
Abstract
Mesenchmal stem cells (MSCs) can be differentiated into either adipocytes or osteoblasts, and a reciprocal relationship exists between adipogenesis and osteogenesis. Multiple transcription factors and signaling pathways have been reported to regulate adipogenic or osteogenic differentiation, respectively, yet the molecular mechanism underlying the cell fate alteration between adipogenesis and osteogenesis still remains to be illustrated. MicroRNAs are important regulators in diverse biological processes by repressing protein expression of their targets. Here, miR-22 was found to regulate adipogenic and osteogenic differentiation of human adipose tissue-derived mesenchymal stem cells (hADMSCs) in opposite directions. Our data showed that miR-22 decreased during the process of adipogenic differentiation but increased during osteogenic differentiation. On one hand, overexpression of miR-22 in hADMSCs could inhibit lipid droplets accumulation and repress the expression of adipogenic transcription factors and adipogenic-specific genes. On the other hand, enhanced alkaline phosphatase activity and matrix mineralization, as well as increased expression of osteo-specific genes, indicated a positive role of miR-22 in regulating osteogenic differentiation. Target databases prediction and validation by Dual Luciferase Reporter Assay, western blot, and real-time polymerase chain reaction identified histone deacetylase 6 (HDAC6) as a direct downstream target of miR-22 in hADMSCs. Inhibition of endogenous HDAC6 by small-interfering RNAs suppressed adipogenesis and stimulated osteogenesis, consistent with the effect of miR-22 overexpression in hADMSCs. Together, our results suggested that miR-22 acted as a critical regulator of balance between adipogenic and osteogenic differentiation of hADMSCs by repressing its target HDAC6.
Introduction
A
The balance between adipocyte formation and osteobalst formation is associated with metabolic regulation and homeostasis maintaining in human tissue. Once the balance between adipogenesis and osteogenesis is disrupted, various metabolic-related disorders and developmental diseases will occur. For example, osteoporosis or age-related bone loss is usually accompanied with a significant increase of adipocytes accumulation [7], and patients with high bone mass phenotype exhibit increased bone mineral density but reduced fat tissue volume in bone marrow [8]. Therefore, investigation of the balance between adipogenic and osteogenic differentiation is of significant importance for elucidation of pathogenesis and development of therapies for bone-related diseases.
MicroRNAs (miRNAs), a group of endogenously expressed small noncoding RNA molecules that are involved in post-transcriptional regulation of their targets via protein transcription repression or mRNA degradation, have been shown to play an important role in diverse biological processes [9,10]. The effect of miRNAs has also been investigated in either adipogenesis or osteogenesis. For instance, miR-27a, miR-27b, and miR-130 inhibit adipocyte formation by targeting peroxisome proliferator-activated receptor gamma (PPARγ), the key adipogenic transcriptional factor, while miR-21 facilitates adipogenic differentiation by repressing TGFBR2 as well as TGF-β signaling in hADMSCs [11 –14]. miR-20a has recently been reported as a positive regulator of osteogenic differentiation of hADMSCs through coregulating the bone morphogenetic protein signaling pathway [15], while miR-23a acts as an osteogenic suppressor by inhibiting runt-related transcription factor 2 (Runx2), the key transcriptional factor of osteogenic differentiation, in its 3′UTR [16].
Although many miRNAs have been identified to regulate adipogenesis or osteogenesis, only a few of them have been implicated to have a role in both processes. Previously, we reported that miR-138 could inhibit adipogenic differentiation of hADMSCs by targeting EID-1 [17]. Months later, another study confirmed its effect as a negative regulator in osteoblast formation through the FAK-ERK1/2 signaling pathway [18]. Besides, miR-204 functioned as an enhancer of adipogenesis but an inhibitor of osteogenesis by repressing Runx2 in MSCs [19]. Moreover, miR-637 and its target osterix (OSX) were demonstrated to maintain the balance between adipogenic and osteogenic differentiation by promoting adipocyte formation and suppressing osteoblast formation [20].
In the present study, miR-22 was found to decrease during adipogenic differentiation but increase in the process of osteogenic differentiation, indicating the dual impact of miR-22 in regulating adipo/osteogenesis of hADMSCs. By transfection of miR-22 mimics into hADMSCs, we found that osteogenic differentiation was promoted, while adipogenic differentiation was significantly suppressed due to miR-22 overexpression. Histone deacetylase 6 (HDAC6), a corepressor of Runx2 [21], was then identified as a target of miR-22. Silencing endogenous HDAC6 expression in hADMSCs exhibited enhanced osteogenesis but repressed adipogenesis. Taken together, our findings provided a novel functional and mechanical link between miR-22 and HDAC6 in regulating the balance between adipogenesis and osteogenesis of hADMSCs.
