
Abstract
Transplanted human bone marrow (BM) and umbilical cord blood (UCB) progenitor cells activate islet-regenerative or revascularization programs depending on the progenitor subtypes administered. Using purification of multiple progenitor subtypes based on a conserved stem cell function, high aldehyde dehydrogenase (ALDH) activity (ALDHhi), we have recently shown that transplantation of BM-derived ALDHhi progenitors improved systemic hyperglycemia and augmented insulin secretion by increasing islet-associated proliferation and vascularization, without increasing islet number. Conversely, transplantation of culture-expanded multipotent-stromal cells (MSCs) derived from BM ALDHhi cells augmented total beta cell mass via formation of beta cell clusters associated with the ductal epithelium, without sustained islet vascularization. To identify paracrine effectors produced by islet-regenerative MSCs, culture-expanded BM ALDHhi MSCs were transplanted into streptozotocin-treated nonobese diabetic/severe combine immune deficient (SCID) mice and segregated into islet-regenerative versus nonregenerative cohorts based on hyperglycemia reduction, and subsequently compared for differential production of mRNA and secreted proteins. Regenerative MSCs showed increased expression of matrix metalloproteases, epidermal growth factor receptor (EGFR)-activating ligands, and downstream effectors of Wnt signaling. Regenerative MSC supernatant also contained increased levels of pro-angiogenic versus pro-inflammatory cytokines, and augmented the expansion of ductal epithelial but not beta cells in vitro. Conversely, co-culture with UCB ALDHhi cells induced beta cell but not ductal epithelial cell proliferation. Sequential transplantation of MSCs followed by UCB ALDHhi cells improved hyperglycemia and glucose tolerance by increasing beta cell mass associated with the ductal epithelium and by augmenting intra-islet capillary densities. Thus, combinatorial human progenitor cell transplantation stimulated both islet-regenerative and revascularization programs. Understanding the progenitor-specific pathways that modulate islet-regenerative and revascularization processes may provide new approaches for diabetes therapy.
Introduction
T
There is mounting evidence that readily accessible and clinically applicable progenitor cells from bone marrow (BM) stimulate islet regeneration after transplantation [19]. We have shown that transplantation of BM-derived stem cells augmented glycemic control in streptozotocin (STZ)-treated mice [20]. Donor progenitor cells were recruited to damaged islets and induced proliferation in recipient beta cells. Importantly, insulin-expressing donor cells were not detected during blood glucose reduction [20,21]. Other groups have reported the capacity of multipotent stromal cells (MSCs) to induce native islet regeneration via paracrine mechanisms [22 –25], suggesting that progenitors from several mesodermal lineages may contribute to islet regeneration [20,22].
We have recently shown that the mechanisms of islet regeneration induced after human progenitor transplantation depended on the progenitor cell types administered [26]. Using fluorescence-activated cell sorting (FACS) purification of multiple progenitor subtypes based on high aldehyde dehydrogenase (ALDH) activity [27,28], transplantation of BM-derived ALDHhi-mixed progenitors [29] improved systemic hyperglycemia and augmented insulin secretion by increasing islet size and vascularization, without increasing islet number. Augmented proliferation within regenerated islets and associated vascular endothelium indicated the stimulation of islet-specific proliferative and pro-angiogenic programs [26]. Conversely, transplantation of culture-expanded MSCs derived from BM ALDHhi cells augmented total beta cell mass via increased formation of small beta cell clusters associated with the ductal epithelium, without sustained beta cell proliferation or islet vascularization [26].
The paracrine interactions whereby specific progenitor cell types modulate islet regeneration remain largely unknown and represent the key to optimizing the regenerative capacity of the pancreas. The aims of these studies were to identify paracrine targets common to progenitor cells that stimulated islet regeneration, and to test the hypothesis that sequential transplantation of BM-derived MSCs and umbilical cord blood (UCB)-derived ALDHhi progenitor cells will stimulate islet formation, and sustain subsequent islet growth and vascularization, respectively.
Research Design and Methods
Progenitor cell isolation and MSC culture
Human BM and UCB were obtained with informed consent from the London Health Sciences Centre (London, ON). Cells were incubated with Aldefluor™ reagent (StemCell Technologies, Vancouver, BC) and FACS-purified (FACSAriaIII; BD Biosciences, Mississauga, ON) for ALDH activity as previously described [27,28]. MSCs were established from BM ALDHhi cells by plastic adherence at limiting dilution and expanded in Amniomax™ media (Invitrogen, Burlington, ON) [29].
RNA isolation and microarray
mRNA was isolated in triplicate from independent BM MSC samples (n=4) at passages 4, 6, and 8, and from UCB ALDHlo and ALDHhi cells (n=3) using mRNeasy Mini kits (Qiagen, Dusseldorf, Germany). Microarray was performed using human gene array chips (Affymetrix, Santa Clara, CA) at the London Regional Genomics Centre. Genes differentially expressed by regenerative versus nonregenerative MSCs, or by UCB ALDHhi versus ALDHlo cells were analyzed using Partek Genomics Suite (Partek, Inc., St. Louis, MO). Unfiltered data have been deposited in a public repository.
Supernatant enzyme-linked immunosorbent assay
MSCs from 3 regenerative and 3 nonregenerative human MSC samples were expanded in Amniomax™ media. Twenty-four hours after media change at passage 3, supernatant was collected and supplemented with dipeptidyl peptidose-4 and protease inhibitor cocktail (Sigma-Aldrich, St. Louis, MO). Multiplex ELISA (Millipore, Billerica, MA) was performed at the Lawson ELISA Core Facility.
