
Abstract
Muscle satellite cells are largely responsible for skeletal muscle regeneration following injury. Side population (SP) cells, which are thought to be muscle stem cells, also contribute to muscle regeneration. SP cells exhibit high mesenchymal potential, and are a possible cell source for therapy of muscular dystrophy. However, the mechanism by which muscle SP cells are committed to differentiation is poorly understood. microRNAs (miRNAs) play key roles in modulating a variety of cellular processes through repression of their mRNA targets. In skeletal muscle, miRNAs are known to be involved in myoblast proliferation and differentiation. To investigate mechanisms of SP cell regulation, we profiled miRNA expression in SP cells and main population (MP) cells in muscles using quantitative real-time polymerase chain reaction-based expression assays. We identified a set of miRNAs that was highly expressed in SP cells as compared with MP cells. One miRNA, miR-128a, was elevated in expression in SP cells, but decreased in expression during continued culture in vitro. Overexpression of miR-128a in SP cells resulted in inhibited cell proliferation. The differentiation potential of SP cells was also decreased when miR-128a was overexpressed. MiR-128a was found to regulate the target genes involved in the regulation of adipogenic-, osteogenic- and myogenic genes that include: PPARγ, Runx1, and Pax3. Overexpression of miR-128a suppressed the activity of a luciferase reporter fused to the 3′-untranslated region of each gene. These results demonstrate that miR-128a contributes to the maintenance of the quiescent state, and it regulates cellular differentiation by repressing individual genes in SP cells.
Introduction
S
microRNAs (miRNAs) are small noncoding RNAs that are usually 19–25 nucleotides long and play important roles and widespread functions in suppressing mRNA translation. Suppression of mRNA translation is accomplished by miRNA binding to the 3′-untranslated region (UTR) of the target genes, including many transcriptional factors. miRNAs control numerous pathways related to development or disease, including cardiac and skeletal muscle hypertrophy, heart disease, and muscular dystrophy [17 –19]. Recent studies have shown that temporarily expressed miRNAs and their target genes are highly regulated and responsible for the differentiation of mesoderm mesenchymal cells into other cellular lineages, such as hematopoietic, osteogenic, and myogenic cells [20].
In this study, we profiled miRNAs that were differentially expressed in muscle SP cells compared with main population (MP) cells. One miRNA, miR-128a, was highly expressed in SP cells, and was shown to have a role as regulator of cell proliferation. In addition, when high levels of miR-128a were induced, the high levels inhibited the differentiation of SP cells into adipogenic-, osteogenic- or myogenic cells. PPARγ, Runx1, and Pax3 were identified as direct targets of miR-128a, and were found to increase in expression following continuous culture of SP cells in vitro as miR-128a expression diminished. Overexpression of miR-128a in SP cells resulted in inhibition of the expression levels of these genes, and suppressed the activity of luciferase reporter fused with the 3′UTR of each gene. Taken together, these results indicate that miR-128a may play a key role in maintaining muscle SP cells in the quiescent state and in regulating their differentiation into multiple cell types.
Materials and Methods
Animals
Animals were housed in the Animal Resources Children's Hospital and all procedures were approved by the Children's Hospital Boston Animal Facilities/IACUC protocols. C57BL/6 mice were originally purchased from Jackson Laboratory (Bar Harbor, ME), housed, and bred according to standard procedures.
Isolation of SP cells
Mononuclear cells were prepared from fore- and hind limb muscles of 8- to 12-week-old male C57BL/6 mice. In brief, muscles were digested with 1.2 U/mL dispaseII (Worthington Biochemical, Lakewood, NJ) and 5 mg/mL collagenase IV (Worthington) for 60 min at 37°C. After lysis of red blood cells (RBCs) by RBC lysis solution (Qiagen, Hilden, Germany), cells were resuspended at 106 cells/mL in Dulbecco's modified Eagle's medium (DMEM; Cellgro, Manassas, VA) containing 2% fetal bovine serum (FBS; Atlanta Biologicals, Lawrenceville, GA) and 12.5 μg/mL Hoechst 33342 (Sigma, St. Louis, MO), then incubated for 60 min at 37°C in either the presence or absence of 50 μM Verapamil (Sigma). They were also stained with FITC-conjugated anti-CD31 antibody (1:200; BD Pharmingen, Franklin Lakes, NJ) and PE-conjugated anti-Sca1 (1:200; BD Pharmingen). Propidium iodide (PI) was added to samples at a concentration of 2 μg/mL to detect and exclude dead cells. Cells were detected and sorted with a FACSVantage flow cytometer (BD Bioscience, Franklin Lakes, NJ).
Cell proliferation and differentiation
For the expansion and proliferation assay, SP cells were cultured in growth medium consisting of DMEM containing 20% FBS, 2.5 ng/mL basic fibroblast growth factor (bFGF; Invitrogen, Carlsbad, CA), and penicillin (100 U/mL)-streptomycin (100 mg/mL; Gibco-BRL, Gaithersburg, MD) on culture dishes coated with Matrigel (BD Biosciences San Diego, CA).
For adipogenic differentiation, cells were cultured for 5 days in adipogenic induction medium that consisted of DMEM with 10% FBS, 0.5 mM isobutylmethylxanthine (Sigma), 0.25 mM dexamethasone (Sigma), and 10 mg/mL insulin (Sigma). The DMEM was then replaced with 10% FBS and 10 mg/mL insulin for 3 days. For osteogenic differentiation, cells were cultured with DMEM containing 5% horse serum and 500 ng/mL mouse bone morphogenetic protein (BMP) 7 (R&D Systems, Inc., Minneapolis, MN) for 5 days [21]. For myogenic differentiation, 1×104 of SP cells were cocultured with the same number of myoblasts in differentiation medium consisting of DMEM with 5% horse serum for 3 days. Myoblasts were isolated from C57BL/6 mice as described [22], by using a CD106 (Vcam1; BioLegend, San Diego, CA) antibody and expanded in vitro in DMEM containing 20% FBS and 2.5 ng/mL bFGF (Invitrogen) prior to coculture.