Materials and Methods
Isolation and culture of hADMSCs
hADMSCs were isolated from human adipose tissue as previously described [22]. Human adipose tissue was collected from 23- to 25-year-old healthy women performing liposuction surgery according to procedures approved by the Ethics Committee at the Chinese Academy of Medical Sciences and Peking Union Medical College. hADMSCs were then resuspended at a density of 3×106 in a 15-mL growth medium [17] and cultured in a 75-cm2 flask after isolation from human adipose tissue. Cell cultures were maintained at 37°C in a humidified incubator with 5% CO2.
Adipogenic and osteogenic differentiation of hADMSCs
Adipogenic differentiation was induced in high glucose of Dulbecco's modified Eagle's medium (H-DMEM) supplemented with 10% fetal bovine serum (FBS), 1 μM dexamethasone, 0.5 mM 3-isobutyl-1-methylxanthine, and 5 μg/mL 0.1 mM l-ascorbic acid. After culturing in an adipocyte induction medium for 4 days, the medium was replaced with an adipocyte-maintaining medium, H-DMEM with 10% FBS. Oil red O staining was performed to visualize lipid droplets in adipocytes as previously described [17].
For osteogenic differentiation, hADMSCs were cultured in H-DMEM containing 10% FBS, 10 mM β-glycerophosphate, 50 μM l-ascorbic acid, and 0.01 μM dexamethasone. The osteoblast phenotype was indicated by alkaline phosphatase (ALP) activity determination, an early marker of osteogenic differentiation. Von Kossa Staining was performed to detect matrix mineralization deposition at the late stage of bone formation (cells were washed with phosphate-buffered saline, fixed in 4% paraformaldehyde for 1 h, incubated with 5% Na2S2O3 in 37°C for 30 min, stained by 1% AgNO3 with UV irradiation for 20 min, and washed gently with water).
Transfection of miRNA mimics and small-interfering RNAs
The synthetic miR-22 mimics and miRNA-negative control (miR-NC) were purchased from GenePharma. Small-interfering RNAs (siRNA) of HDAC6 (siHDAC6) and their negative control (siNC) were purchased from Invitrogen. About 50 nM miRNA mimics and 40 nM siRNA with siPORT™ NeoFX™ Transfection Agent (AM-4511, Ambion; Applied Biosystems) were transfected in the OPTI-MEM Reduced Serum Medium (Gibco) according to the manufacturer's instructions.
Real-time polymerase chain reaction analysis
Total RNA was extracted with TRIzol (Invitrogen), and cDNA was prepared as described [17,23]. Real-time polymerase chain reaction (PCR) was amplified in triplicates according to procedures reported previously [17]. Relative expression of mRNA or miRNA was evaluated by the 2−ΔΔCt method and normalized to the expression of GAPDH or U6, respectively. Primers of mRNA real-time PCR analysis are listed in Table 1.
ALP, alkaline phosphatase; HDAC6, histone deacetylase 6; PPARγ, peroxisome proliferator-activated receptor gamma; OSX, osterix; LPL, lipoprotein lipase; OPN, osteopontin; OCN, osteocalcin; IBSP, integrin-binding sialoprotein; FABP4, fatty acid binding protein 4.
Western blot analysis
Proteins were extracted with radioimmunoprecipitation (RIPA) lysis buffer with PMSF, quantified by BCA Protein Assay Kit, separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis gel, and then transferred onto a polyvinylidene fluoride membrane. The membrane was incubated with primary antibody, anti-HDAC6 (1:2,000; 07-732; Millipore), or anti-β-actin (1:5,000; sc-47778; Santa Cruz) overnight at 4°C, and then incubated with HRP-conjugated anti-rabbit (sc-2004; Santa Curz) or anti-mouse (sc-2005; Santa Cruz) secondary antibody for 1 h at room temperature. The immunodetection was then visualized by a chemiluminescent ECL reagent (Millipore).