Co-culture of progenitor cells with βTC6 and PANC-1 cells
Murine beta cell insulinoma (βTC6, ATCC-11506) or human ductal epithelial carcinoma (PANC-1 and ATCC-1469) cells were co-cultured with MSC-conditioned supernatant (secretion by 4×104 cells for 24 h), regenerative human MSCs (4×104 cells), or UCB ALDHlo or ALDHhi cells (4×104) in 5 μm pore Millicell™ hanging cell culture inserts (Millipore). After 3-day (PANC-1) or 5-day (βTC6) culture, viable cells were enumerated using trypan blue exclusion.
Transplantation of hyperglycemic nonobese diabetic/severe combined immune deficient mice
Eight- to 10-week-old nonobese diabetic (NOD)/severe combine immune deficient (SCID) mice (Jackson Labs, Bar Harbor, ME) were intraperitoneally injected with 35 mg/kg/day STZ (Sigma-Aldrich) for 5 consecutive days as previously described [20]. On day 10, hyperglycemic mice (15–25 mmol/L) were sublethally irradiated (300cGy) and transplanted by tail vein injection with either phosphate-buffered saline (PBS) or BM-derived MSCs (5×105). On day 17, mice received an additional injection of PBS, or UCB-derived ALDHlo or ALDHhi cells (2×105), or sample-matched MSCs (2×105). Nonfasted blood glucose was monitored weekly (Abbott Diabetes, Alameda, CA). Twenty-four hours before euthanasia, mice received 200 μg EdU by intraperitoneal (i.p.) injection (Invitrogen), and glucose tolerance was performed by i.p. injection with 2.0 g/kg glucose and glycemia was monitored for 2 h. On day 42, serum was collected for insulin quantification, and tissues were analyzed by FACS for human cell engraftment as previously described [26].
Immunohistochemical and immunofluorescent analysis
The splenic portion of the pancreas was frozen, sectioned, and stained for insulin to quantify islet number, islet size, and total beta cell mass. Immunofluorescent staining was performed to quantify islet blood vessel density (vWF or CD31), islet-duct association (CK19), and EdU-incorporation by islet cells as previously described [26]. Data were expressed as mean±standard error of mean, and analysis of significance was performed by 1-way analysis of variance (ANOVA) for histochemistry, and by 2-way ANOVA for blood glucose concentrations and glucose tolerance.
Results
Transplanted MSCs demonstrated variable capacity for islet regeneration
MSCs established after ALDHhi cell purification from 12 human BM samples, were ex vivo expanded and tail vein-injected into STZ-treated NOD/SCID mice on day 10. On the basis of recovery from hyperglycemia versus PBS-injected mice (mean glucose on days 17–42, MGD17–42=30.5±1.4 mmol/L), samples were divided into tertiles representing highly regenerative (Rhi, MGD17–42=15.9±2.7 mmol/L), moderately regenerative (Rmid, MGD17–42=22.4±3.5 mmol/L), and nonregenerative MSCs (Rlow, MGD17–42=30.6±1.8 mmol/L). These analyses produced glucose concentrations where Rhi-MSCs<Rmid-MSCs<Rlow-MSCs (*P<0.05, Fig. 1A). Three of 12 samples (25%) were Rhi, 4 samples (33%) were Rmid, and 5 samples (42%) were Rlow, permitting subsequent microarray analyses of global mRNA expression comparing Rhi-MSCs versus Rmid-MSCs versus Rlow-MSCs.

Functional and genomic characterization of human BM-derived MSCs with high versus low islet regenerative capacity. MSC cultures were established from ALDHhi cells from 12 human BM samples.
Prolonged ex vivo expansion of MSCs reduced islet regenerative functions
Rhi-MSCs (n=3) were expanded for 8 passages ex vivo, and transplanted at 5×105 cells into hyperglycemic NOD/SCID mice at P4, P6, or P8. We have previously shown that >95% of ALDHhi-MSCs express MSC markers (CD90, CD73, and CD105), and retain multipotent capacity to form bone, cartilage, and adipose lineages up to P8 [29]. Rhi-MSCs transplanted at P4 demonstrated robust reduction in hyperglycemia at days 17–42 (MGD17–42=14.7±3.7 mmol/L). Sample-matched MSCs transplanted at P6 (MGD17–42=22.4±2.5 mmol/L) or P8 (MGD17–42=30.1±2.6 mmol/L) showed a stepwise increase in systemic glucose concentrations (Fig. 1B), indicating that islet regenerative function was reduced as a result of prolonged culture. These analyses produced glucose concentrations such that P4-MSCs<P6-MSCs<P8-MSCs (*P<0.05, Fig. 1B), permitting microarray analyses of global mRNA expression comparing Rhi-MSCs at P4 versus P6 versus P8 as blood glucose lowering capacity was diminished in vivo.