RNA extraction, real-time polymerase chain reaction-based miRNA array, and TaqMan miRNA expression assay
Total RNAs containing miRNA were extracted from SP cells, MP cells, and cultured SP cells using a miRNeasy RNA isolation kit (Qiagen). The miRNA expression profiling was performed with 50 ng of enriched miRNA using Stem cell miRNA quantitative real-time polymerase chain reaction (qPCR) array (System Biosciences, Mountain View, CA) according to the manufacturer's instructions. This assay contains a panel of 95 stem cell-related and well-established mature miRNA probes, and the U6 transcript as a normalization signal. TaqMan miRNA expression assays were performed by using TaqMan miRNA reverse transcription kit and TaqMan microRNA kit (Applied Biosystems, Foster City, CA) according to the manufacturer's instructions. The expression of miRNA was normalized against the expression of U6, and levels measured by comparative Ct (ΔΔCt) method [23].
Quantitative real-time PCR
First strand cDNA was synthesized with 100 ng of total RNA using a QuantiTect Reverse Transcription Kit (Qiagen). The expression levels of PPARγ, Runx1, Pax3 mRNA, and 18S rRNA were quantified using a SYBR Green PCR Master Mix on an ABI 7900HT real-time PCR machine (Applied Biosystems) following the manufacturer's instructions. Primer sequences for real-time PCR are listed in Supplementary Table S1 (Supplementary Data are available online at
Lentiviral miRNA overexpression and inhibition
Lentivirus-based expression plasmids containing green fluorescent protein (GFP) that overexpress pre-miR-128a, express an antimiR-128 (miRZip) and miRNA control, were purchased from System Biosciences. Lentiviral vectors, along with packaging plasmids (MDL/RRE, Rev, and VSV-G), were transfected into HEK 293T cells using Lipofectamine 2000 (Invitrogen). Three days after transfection, viral supernatants were collected and filtered through 45 μm filters (VWR, West Chester, PA), mixed with Lenti-X lentivirus concentrator (Clonetech, Palo Alto, CA), and incubated overnight at 4°C. The following day, the virus coprecipitate was concentrated by centrifugation at 1,500 g for 60 min at 4°C. Viral pellets were resuspended in phosphate-buffered saline. To overexpress or inhibit miRNA expression, the virus was added to the SP cells in culture. Seventy-two hours after induction, GFP (+) miR-128a overexpressed or inhibited SP cells were collected based on GFP expression by flow cytometer and the cell size was analyzed with CellQuest (BD Biosciences).
Cell cycle analysis by flow cytometry
Cells were collected by trypsinization and centrifugation and resuspended at 106 cells/mL in DMEM (Cellgro) containing 2% FBS (Atlanta Biologicals). Cells were stained with 5 μg/mL Hoechst 33342 (Sigma) for 45 min at 37°C and then analyzed with a FACSVantage flow cytometer (BD Bioscience). The cell cycle was analyzed with Flow Jo software (TreeStar, Inc., Ashland, OR).
Cytochemistry and histochemistry
Cultured cells were fixed on 8-well Lab-Tek Chamber Slides (Nunc, Rochester, NY). To stain lipids, cells were fixed in 10% formalin, rinsed with water and 60% isopropanol, and then stained with Oil red O (Sigma) in 60% isopropanol. To measure the extent of adipogenic differentiation, stained oil droplets were extracted with 4% Nonidet P-40 in isopropanol, and the absorbance of the extract was assayed at 520 nm. Alkaline phosphatase (ALP) activity was detected by the Alkaline Phosphatase Substrate Kit III (Vector Laboratories, Inc., Burlingame, CA), and then the number of ALP-positive cell was counted. After staining, cells in plates are trypsinized and then counted with Countess Automated Cell Counter (Invitrogen). To determine the adipogenic or osteogenic differentiation index, the absorbance or the number of ALP-positive cells was normalized to the total number cells after staining. To stain myotubes, cells were fixed in 4% paraformaldehyde for 5 min, stained with mouse anti-Myosin Heavy Chain (1:10; clone: MF20; Developmental Studies Hybridoma Bank, Iowa City, IA), at 4°C overnight and then incubated with the secondary antibody conjugated with Alexa 568 (Molecular Probes, Eugene, OR). Nuclei were stained with 4,6-diamidino-2-phenylindole (DAPI). The fusion indexes were calculated as the percentage of nuclei in Myosin Heavy Chain-positive myocyte or myotubes per total nuclei in the dish.
Luciferase assay
The 3′UTR of the mouse PPARγ, Runx1, and Pax3 genes were amplified by PCR and cloned into a modified version of the pGL2Basic vector (containing a novel multi-cloning site between the luciferase open-reading frame and the SV40 polyA signal; gift from Dr. J.A. Kreidberg) [24] using the following sequence-specific primers: PPARγ, 5′-AATTACGCGTCTATTATTTTGAGGGAAAAA-3′ and 3′-AATTAAGCTTACAATATGCATAAAATAGATCA-5′, Runx1, 5′-AAT TGTTTAAACAGGGCAGCCATAGCAACT-3′ and 5′-AATTAAGCTTCACTAAAT GGCTCTAACAC-3′, and Pax3, 5′-AATTCTCGAGTAGAAACACATGTTCTTAAT- 3′ and 5′-AATTGAATTCATTCTTCATGTCTAGTCTGT-3′. The predicted miR-128a binding sites located in the 3′UTR were mutated using the QuikChange II Site-Directed Mutagenesis Kit (Strategene, La Jolla, CA) and the following primers: PPARγ, 5′-CTGACACCTAAGAAATTTCCGGGGAAAAAGCATTTAAAAAC-3′ and 3′-GT TTTTAAATGCTTTTTCCCCGGAAATTTCTTAGGTGTCAG-5′, Runx1, 5′-ATCC CGGAGGGAACCGGGGAATGCTTCTGA-3′ and 3′- TCAGAAGCATTCCCCGGT TCCCTCCGGGAT-5′, and Pax3, 5′-GCCCCTAAATTTTAATTGTCCC-3′ and 3′-GGGACAATTAAAATTTAGGGGC-5′. For luciferase reporter assay, HEK293T 3×104 cells were initially plated in a 48-well plate 24 h prior to transfection. The cells were transfected using Lipofectamine 2000 (Invitrogen) with 30 ng of reporter construct and 100 ng of miRNA overexpression plasmids or scrambled miRNA controls (Origene, Rockville, MD). Luciferase activity was measured 48 h after transfection with the Dual Reporter Assay System (Promega, Madison, WI) on a single-tube luminometer (Berthold Technologies, Bad Wildbad, Germany). Transfections were performed in triplicate, and the individual experiments were repeated 3 times.