Dual luciferase reporter assay
A 70-bp synthetic fragment of HDAC6 3′UTR containing the predicted seed match site with miR-22, or the mutant site was inserted between the NotI and XbaI cleavage sites of pRL-TK vector (Promega), downstream of the Renilla luciferase reporter gene. Cells were cotransfected with pRL-TK vector with or without the synthetic fragment of HDAC6 3′UTR, pGL-3 vector with Firefly luciferase reporter gene, and 50 nM miR-22 mimics or miR-NC by lipofectamine 2000 (Invitrogen) according to the manufacturer's instructions. Luciferase activity was measured in triplicates for 36 h after transfection by using the Dual Luciferase Reporter Assay System (E1910; Promega). Renilla luciferase activity was normalized to firefly luciferase activity.
miRNA target prediction
Prediction of miRNA target genes was relied on databases TargetScan (
Statistical analysis
Data are presented as mean±SD. Statistical analysis was performed by 2-tailed t-test using SPSS Statistics 17.0. Differences with P<0.05 were considered statistically significant.
Results
Expression level of miR-22 during adipogenic and osteogenic differentiation of hADMSCs
To gain a comprehensive understanding of the roles miRNAs play in the balance between adipogenesis and osteogenesis of hADMSCs, we performed microarray-based miRNA expression profiling during both adipogenic and osteogenic differentiation of hADMSCs and searched for miRNAs with an inverse expression pattern between adipogenesis and osteogenesis. Among these miRNAs, miR-22 was shown to decrease in adipo-differentiated hADMSCs while increase in osteo-differentiated hADMSCs according to the microarray analysis (unpublished data), suggesting its role in the regulation of adipo/osteogenic differentiation. We then performed real-time PCR to examine endogenous expression of miR-22 in hADMSCs during the process of osteogenic and adipogenic differentiation. RNA samples of hADMSCs were harvested every 4 days after induction into osteogenic differentiation, as shown in Fig. 1A; the expression level of miR-22 was upregulated during osteogenic differentiation in hADMSCs. In contrast, compared with the undifferentiated control group (day 0), the expression of miR-22 was downregulated by >70% at day 4 and maintained lower than the undifferentiated group during adipogenic differentiation of hADMSCs (Fig. 1B). These results indicated that miR-22 might play a role in regulating osteogenic and adipogenic differentiation of hADMSCs.
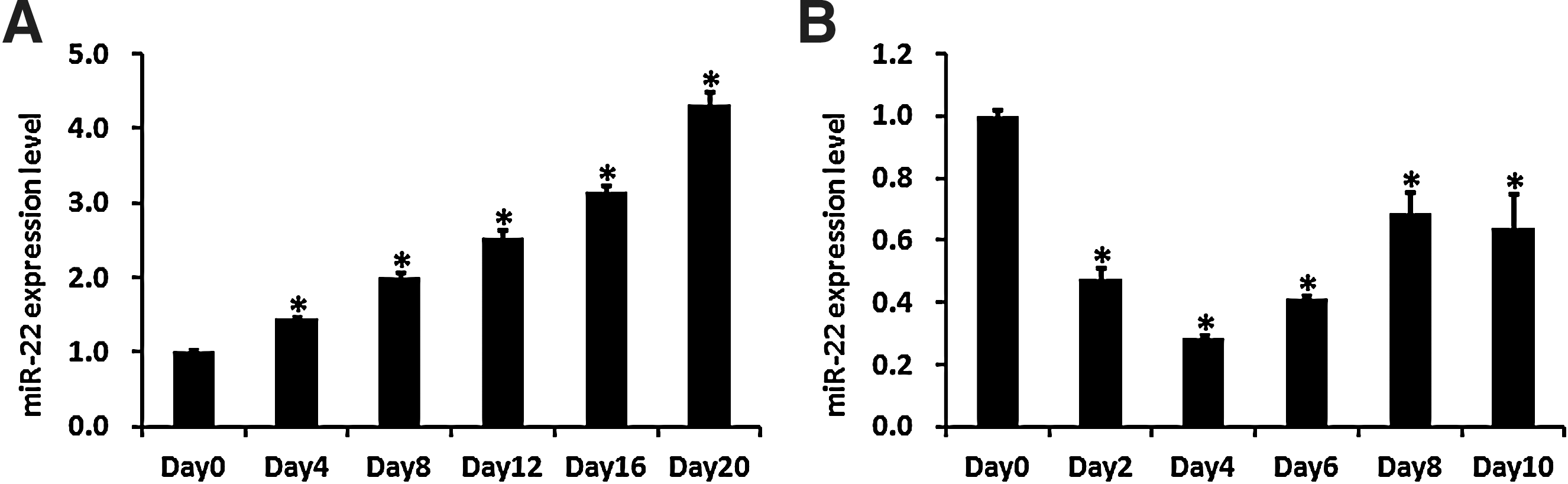
miR-22 is upregulated during osteogenic differentiation and downregulated during adipogenic differentiation of hADMSCs. The endogenous expression level of miR-22 was measured by real-time PCR at different time points during
Effect of miR-22 on adipogenic and osteogenic differentiation of hADMSCs
To investigate the biological function of miR-22 on adipogenic and osteogenic differentiation of hADMSCs, synthetic miR-22 mimics were transfected into hADMSCs, and the transfection efficiency was evaluated by a fluorescent microscope (Fig. 2A). Analysis of real-time PCR after 48-h transfection showed that miR-22 was increased more than 3 times compared with the untreated control (Con) group and the miR-NC trasnfection group (Fig. 2B).