Regenerative MSCs demonstrated increased transcription of matrix metalloproteases, epidermal growth factor receptor family ligands, and downstream effectors of Wnt-signaling
To identify novel targets associated with islet-forming function after transplantation [26], global mRNA expression was assessed in triplicate on MSCs expanded from 4 independent BM samples at P4, P6, and P8, using a total of 36 Affymetrix™ gene chips. Representative analyses of transcripts with >2.5-fold differential expression between Rhi-MSCs versus Rlow-MSCs revealed 29 unique genes translating to known secreted proteins (Fig. 1C). Upregulated genes included matrix metalloproteases (MMP1 and MMP13), insulin-like growth factor binding protein 5 (IGFBP5), downstream effectors of Wnt-signaling (WISP1 and SFRP1), and amphiregullin (AREG), a ligand of the epidermal growth factor receptor (EGFR) superfamily. Interestingly, these genes consistently demonstrated intermediate expression in Rmid-MSCs (Fig. 1C), indicating that increased expression of these transcripts correlated with high islet regenerative capacity in vivo. Rhi-MSCs also showed decreased expression of pro-inflammatory cytokines (IL1a, IL1β, and IL33) and chemokines (CCL2 and CXCL6), suggesting that decreased expression of pro-inflammatory genes was associated with high islet regenerative function [30]. Identical analyses comparing Rhi-MSCs at P4 versus P8 (Fig. 1D) again revealed transcripts for the MMPs, and several EGFR superfamily ligands (AREG, EREG, EPGN, and HBEGF), as well as targets implicated in angiogenesis (FGF16, ANGPTL4, and HGF) and tissue repair (BMP2 and JAG1), were highest at P4, intermediate at P6, and lowest at P8. Thus, reduced expression of these transcripts correlated with diminished regenerative function after prolonged ex vivo culture.
Venn diagrams were generated comparing >2-fold differential gene expression between all regenerative versus nonregenerative samples, and revealed 16 unique genes that were increased in all 3 regenerative samples (triple hits), and an additional 47 genes increased in 2 of 3 regenerative samples (double hits; Fig. 1E). Filtering these genes for external localization (extracellular or membrane bound) generated a list of 11 secreted targets, including MMP1, MMP13, CYTL1, WISP1, SFRP1, and 7 cell surface targets that included KDR vascular endothelial growth factor receptor 2 (VEGFR2) and CD24 (Table 1). Using an alternative strategy, the double or triple hits with increased expression in Rhi-MSCs versus Rlow-MSCs were cross-referenced with transcripts with increased expression in Rhi-MSCs at P4 versus P8 (Fig. 1F). These analyses generated a list of 5 secreted targets, including MMPs, AREG, and IGFBP5, and 5 cell surface targets, again including KDR and CD24 (Table 2). Collectively, these data identified cell surface marker expressions common to MSCs with islet regenerative function, and repeatedly implicated the MMPs, EGFR superfamily members, and downstream effectors of Wnt signaling as potential regulators of MSC-induced islet regeneration.
FC, fold change; VEGFR2, vascular endothelial growth factor receptor 2.
EGFR, epidermal growth factor receptor.
UCB ALDHhi cells were comprised of hematopoietic and endothelial progenitors, and expressed stem cell-associated surface markers and cytokines
Uncultured BM ALDHhi cells contain multiple progenitor subtypes with islet regenerative potential [26], but healthy donor sample availability is limited due to the invasiveness of BM aspiration. Because UCB samples represent an analogous population more readily accessible for preclinical studies, we similarly characterized the hematopoietic, endothelial, and mesenchymal colony-forming capacity (CFC) of UCB ALDHlo versus ALDHhi cells (n=4–6), and performed Affymetrix microarray on matched cell cohorts (n=3). UCB ALDHhi cells were highly enriched for multipotent hematopoietic CFC (1 in 3.6 ALDHhi cells), and for endothelial CFC (1 in 4.6×104 ALDHhi cells). In contrast to BM-derived ALDHhi cells that possessed CFC capacity all 3 progenitor lineages [26], UCB ALDHhi cells were devoid of mesenchymal CFC capacity, allowing further investigation of regenerative functions from culture-expanded BM MSCs and uncultured UCB ALDHhi hematopoietic and endothelial progenitors.
Representative gene array analyses for transcripts with >5-fold differential expression in 3 UCB ALDHhi cell cohorts compared with sample-matched ALDHlo cells generated a list of 256 unique genes translating to membrane bound or extracellular localizations. As an internal control for ALDH activity, ALDH1A1 mRNA expression was increased 42-fold in ALDHhi versus ALDHlo cells (P=2.08×10−5). The most highly expressed genes in ALDHhi cells (Supplementary Fig. S1A; Supplementary Data are available online at
To identify paracrine effectors with increased expression in UCB ALDHhi progenitors, differentially expressed transcripts filtered for known extracellular targets included myeloid cell-secreted cytokines (IL18, IL1B, and CYTL1), pro-angiogenic factors (ANGPT1), and similar to Rhi-MSCs, ligands of the EGFR superfamily (AREG and EREG). Downregulated genes included secreted chemokine ligands (CCL5 and CCL4L1), potent immuno-stimulatory factors such as interferon-γ (IFNG), and effectors produced by lymphocytes and natural killer (NK) cells (PRF1 and GZMK; Supplementary Fig. S1B). Collectively, UCB ALDHhi purification selected for cells with primitive stem cell and myeloid progenitor phenotypes, depleted cells with mature lymphocyte and natural killer (NK)-cell phenotypes [27,31], and identified cytokines highly expressed by UCB ALDHhi progenitor cells [26].
Islet-regenerative MSCs demonstrated increased secretion of pro-angiogenic cytokines and decreased secretion of pro-inflammatory cytokines
Rhi-MSC (n=3) and Rlow-MSC (n=3) supernatants were analyzed by Multiplex™ ELISA to further identify secreted proteins associated with islet regenerative capacity. On the basis of our microarray data (Fig. 1C–F), we focused on hepatocyte growth factor (HGF), a cytokine with high expression in Rhi-MSCs at early passage with previously documented islet-repair function [32 –34], and on the axis between pro-angiogenic versus pro-inflammatory cytokines. Compared with Rlow-MSC supernatants, Rhi-MSC supernatants showed increased concentrations of HGF and several pro-angiogenic cytokines [VEGF, basic fibroblast growth factor (bFGF), and Ang-1; Fig. 2A], and decreased concentrations of cytokines involved in inflammation [interleukin-6 (IL-6)] and chemoattraction (monocyte chemotactic protein [MCP-1]; Fig. 2B). Cross-referencing to the microarray data (Fig. 1C) comparing Rhi-MSCs versus Rlo-MSCs confirmed increased mRNA expression for VEGFA [fold change (FC)=1.33], bFGF or FGF2 (FC=1.36), Ang-1 or ANGPT1 (FC=1.43), and decreased mRNA expression for IL6 (FC=−1.25) and MCP-1 or CCL2 (FC=−2.44). Collectively, these data indicated that islet-regenerative MSCs demonstrated a pro-angiogenic versus pro-inflammatory secretory phenotype in vitro.