Statistics
All quantitative data are represented as mean±standard error. Analysis was performed between different groups using 2-tailed student's t-test and nonparametric Mann–Whitney U test. A probability of less than 5% (P<0.05) was considered statistically significant.
Results
miRNA expression comparison between SP cells and MP cells
Real-time PCR-based miRNA expression array was performed on isolated SP cells and MP cells. SP cells were defined as the cell fraction that efficiently effluxes Hoechst 33342 dye and represent 0.1% of muscle mononuclear cells (Fig. 1A). Verapamil, which is a ATP-binding cassette transporter inhibitor, was able to block Hoechst dye efflux from SP cells and was used to define the SP population (Fig. 1A). Prior to qPCR array, SP cells and MP cells were subfractionaized via cell surface markers, since it is known that muscle SP and MP cells are heterogeneous. As observed by others, most SP cells express both CD31 and Sca-1, the former as a marker of endothelial cells and the latter of hematopoietic progenitor cells and muscle stem cells recognition [25], while MP cells were more heterogeneous (Fig. 1A, B). MP cells consisted of 64.9%±1.5% of CD31(+) Sca-1(+) cells, 14.4%±3.2% of CD31(−) Sca-1(−) cells, 17.6%±4.0% of CD31(−) Sca-1(+) cells, and 2.0%±0.2% of CD31(−) Sca-1(−) cells (Fig. 1C). For miRNA profiling, total SP cells and CD31(+) Sca-1(+) MP cells were collected in parallel. To investigate the miRNAs that may regulate the lineage potential of SP cells, miRNA profiling was performed using a commercial qPCR array plate, which assays 95 stem cell-related and well-established mature miRNAs using U6 as a normalization signal.
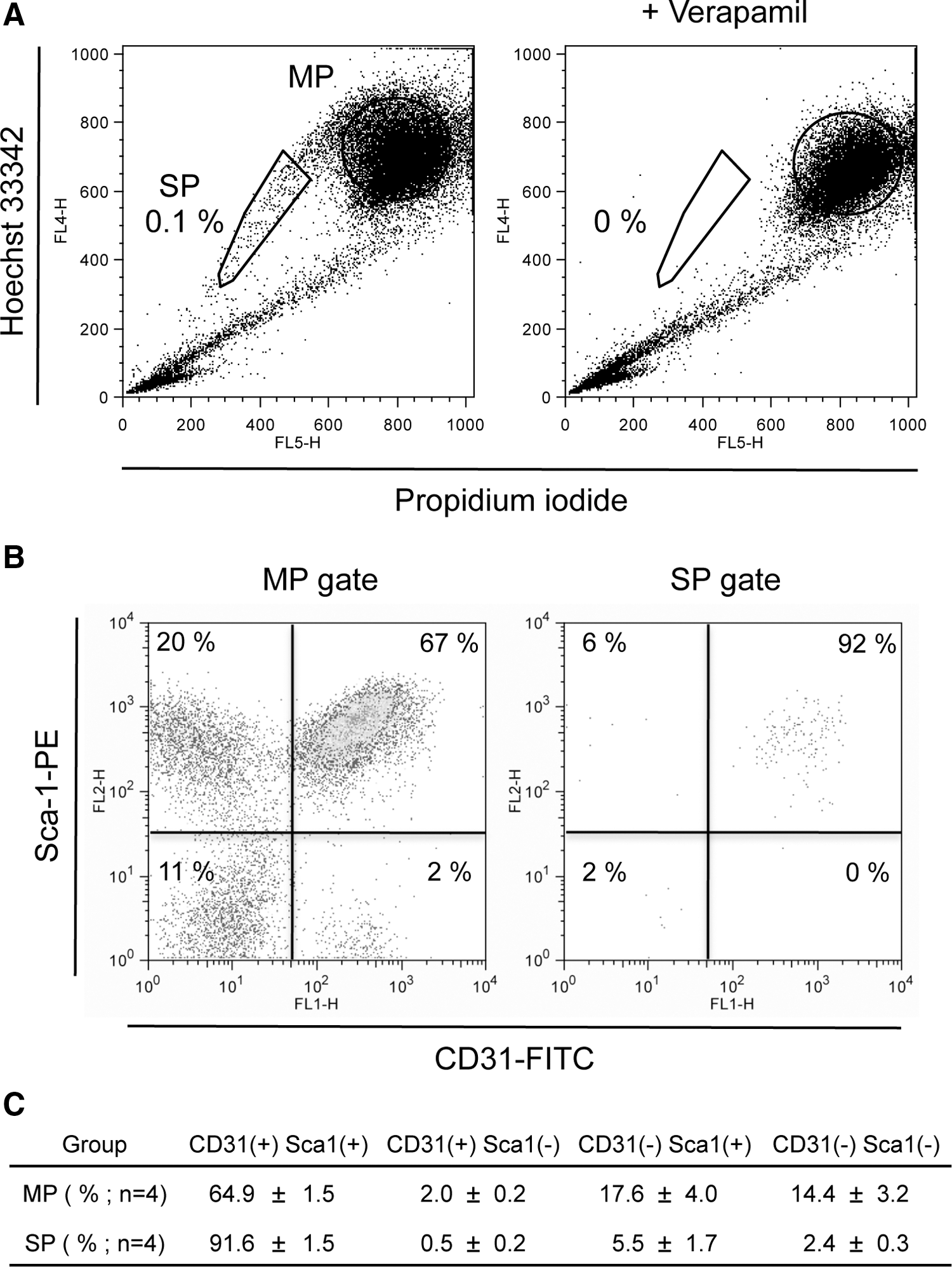
Characterization of muscle main population (MP) and side population (SP) cells.
miRNAs array was performed using a SYBR green-based qPCR, and miRNAs were identified to be expressed at higher levels in SP cells than in CD31(+) Sca-1(+) MP cells (Table 1A), and those that were expressed at lower levels in SP cells relative to their expression levels in CD31(+) Sca-1(+) MP cells (Table 1B). Array data were verified by quantitative real-time TaqMan PCR assay and fold change was confirmed. The miRNA expression array revealed that miR-128a was the most highly expressed in SP cells compared with CD31(+) Sca-1(+) MP cells, and this increased expression was confirmed by TaqMan qPCR assay (Fig. 2A).