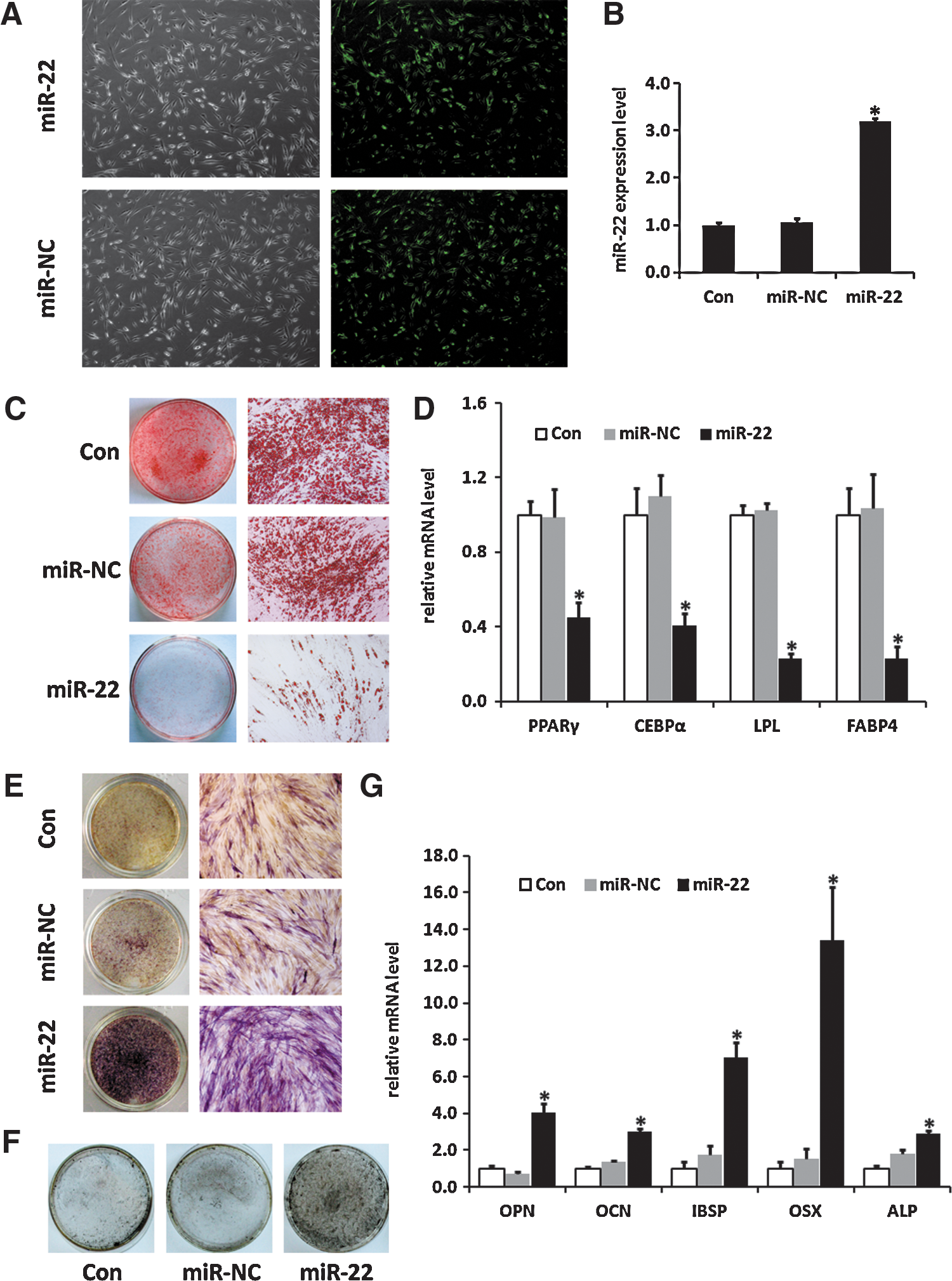
Overexpression of miR-22 inhibits adipogenic differentiation and promotes osteogenic differentiation of hADMSCs.