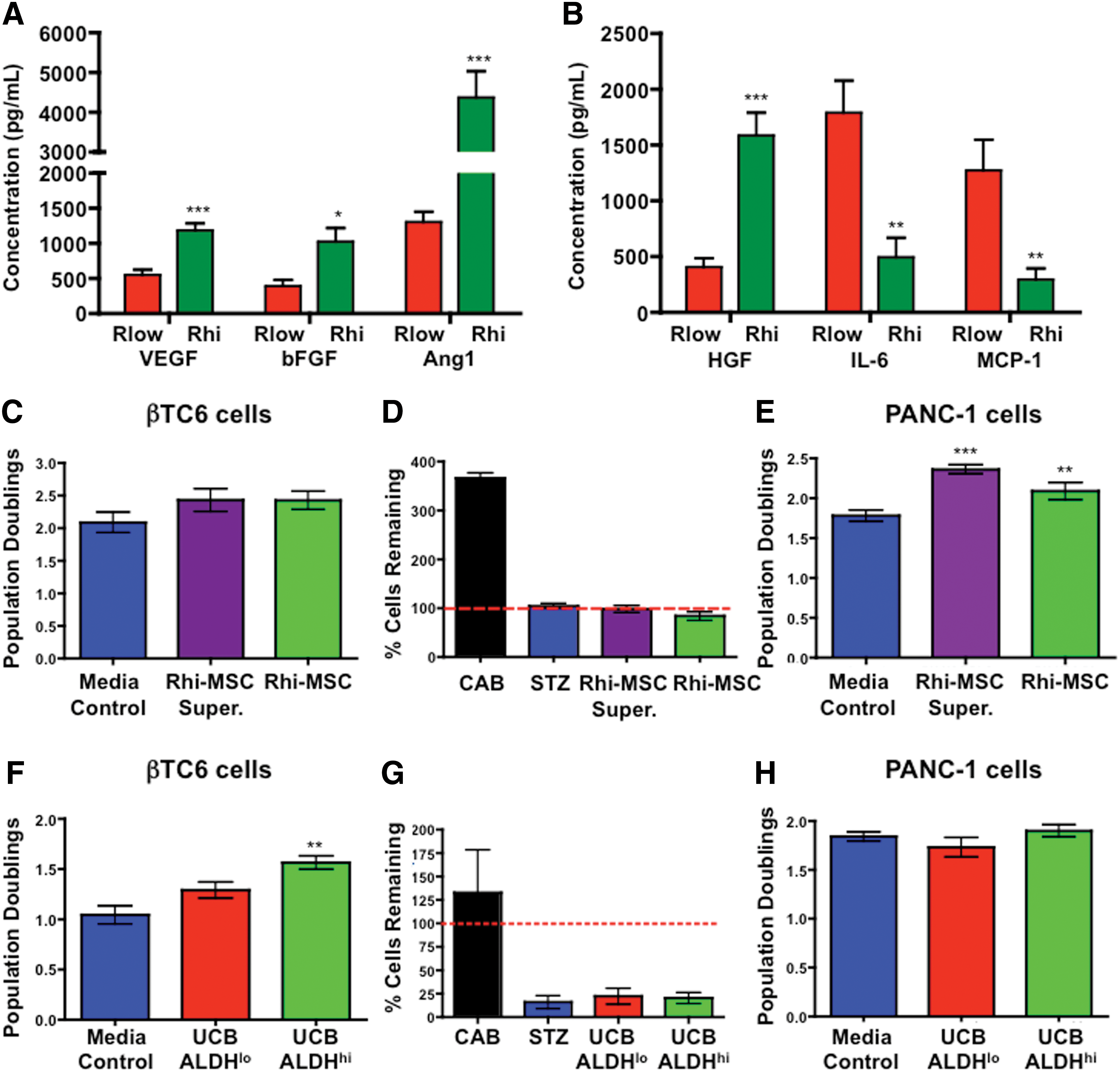
Islet-regenerative MSCs secreted pro-angiogenic cytokines and increased ductal epithelial cell (PANC-1) but not beta cell (βTC6) proliferation in vitro. Supernatant was collected at P4 from Rhi-MSCs (green, n=3) and Rlow-MSCs (red, n=3), and analyzed for the concentrations of pro-angiogenic versus pro-inflammatory proteins by multiplex ELISA.
Rhi-MSCs or UCB ALDHhi cells stimulated the proliferation ductal epithelial (PANC-1) or beta cell (βTC6), respectively
Co-culture of βTC6 cells or PANC1 cells with Rlo-MSCs or Rhi-MSCs was performed to investigate the effects of MSC-secreted stimuli on the proliferation of beta cells or ductal epithelial cells, respectively. Exposure to Rhi-MSC supernatant or co-culture with Rhi-MSCs in hanging wells increased PANC-1 cell proliferation, but did not increase βTC6 cell proliferation or survival after STZ treatment (Fig. 2C–E). Conversely, co-culture with UCB ALDHhi but not ALDHlo cells stimulated the proliferation of βTC6 cells, but did not affect PANC-1 cell proliferation (Fig. 2G, H). Thus, paracrine signals from islet-regenerative MSCs or UCB ALDHhi cells stimulated the proliferation of ductal or beta cell lineages, respectively.