Changes of microRNA-128a expression in SP cells compared with CD31(+) Sca-1(+) MP cells.
The quantitative real-time polymerase chain reaction-based miRNA expression array was performed to compare SP cells with MP cells as described in Materials and Methods. Data show the 10 miRNAs that were most highly expressed in SP cells relative CD31(+) Sca-1(+) MP cells (A) or in CD31(+) Sca-1(+) MP cells relative to SP cells (B).
SP, side population; MP, main population; miRNA, microRNA.
MiR-128a regulates the cell compartment, cell size, and proliferation of SP cells
To begin to address the function of miRNA128a in SP cells, its expression was analyzed following culture of SP cells in vitro. SP cells were sorted based on Hoechst dye exclusion and, prior to culture in vitro, were largely Ki67-negative, which has been shown to be a marker of mitosis and cell proliferation [25]. Upon culturing of SP cells there were increased numbers of Ki67-positive cells (data not shown). To determine whether this change in proliferation markers was due to the change of miR-128a expression, the expression of miR-128a in SP cells was analyzed after 7 days of in vitro cell culture. Interestingly, miR-128a expression decreased during culture (Fig. 2B). The correlation between increased expression of Ki67 and decrease in miR-128a suggests that miR-128a may be important to maintain cells in a quiescent state. When miR-128a expression in SP cells is induced or inhibited (anti miR) along with a GFP tag, infected cells can be sorted based on GFP expression by flow cytometer, and necrotic or apoptotic cells excluded by PI staining. To investigate the effect of miR-128a on cell proliferation in SP cells, miR-128a or control (scrambled miR) were introduced in SP cells, and cultured in vitro for 6 days, following which cells were counted. The number of SP cells when miR-128a was overexpressed is significantly lower than that of scrambled miR-SP cells, suggesting that the overexpression of miR-128a resulted in inhibition of cellular proliferation causing a decrease in the number of SP cells compared with scrambled miR (Fig. 3A). To support this result, the expression of cell cycle regulatory genes, p16, p19, p21, and p27, was analyzed by qPCR in SP cells. Consistent with inhibition of cell proliferation, miR-128a overexpression increased the expression of these genes (Supplementary Fig. S1A). This indicates that miR-128a can regulate cellular proliferation of SP cells. On the other hand, when miR-128a was inhibited, SP cells or scrambled miR-SP cells cultured in vitro for 6 days, the suppression of miR-128 promoted cell proliferation by increasing cell numbers (Fig. 3B), and was paralleled by the decrease in expression of cell cycle inhibitory genes (Supplementary Fig. S1B). To further confirm that miR-128a would regulate cell proliferation, cell cycle analysis was performed by FACS in SP cells with overexpression or inhibition of miR-128a. The percentage of cells in G0-G1 phase in cells where miR-128a overexpressed was higher than that in scrambled miR-SP cells, and concomitantly decreased in S phase and G2-M phase (Fig. 3C, D). In contrast, the inhibition of miR-128a exhibited increased percentage of cells in G2-M phase compared with control SP cells (Fig. 3C, D). In addition, the cell size in miR-128a overexpressed or inhibited SP cells was analyzed using FACS, since cell size and protein synthesis is a reflection of the cell proliferation [26,27]. Overexpression in SP cells of miR-128a resulted in decreased cell size by ∼20% as compared with control SP cells when analyzed by FACS (Fig. 4A, B), whereas miR-128 inhibition did not affect cell size (Fig. 4E, F). To confirm that cell size is decreasing by miR-128a, SP cells where miR-128a was overexpressing were stained with Sca-1 and DAPI to visualize the cell outline and nuclei, and then the nuclear to cytoplasm ratio was calculated. As shown in Fig. 4C, overexpression in SP cells of miR-128a resulted in decreased cell size compared with control SP cells.