hADMSCs transfected with miR-22 mimics were then cultured in the adipogenic induction medium. According to the results of Oil red O staining, by day 10 of differentiation, the accumulation of lipid droplets decreased significantly after transfection with miR-22 mimics (Fig. 2C). In addition, mRNA expression levels of adipogenic transcription factors and adipocyte-specific markers were also detected by real-time PCR, as illustrated in Fig. 2D, PPARγ, CCAAT/enhancer-binding protein alpha (C/EBPα), lipoprotein lipase (LPL), and fatty acid binding protein 4 (FABP4) decreased remarkably in cells transfected with miR-22 mimics compared with the control group and the miR-NC-treated group (Fig. 2D). The results above suggested that miR-22 acted as a negative regulator in adipogenic differentiation of hADMSCs.
In parallel, hADMSCs were also induced into osteogenic differentiation after being transfected with miR-22 mimics or miR-NC. Overexpression of miR-22 enhanced osteogenic differentiation, which was indicated by the increase of ALP activity (Fig. 2E), as well as accelerated matrix mineralization visualized by Von Kossa staining (Fig. 2F). Moreover, higher mRNA expression levels of osteo-specific markers were detected due to the increase of miR-22 in hADMSCs (Fig. 2G). Therefore, the data above revealed that miR-22 positively regulated osteogenic differentiation of hADMSCs.
HDAC6 is a direct downstream target of miR-22
To reveal the molecular mechanism underlying the regulation of hADMSCs by miR-22, we searched for potential downstream targets of miR-22 by 2 different miRNA target prediction databases TargetScan and miRanda. Among the targets predicted in both databases, we focused on HDAC6, which belonged to class II of the HDAC family and played a critical role in transcriptional regulation by suppressing the activity of other transcription factors [24]. We chose HDAC6 as the candidate target of miR-22 in the regulation of hADMSCs differentiation because HDAC6 was reported to directly interact with Runx2 and functioned as a corepressor of Runx2 in pre-osteoblasts [21]. Runx2, the key osteogenic transcription factor, was identified as a modulator of the balance between adipogenic and osteogenic differentiation since Runx2 could propel osteogenic differentiation while suppress adipogenic differentiation [19,25]. According to the computational analysis, HDAC6 has a 7-nt seed region, which is complementary to miR-22, within its 3′UTR (Fig. 3A). To investigate whether miR-22 inhibits the expression of HDAC6 through the seed region in its 3′UTR, we inserted a 3′UTR sequence of HDAC6 containing the predicted binding seed site for miR-22 into a dual luciferase construct, pRL-TK, which was then cotransfected with miR-22 mimics to detect HDAC6 protein translation via Renilla luciferase activities. Results showed that miR-22 repressed the luciferase activity through 3′UTR of HDAC6 compared with the miR-NC transfection control group (Fig. 3B). Additionally, miR-22 resulted in no effect on luciferase activity in the group inserted with mutated seed match site in 3′UTR, or the control group without a 3′UTR sequence of HDAC6.