Combinatorial transplantation of Rhi-MSCs with UCB ALDHhi cells improved systemic hyperglycemia via increased islet size, islet number, and beta cell mass
STZ-treated NOD/SCID mice were transplanted with PBS or Rhi-MSCs (5×105) on day 10, and subsequently received a second transplantation of PBS, sample-matched Rhi-MSCs (2×105), or UCB-derived ALDHlo (Alo, 2×105) or ALDHhi (Ahi, 2×105) cells at day 17. UCB-derived cells were used on day 17, rather than BM-derived cells due to increased UCB availability and the requirement for timed transplantation. Injection dates were staggered by 7 days to permit an initial wave of MSC-stimulated islet regeneration (Fig. 1A), followed by the delivery of potentially pro-angiogenic stimuli provided by uncultured ALDHhi progenitors [26,29]. Compared with PBS controls, all mice that received Rhi-MSC transplantation at day 10 showed a decrease in blood glucose by day 17 (Fig. 3A), confirming the immediate glucose-lowering capacity of a single injection of Rhi-MSCs. Surprisingly, mice intravenously injected with ALDHlo cells at day 17 (primarily nongranular leukocytes [27]) quickly returned to hyperglycemic levels similar to PBS controls. In contrast, mice injected with UCB ALDHhi cells or sample-matched Rhi-MSCs maintained lower glucose concentrations until day 42 (Fig. 3A). MSC/Ahi mice also demonstrated a second phase of hyperglycemic reduction between days 28–35, resulting in blood glucose levels significantly lower than PBS/PBS controls (***P<0.001). Indeed, MSC/Ahi mice showed blood glucose concentrations that were lower at day 42 than at day 10 (*P<0.05), indicating that MSC/Ahi combinatorial transplantation not only stabilized glucose levels but reversed established hyperglycemia.

Combinatorial transplantation of islet-regenerative MSCs with UCB ALDHhi cells improved systemic hyperglycemia, glucose tolerance, and serum insulin via increased islet number, islet size, and total beta cell mass.
Glucose tolerance was performed at day 42 to address whether transplanted mice were responsive to glucose challenge. Compared with normoglycemic (5.1±1.4 mmol/L) control mice, where blood glucose peaked at 15 min (11.5±2.7 mmol/L) and returned to basal levels within 60 min (6.2±0.8 mmol/L), STZ-treated MSC/Ahi mice showed moderately increased fasted blood glucose levels (10.5±5.7 mmol/L) that peaked at 30 min (21.6±9.2 mmol/L) and gradually returned to baseline within 120 min (13.4±7.2 mmol/L). MSC/MSC mice followed similar kinetics (Fig. 3B). In contrast, mice transplanted with PBS/PBS or MSC/Alo spiked and maintained maximal glucose levels (33 mmol/L) for>120 min (Fig. 3B). The glucose-lowering kinetics, combined with increased serum insulin at day 42 (Fig. 3C), indicated that the MSC/Ahi mice demonstrated significant, although partial, recovery of endocrine function in response to glucose challenge.
To address tissue engraftment in relation to hyperglycemic recovery in vivo, flow cytometry was performed on day 42 for human-specific HLA-A,B,C (nucleated cells) and CD45 (hematopoietic cells) in the BM, spleen, liver, and pancreas of transplanted mice. Similar to previous observations [24,26], MSC/MSC or MSC/Alo combinatorial transplantation did not result in detectable human cell engraftment in examined tissues (Supplementary Fig. S2A–D). In contrast, MSC/Ahi mice demonstrated human cell engraftment in the BM and spleen, without detection of engraftment in the pancreas at day 42. However, low levels of HLA-A,B,C+/CD45+ cell engraftment were detected in the pancreas at day 28 (0.5%±0.15%, n=3) in MSC/Ahi transplanted mice only. These data suggested that human progenitor cells exert glycemia-lowering effects without permanent residence in the pancreas, presumably by paracrine activities.
Insulin staining of sectioned pancreas was used to assess the recovery of islet size and number after combinatorial transplantation. Compared with PBS/PBS controls, MSC/Ahi-transplanted mice showed increased islet size (Fig. 3D), islet number (Fig. 3E), and total beta cell mass (Fig. 3F). Importantly, MSC/MSC-transplanted mice showed an increased islet number without an increase islet size, confirming our previous observations that transplanted MSCs induced small beta cell cluster formation [26]. MSC/Alo-transplanted mice showed abrogated islet recovery, with no change in islet size or number (Fig. 3D–F). Overall, these data confirmed the potential of transplanted Rhi-MSCs to increase islet number, while UCB-derived ALDHhi cells also increased islet size, resulting in improved glycemic control.
Combinatorial transplantation of Rhi-MSCs with UCB ALDHhi cells increased islet vascularization
We have previously shown that transplantation of BM-derived ALDHhi cells increased islet-specific vascularization [26]. Here, we used immunofluorescent staining for insulin combined with vWF (marking large vessels; Fig. 4A–D) or CD31 (marking intra-islet capillaries; Fig. 4E–H) to show that only MSC/Ahi-transplanted mice showed increased vWF+ vessels associated with islets (Fig. 4I), and CD31+ capillary density within islets (Fig. 4J). These data implicated a role for UCB ALDHhi progenitors increasing insulin delivery to the circulation via the stimulation of islet-specific vascularization. Proliferating cells within the pancreas were pulse-labeled with EdU 24 h before sacrifice (Supplementary Fig. S3A–D). Quantification of EdU+ cells within islets at day 42 was low (≈1%) for all transplanted cohorts and was not significantly increased after combinatorial transplantation (Supplementary Fig. S3A–F).