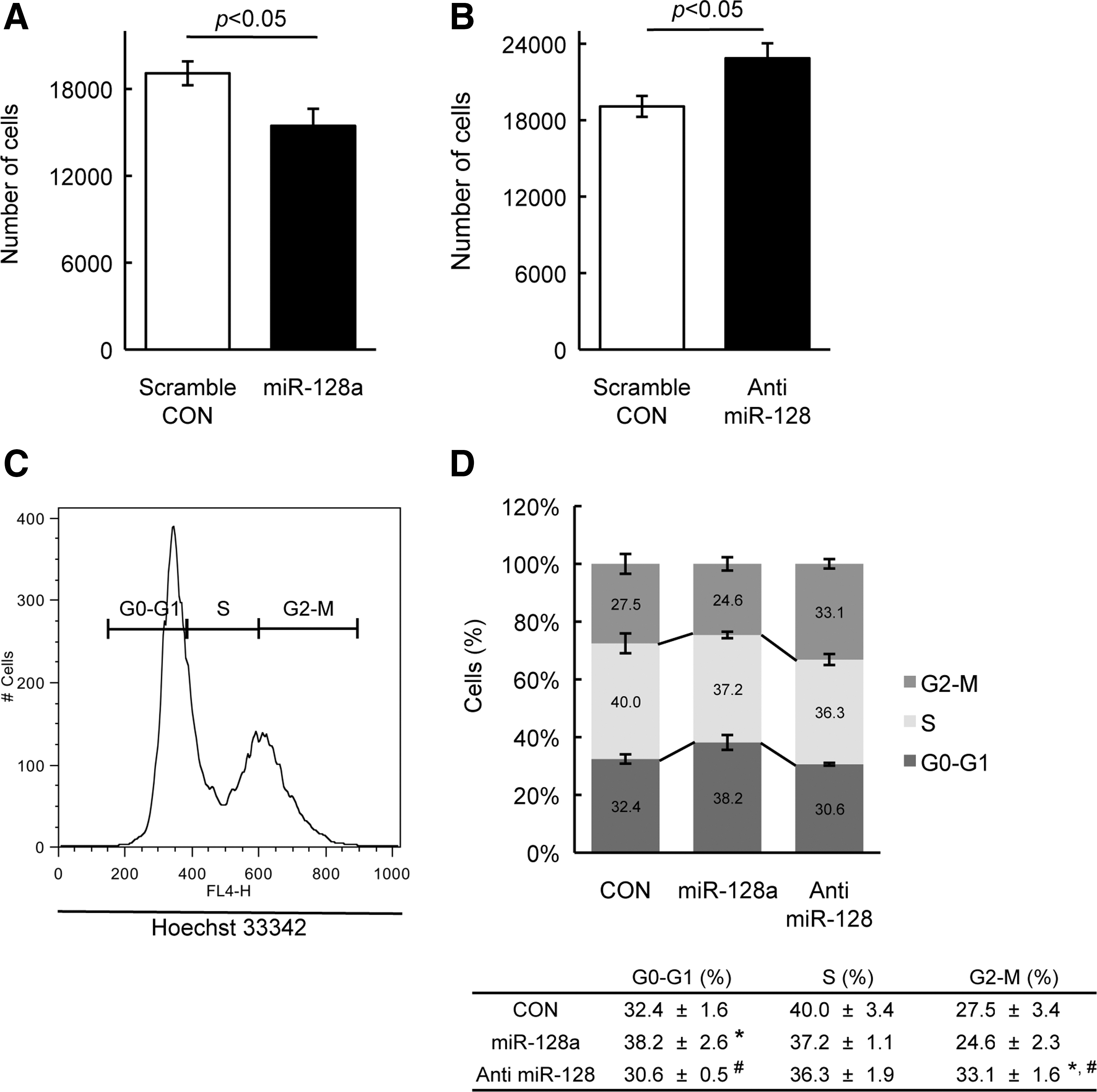
Overexpression of miR-128a affects proliferation of SP cells.
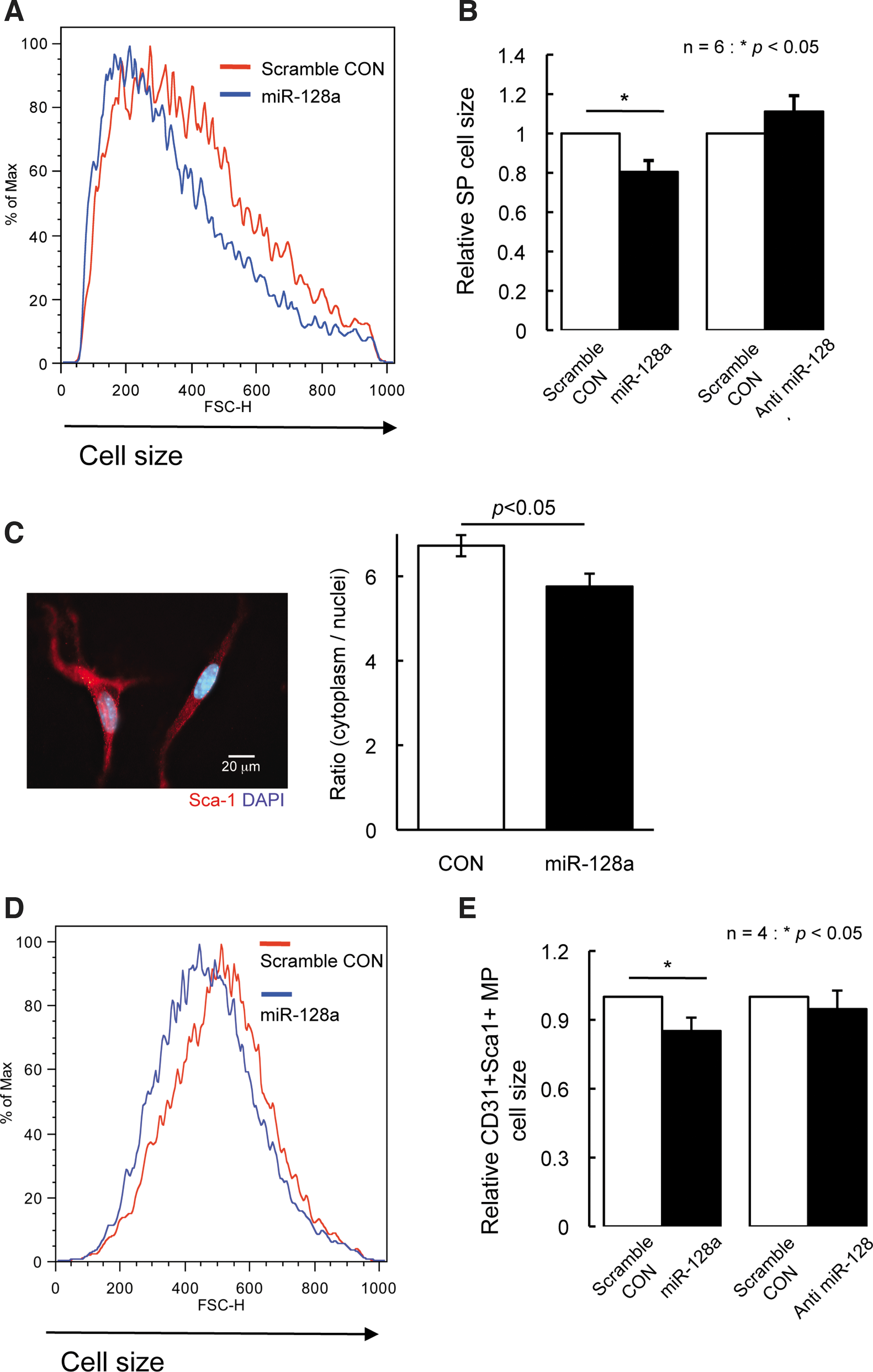
Overexpression of miR-128a affects cell size of SP cells.
