance. Increased population purity will, in turn, facilitate improvement in stem cell therapy quality control and in so doing minimize the risks involved in such therapies.
Footnotes
Acknowledgments
We thank Matthew R. Pearce, Mentorship Program Director at The Thomas Jefferson High School for Science and Technology, Alexandria, VA. Ms Li started this project while in this program, which enables students to do research with mentors in laboratories at major universities. Dr. Gallicano served as Ms. Li's mentor. Also, we thank Dr. Shayn Peirce in the Department of Biomedical Engineering, University of Virginia for generous use of her lab to complete some final experiments and Dr. Anindya Dutta in the Department of Biochemistry and Molecular Genetics, UVA for assisting in the editing of the paper and giving useful suggestions for improvement of the manuscript. This work was funded in part by an NIH grant R01HL070204-05.
Author Disclosure Statement
All authors declare that no competing financial interests exist.