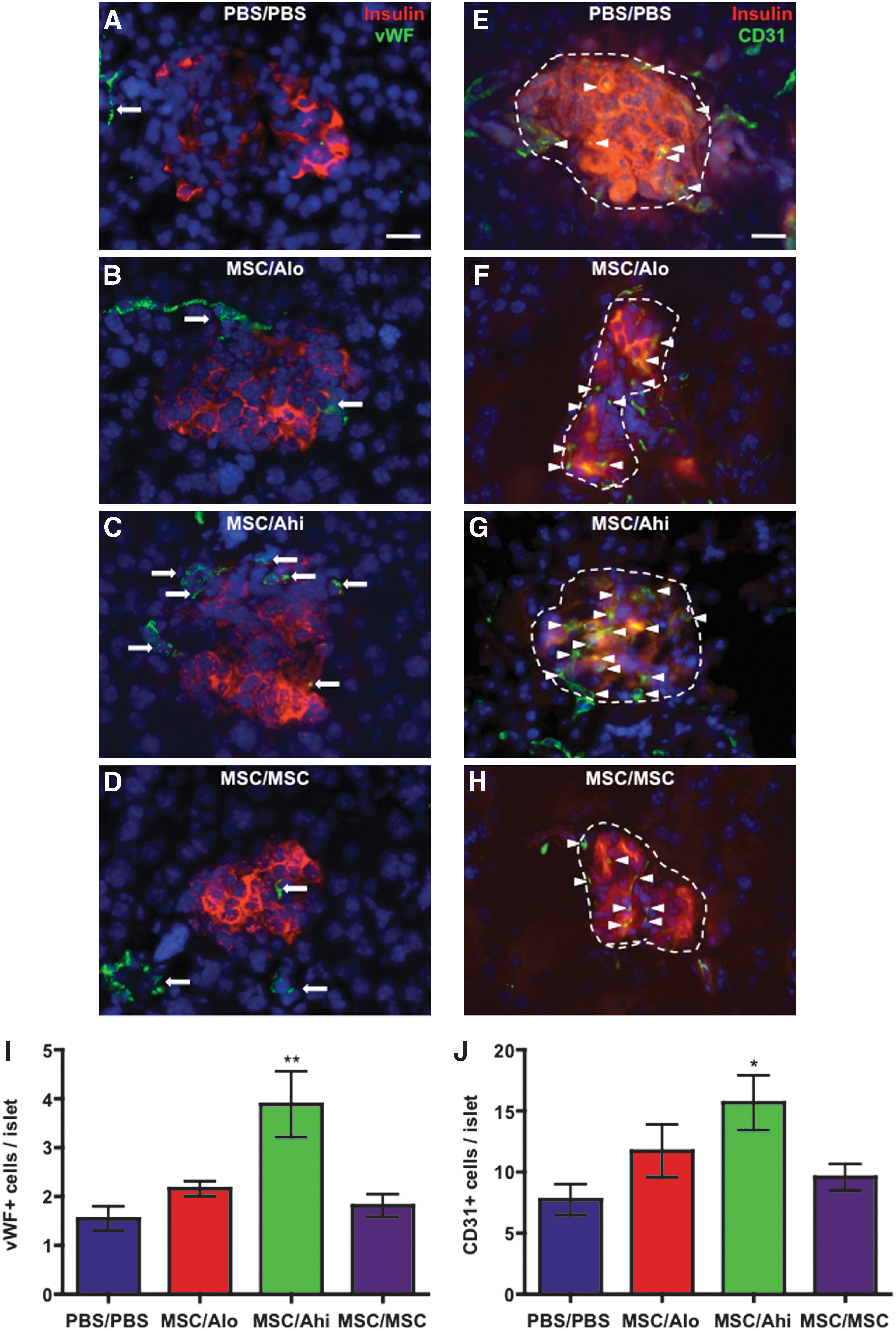
Combinatorial transplantation of islet-regenerative MSCs with UCB ALDHhi cells increased islet vascularization. Representative photomicrographs of pancreas sections stained for insulin (red), and vWF (green,
Combinatorial transplantation of Rhi-MSCs with UCB-derived ALDHhi cells increased islet association with CK19+ ductal epithelium
Previously, MSC transplantation was shown to increase the number of small beta cell clusters associated with the ductal epithelium, suggesting the ductal epithelial niche as a site for MSC-induced islet regeneration [26]. Co-staining for insulin and CK19 to mark ductal epithelial structures (Fig. 5A–D), we confirmed that MSC/Ahi or MSC/MSC transplanted mice had an increased frequency of islets associated with the ductal epithelium (Fig. 5E). Although MSC/Alo-transplanted mice received regenerative MSCs, islet association with ducts was not increased. Interestingly, 32.2%±7.5% of islets in MSC/Ahi mice and 24.5±6.1% of islets in MSC/MSC mice contained CK19+ cells, compared with 8.0%±2.9% of islets in PBS/PBS mice (Fig. 5F). These data further implicate the CK19+ ductal epithelium as a niche for islet regeneration after combinatorial progenitor cell transplantation.