We further evaluated the cell size by FACS in CD31(+) Sca-1(+) MP cells if miR-128a would have a same function in MP cells. CD31(+) Sca-1(+) MP cells where miR-128a was overexpressed was smaller compared with control CD31(+) Sca-1(+) MP cells, whereas there was no affect of cell size in CD31(+) Sca-1(+) MP cells where miR-128 was inhibited. Intriguingly, miR-128a did not affect cell size in myoblast from wild-type mice where miR-128a was overexpressed or inhibited (data not shown). These findings revealed that miR-128a could regulate the cell size in SP cells and CD31(+) Sca-1(+) MP cells, and also the effect of miR-128a was different depending on cell types. In summary, our results are suggesting that miR-128a might play an important role of proliferation, and regulate cell size of SP cells.
MiR-128a can regulate osteogenic, adipogenic, and myogenic differentiation in SP cells
Muscle SP cells have been shown to have the potential to differentiate into multi-cell types under different environmental conditions in vivo and in vitro [11,12,25,28]. Interestingly, previous reports indicated that miR-128a is expressed in early progenitor cells and is thought to play a role in regulating lineage specific commitment [29,30].
SP cells can differentiate into adipocyte containing liquid droplets in cytoplasm when cultured with adipogenic differentiation medium [25]. Following adipogenic induction, control-SP cells can differentiate into Oil red O-positive adipocytes, while SP cells overexpressing miR-128a have reduced ability to differentiate into adipocytes (Fig. 5A). To determine whether miR-128a might also regulate osteogenic differentiation, SP cells were cultured with osteoblast differentiation medium containing BMP7 [21]. After induction, ALP activity in SP cells was analyzed, ALP is a marker of osteoblast differentiation. Scrambled miRNA control SP cells, when induced with BMP7, gave rise to ALP-positive cells, whereas SP cells in which miR-128a was overexpressed, had reduced numbers of ALP-positive cells (Fig. 5B). For myogenic differentiation, SP cells need to be cocultured with myoblasts to form myotubes since most SP cells cannot form myotubes by themselves [25]. SP cells in which miR-128a was overexpressed or scrambled miR-SP cells were cocultured with myoblasts for 3 days and then stained with anti-myosin heavy chain. The fusion index (calculated by the number of nuclei in myosin heavy chain-positive cells per total nuclei) of miR-128a overexpressed SP cells was ∼20% lower than that of myoblasts cocultured with control-SP cells, suggesting that the potential of myogenic differentiation was lower in miR-128a overexpressing SP cells than in control SP cells (Fig. 5C, D). These combined findings indicate that miR-128a levels might influence the differentiation of SP cells into different cell lineages.

MiR-128a regulates the cell differentiation potential of SP cells into adipogenic, osteogenic, and myogenic lineages.
PPARγ, Runx1, and Pax3 are regulated by miR-128a in vitro
Computational analysis using prediction algorithms was used to elucidate the potential targets of miR-128a. These included TargetScan (
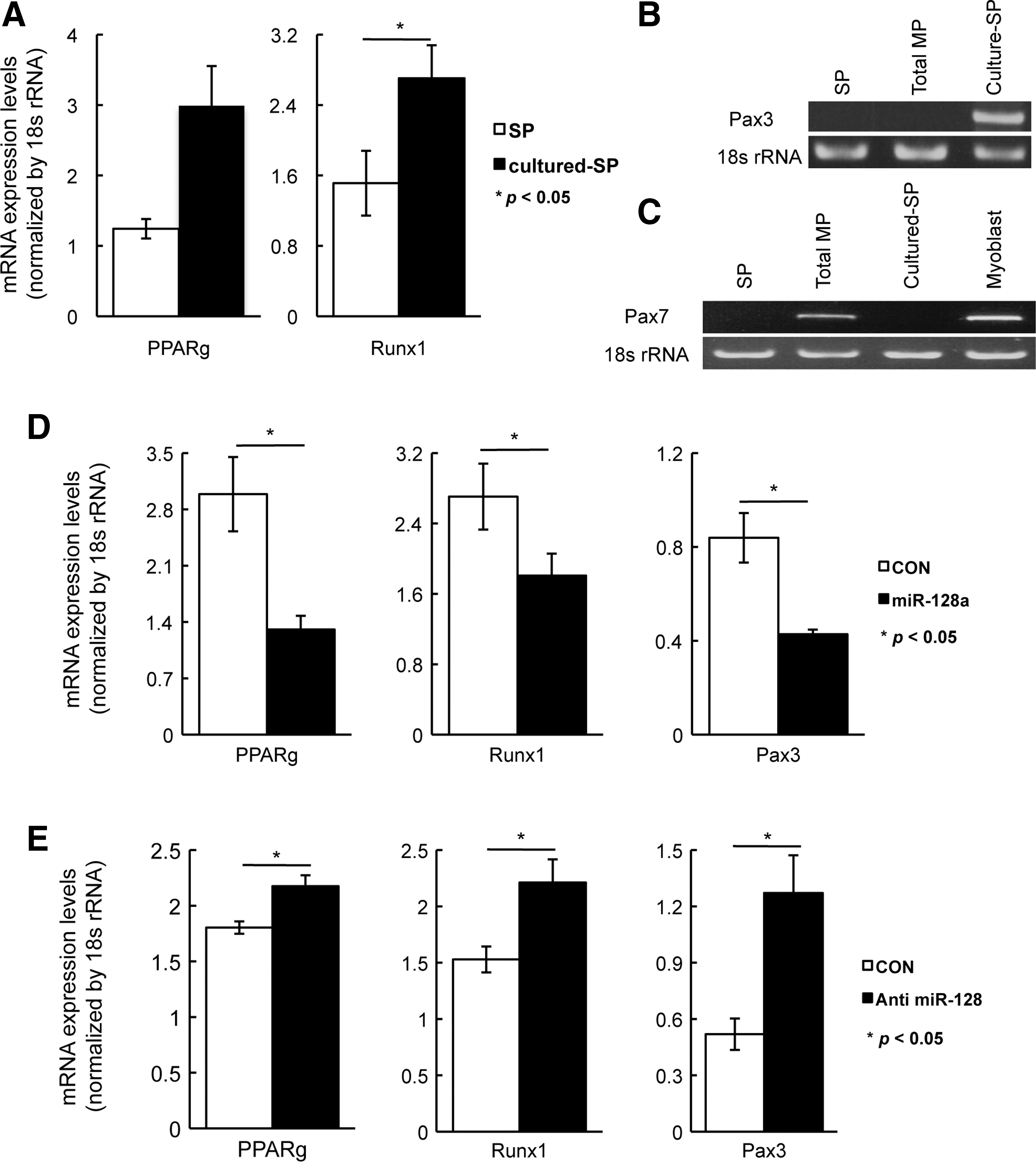
Effects of target gene expression by miR-128a in SP and total MP cells.