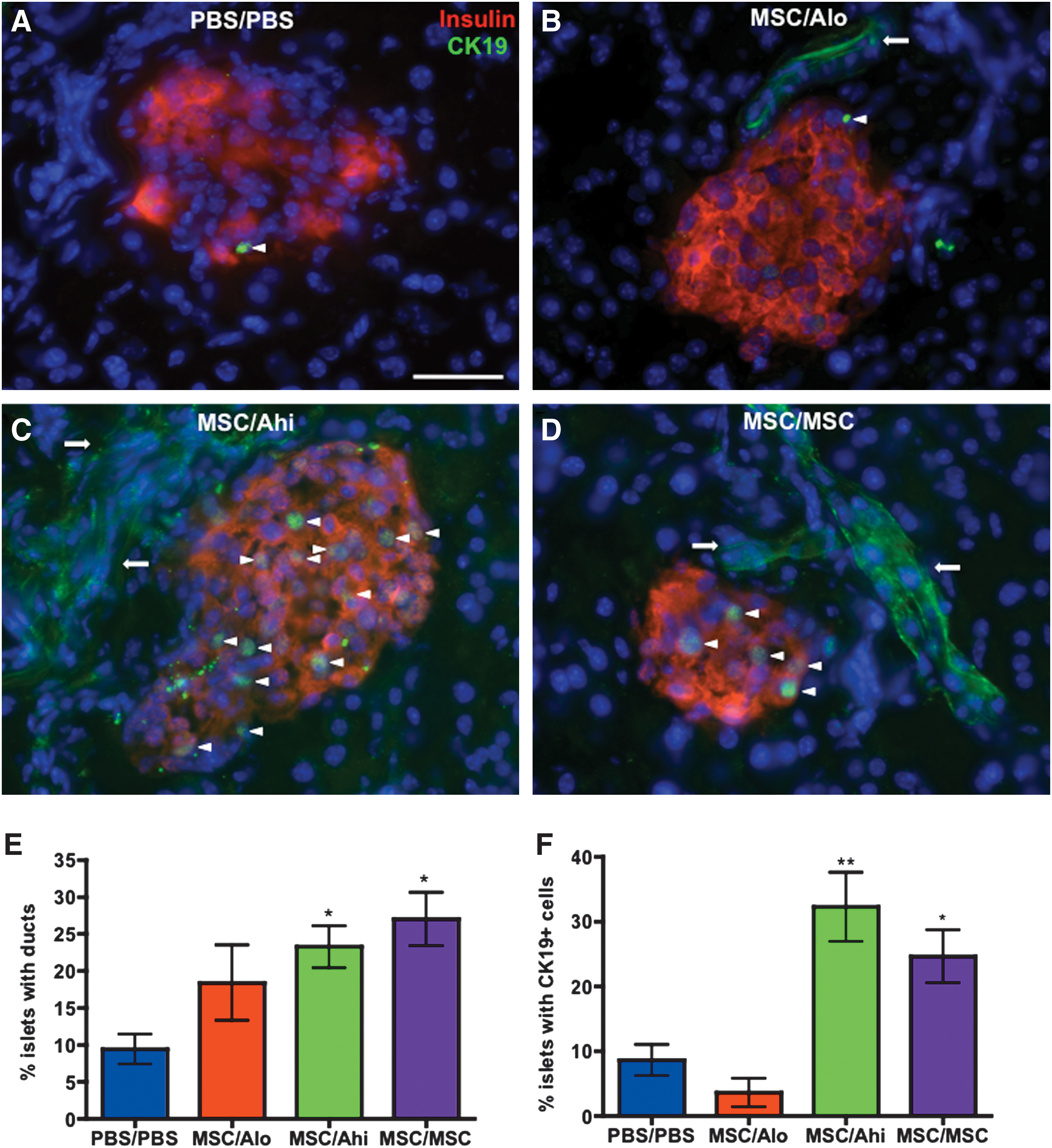
Combinatorial transplantation of islet-regenerative MSCs with UCB ALDHhi cells increased islet association with ducts and CK19+ cells within islets. Representative photomicrographs of pancreas sections stained for insulin (red) and CK19 (green) in mice transplanted with
Discussion
This study demonstrates that islet regeneration can be optimized via the sequential administration of distinct progenitor subtypes from human BM or UCB sources. We have previously shown that neoislet formation was not supported by transplantation of BM-derived ALDHhi progenitors, and increased islet perfusion was not mediated by culture-expanded MSCs, resulting in suboptimal glycemic control after administration of either cell type alone [26]. In contrast, sequential infusion of BM MSCs with UCB ALDHhi cells reduced hyperglycemia, augmented glucose-stimulated insulin secretion, and increased both islet vascularization and beta cell mass associated with the ductal epithelium. Sequential injection of MSCs at days 10 and 17 did not improve islet vascularization or serum insulin in vivo, indicating that transplanted UCB ALDHhi progenitors conferred pro-angiogenic functions that improved systemic insulin release. Thus, we propose a model whereby MSC/Ahi combinatorial transplantation stimulated MSC-mediated islet formation in ductal regions, and UCB ALDHhi cell-mediated neoislet vascularization (Fig. 6).

Endogenous islet regeneration after combinatorial transplantation of islet-regenerative MSCs with UCB ALDHhi cells. Schematic of the mechanisms involved in islet regeneration after progenitor cell transplantation. Transplantation of islet-regenerative MSCs augments the formation of small beta cell clusters associated with CK19+ ductal epithelium. Transplantation of ALDHhi progenitor cells augmented islet size and vascularization through increased beta cell and endothelial cell proliferation, but did not increase islet number. Combinatorial transplantation of islet-regenerative MSCs with UCB ALDHhi cells increased islet size, islet number, total beta cell mass, and increased dynamic control of blood glucose via the sequential induction of islet formation in ductal regions and the stimulation of neoislet revascularization, respectively. Color images available online at
Although human cell engraftment after combinatorial transplantation of MSC/ALDHhi cells was not detected in the pancreas at day 42, HLA-A,B,C+/CD45+ ALDHhi cells were detected at day 28, indicating transient engraftment of UCB ALDHhi cells in the pancreas. Our group and others have also previously shown that transplanted human MSCs can augment islet regeneration after engraftment at ectopic sites [24,26]. Furthermore, circulating progeny of human ALDHhi progenitor cells significantly perfuse the pancreas microvasculature in both ductal and islet regions [35], and transiently engraft the pancreas within the first 7 days of transplantation [26]. Because local paracrine and contact-dependent activities are likely to improve islet regeneration, intrapancreatic-delivery or pro-survival strategies after transplantation warrants further investigation to permit direct and prolonged delivery of progenitor-specific regenerative or pro-angiogenic stimuli.
Rhi-MSC supernatant stimulated the proliferation of ductal epithelial but not beta cell lines in vitro. In contrast, co-culture with UCB ALDHhi cells stimulated beta cell but not epithelial cell proliferation. Although ductal and beta cell lines were used in these experiments, co-culture with primary murine ductal epithelial or beta cells would more accurately reflect the interaction between transplanted human progenitor cells and resident murine pancreatic cells, leading to islet regeneration in vivo. Furthermore, MSC-mediated islet regeneration was associated with CK19+ ductal structures and an increased proportion of CK19+ cells were found within regenerating islets. Although these studies implicated the ductal epithelium as a niche for MSC-mediated regeneration, the signal-receiving cells that mediate islet neogenesis remains a topic of intense debate [12,36 –38]. Recently, Smukler et al. have demonstrated that the human adult pancreas contain rare multipotent progenitors (PMPs) with extensive proliferative and self-renewal capacities [39]. Interestingly, PMPs did not demonstrate a mature beta cell phenotype, as evidenced by low insulin and Glut-2 expression [39]. Although our data implicate the ductal epithelium as a potential intermediate in MSC-induced islet regeneration, we cannot rule out the possibility that islet-derived PMPs may evade Glut-2-mediated STZ toxicity, and are subsequently activated by progenitor-specific paracrine stimuli that promote islet regeneration.
This study also demonstrated that the transcriptional and secretory activities of human MSCs, prospectively screened for the capacity to reduce hyperglycemia after transplantation, could be used to identify pathways implicated in MSC-induced islet regeneration. Rhi-MSCs highly expressed the MMPs, ligands of the EGFR superfamily (AREG, EREG, EPGN, and HBEGF) [40 –42], modulators of the transforming growth factor beta (TGF-β)/activin availability (FBN2), and matrix modifiers downstream of Wnt-signaling (WISP1 and SFRP1) [43]. AREG and EREG were also highly expressed by UCB ALDHhi progenitors. Although Rhi-MSCs also showed increased secretion of pro-angiogenic proteins, and decreased secretion of inflammatory cytokines, intravenously transplanted Rhi-MSCs did not home significantly to the damaged pancreas, and did not increase islet vascularization, suggesting that pro-angiogenic support by MSCs may be required directly at the site of injury to optimize islet vascular repair [44]. These findings underscore the inherent complexity of the islet regenerative niche, and highlight the importance of understanding the microenvironmental modifications by specific progenitor subsets to maximize endogenous islet regeneration.
The EGFR superfamily has long been implicated in the regulation of pancreatic beta cell mass [42], and EGF and gastrin have been shown to promote islet function in mice and humans [45]. Furthermore, EGFR signaling was essential for beta cell mass expansion in response to high-fat diet or pregnancy [40], but was not required during islet neogenesis after ductal ligation [13,40]. Although the EGFR represents a pleiotropic target for drug development, the identified EGF ligands (amphiregulin and epiregulin) represent specific effectors that warrant further investigation in the context of progenitor cell-induced islet regeneration.
The landmark protocols developed by D'amour et al. and Kroon et al. [6 –8] have firmly established the role of activin/TGF-β signaling during endoderm specification by pluripotent cells. Recently, the Keller group has elucidated temporal requirements for TGF-β and Wnt signaling to enhance pancreatic lineage development [46]. Our data suggest that local modulation of matrix TGF-β and bone morphogenetic protein bioavailability by fibrillin-2 [47,48] or WISP-1 [49] may also be implicated in MSC-induced islet regeneration. Importantly, the immediate kinetics of MSC-induced hyperglycemic recovery and the appropriate response of regenerated islets to glucose challenge suggest that Rhi-MSCs may be investigated as feeder cells to efficiently direct beta cell differentiation from both hESC-derived or pancreas-intrinsic endocrine precursors [39,50].
In summary, combinatorial transplantation with BM and UCB progenitor subsets represents a clinically applicable strategy for the development of cellular therapies to treat diabetes. However, further investigation is warranted to understand the paracrine contributions of transplanted progenitor cells for islet-regenerative peptide and drug development. Mechanistic elucidation of the progenitor cells and signals that formulate an islet regenerative niche is also required to maximize the endogenous regenerative potential of endocrine pancreas for clinical applications.
Footnotes
Acknowledgments
This work was supported the Canadian Institutes of Health Research (MOP#86702), and a New Investigator Award from the Heart and Stroke Foundation of Canada to D.A. Hess. We acknowledge Dr. Cheryle Seguin for critical review of the article, David Carter for analysis of microarray data, and Dr. Kristin Chadwick for cell sorting.
Author Disclosure Statement
All authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.