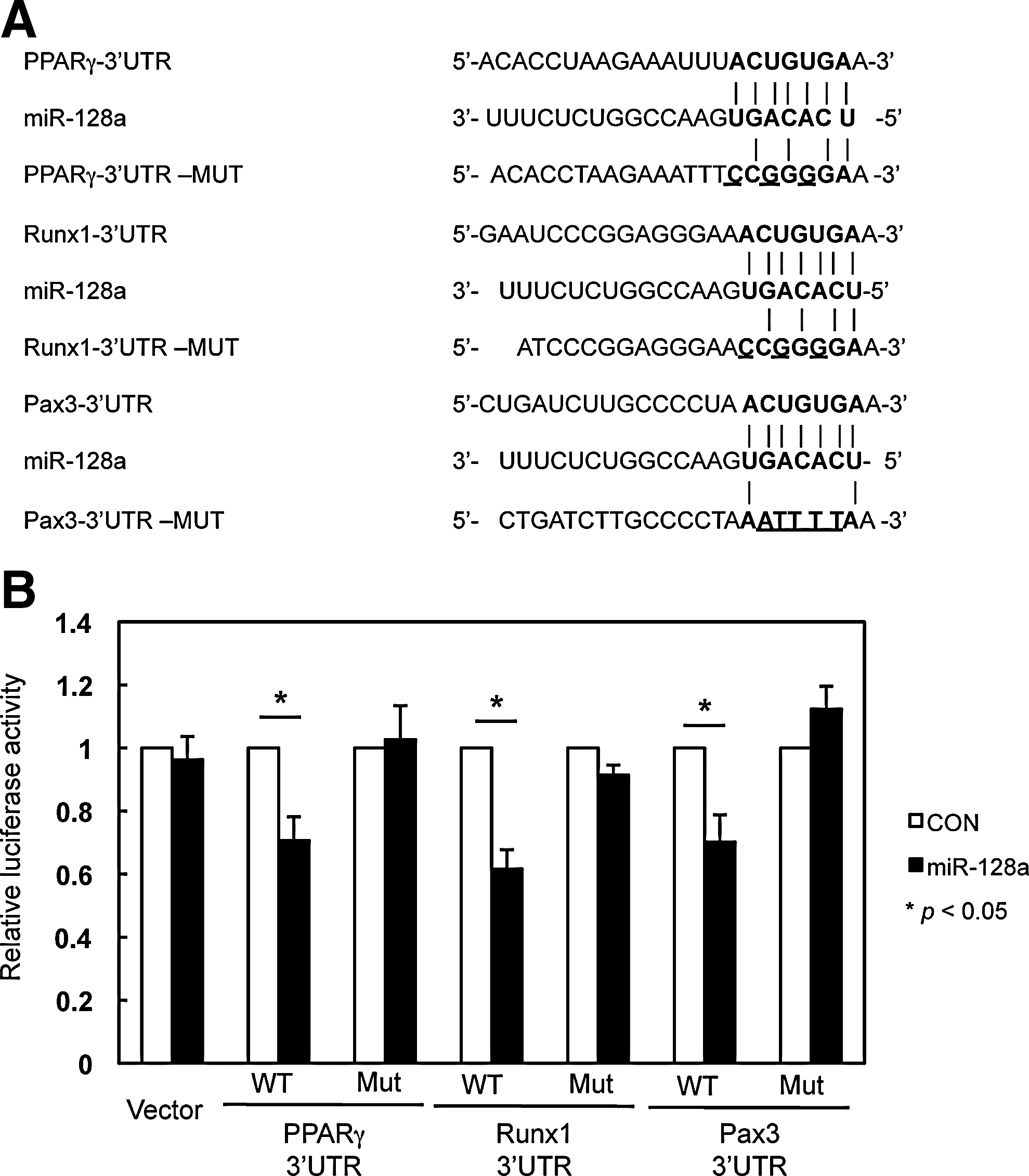
MiR-128a targets the cell differentiation-related genes, PPARγ, Runx1, and Pax3.
Discussion
Mesenchymal stem cells including muscle SP cells have the potential to differentiate into many different cell lineages, such as fat, bone, or muscle, both in vivo and in vitro. Some of this differentiation is likely modulated by miRNA expression, and this should be detectable by profiling miRNA expression in different cell populations. Profiling miRNA expression in muscle SP cells identified those miRNAs that were highly expressed in muscle SP cells. One of them, miR-128a, exhibited increased expression in SP cells compared with MP cells (Table 1A). Although very little is known about the function or targets of miR-128a, the mature miR-128a transcript has been highly expressed in brain and skeletal muscle [33].
Recent studies have reported that miR-128a was detected in some types of tumors such as glioblastoma or medulloblastoma, and that these tumors have been shown to have reduced levels of miR-128a expression, resulting in increase of cell proliferation [30 –32]. These results are consistent with our results presented here that miR-128a could regulate cell proliferation in SP cells. Recent articles have described that the influence of miR-128a on cell cycle progression is through the regulation of the cell cycle-related gene, Bmi-1, which is the target gene of miR-128a [31,32]. Bmi-1 is a polycomb gene that was originally identified as an oncogenic partner of c-Myc in murine lymphomagenesis. It is required for cell proliferation, self-renewal, and maintenance, regulating p16, p19, p21, and p27 as downstream effectors of Bmi-1, in hematopoietic and neural stem cells [34 –37]. Indeed, miR-128a overexpression resulted in an increase of cell cycle-related genes although the decrease of Bmi-1 expression was not significant in SP cells (data not shown). These data indicate that miR-128a can regulate the cell cycle, and other unidentified miR-128a target genes may contribute to cell cycle-related genes in SP cells. According to computational analysis in TargetScan, there are more than 600 predicted target genes for miR-128a, some of which are contained within MAPK signaling pathways (eg, MAPK14, MAP3K12, MAPK8IP3, Rap1b, MAP2K7, and RASA2), or insulin signaling pathways (eg, PDE3A, INSR, IRS1, PIK3R1, PDPK1, and RPS6KB1). These signaling pathways and other potential target genes that may involve cell proliferation should be elucidated in future studies. In addition, cell size and protein synthesis are associated with cell proliferation. The control of protein synthesis and cell size are linked [26,27]; thus, the decreased cell size by miR-128a overexpression with attenuating cell proliferation is likely to support the hypothesis that miR-128a regulates cell proliferation in SP cells.
SP cells have been reported to be able to differentiate into various cell lineages, such as hematopoietic cells, adipogenic cells, osteogenic cells, and myogenic cells in vivo and in vitro [11,12,25]. However, the mechanism through which the fate of SP cells is determined is poorly understood. MiR-128a, as previous reports described, is expressed in early progenitor cells in hematopoietic stem cells and is considered to be a regulator of differentiation into mature hematopoietic cells [29]. In addition, miR-128a also plays an important role in regulating lineage-specific commitment in neuronal stem cells [30]. Altogether, these finding are consistent with a possible role of miR-128a in SP cells in the regulation of cell fate by targeting key regulatory factors.
According to previous studies, the lineage-negative fraction of SP cells is the one with differentiation potential and largely contributes to the differentiation and CD31-positive SP cells have a little potential of differentiation in vitro [25]. In this study, total SP cells, which contain 90% of CD31-positive and about 10% of CD31-negative SP cells, were analyzed. Interestingly, miR-128a expression in CD31-positive SP cells was slightly higher than that in lineage-negative SP cells (Supplementary Fig. S4), suggesting that miR-128a may regulate not only lineage-negative SP cell fraction but also CD31-positive SP cells, and miR-128a have other functions in CD31-positive SP cells. Difference of miRNAs expression in CD31-positive orCD31-negative SP cells and further analysis of the function of miR-128 will be elucidated in the future studies.
Gene expression of PPARγ, Runx1, and Pax3 was shown to be increased in SP cells, following differentiation to adipocyte, osteocyte, or myotube under the different differentiation conditions (Figs. 5 and 6). PPARγ regulates differentiation to adipogenesis by associating with C/EBPa, and is essential for adipocyte differentiation [38 –40]. Runx transcriptional factors, such as Runx1, 2, and 3, are well known to contribute to the regulation of osteogenic differentiation [41 –44]. Although it has been established that Runx1 regulates hematopoiesis [45,46], recent reports indicate that Runx1 is also expressed in osteocytes, and contributes to the early stage of osteogenic differentiation and bone renewal [42,43,47]. Pax3 is typically expressed in muscle progenitor cells prior to the expression of myogenic genes, MyoD and Myf5, and is essential as an activator of these genes in muscle cells [48]. Increasing Pax3 expression in cultured SP cells also may reflect the myogenic potency, as previous studies have demonstrated that cultured SP cells can engraft into dystrophic muscles more efficiently than freshly isolated SP cells [13]. Taken together, elevated individual genes coordinated with decreasing miR-128a are key regulators of cell differentiation, and our results support the role that miR-128a might be able to function as a direct regulator to cell differentiation in SP cells.
However, overexpression of miR-128a could not inhibit individual differentiation completely, most likely due to the heterogeneity of SP cells, or gene regulation by other miRNAs. Although SP cells predominantly express CD31 and Sca-1, SP cell fractions are enriched with mesenchymal stem cells, but are still heterogeneous [15,25,49]. The hypothesis is that miR-128a could regulate a subset of SP cells, which have differentiation potential. Whether miR-128a can regulate differentiation in other cells needs examination in future studies.
To date, many studies have shown and reported that cell fate determination could be controlled by several miRNAs, and miRNAs could induce or inhibit the adipogenic [50 –55], osteogenic [56 –58], or myogenic [59 –62] differentiation in mesenchymal stem cells. Intriguingly, miR-204/211 can inhibit osteogenic differentiation and simultaneously stimulate adipocyte differentiation [63]. A recent article has also shown that miR-138 inhibits mesoderm type lineage differentiation (osteocyte, adipocyte, and chondrocyte), and has a role to maintain undifferentiating state in mesenchymal stem cells [64]. Obviously, miR-128a controlled these (osteogenic, adipogenic, and myogenic) differentiation pathways and had possible target genes in these pathways, suggesting that miR-128a also might be one of those miRNAs that could regulate cell differentiation in SP cells.
In conclusion, we have identified miR-128a as highly expressed in SP cells and a possible regulator of the cell lineage differentiation through targeting of PPARγ, Runx1, and Pax3. MiR-128a may be a regulator of cell fate, since several miRNAs and their target genes are involved in the cellular differentiation. These studies might be useful to understand the regulation of stem cells and to develop new stem cell-based therapies for muscular diseases, such as Duchenne muscular dystrophy.
Footnotes
Acknowledgments
Funding for this work was generously provided by the Bernard F. and Alva B. Gimbel Foundation (L.M.K.) and through an NIH grant (5P50 DK49216). We would like to thank members of the Kunkel laboratory, Dr. Emanuela Gussoni for her helpful advice, Marie Torres and Richard Bennett who are members of the Stem Cell Core Facility at Children's Hospital Boston (supported by National Institutes of Health award NIH-P30-HD18655) for technical support.
Author Disclosure Statement
The authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.