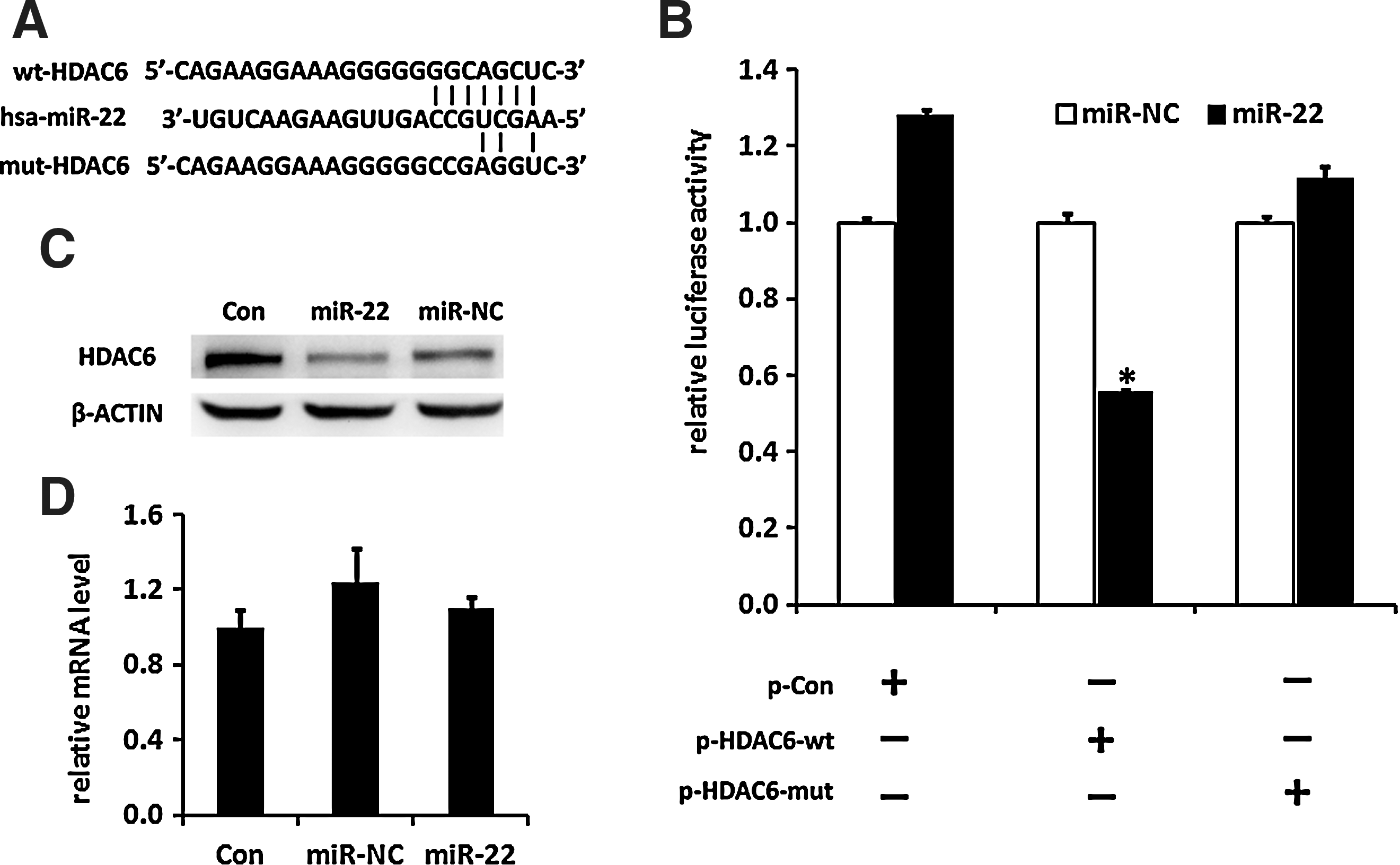
miR-22 inhibits HDAC6 protein expression by targeting its 3′UTR.
To further determine the effect of miR-22 on endogenous HDAC6 expression, protein and RNA samples of hADMSCs were harvested after transfection with miR-22 mimics. As shown in Fig. 3C, increasing miR-22 repressed HDAC6 protein expression in hADMSCs without affecting the level of its mRNA (Fig. 3D). Taken together, these results revealed that miR-22 regulates HDAC6 protein expression through its 3′UTR, indicating that HDAC6 might be involved in the regulation of adipo/osteogenic differentiation of hADMSCs.
Expression level of HDAC6 during adipogenic and osteogenic differentiation of hADMSCs
We then evaluated mRNA and protein expression levels of HDAC6 during the process of adipogenic and osteogenic differentiation of hADMSCs by real-time PCR and western blot. As shown in Fig. 4A, the endogenous mRNA level of HDAC6 increased as hADMSCs were being induced into adipocytes. Analysis of western blot revealed that the protein level of HDAC6 was also upregulated during adipogenic differentiation (Fig. 4B). On the contrary, both mRNA and protein levels of HDAC6 were reduced and maintained at a low level in hADMSCs during the whole process of osteogenic differentiation (Fig. 4C, D). Taken together, the expression kinetics of HDAC6 in hADMSCs suggested that HDAC6 might be a repressor of osteogenic differentiation but a promoter of adipogenic differentiation.
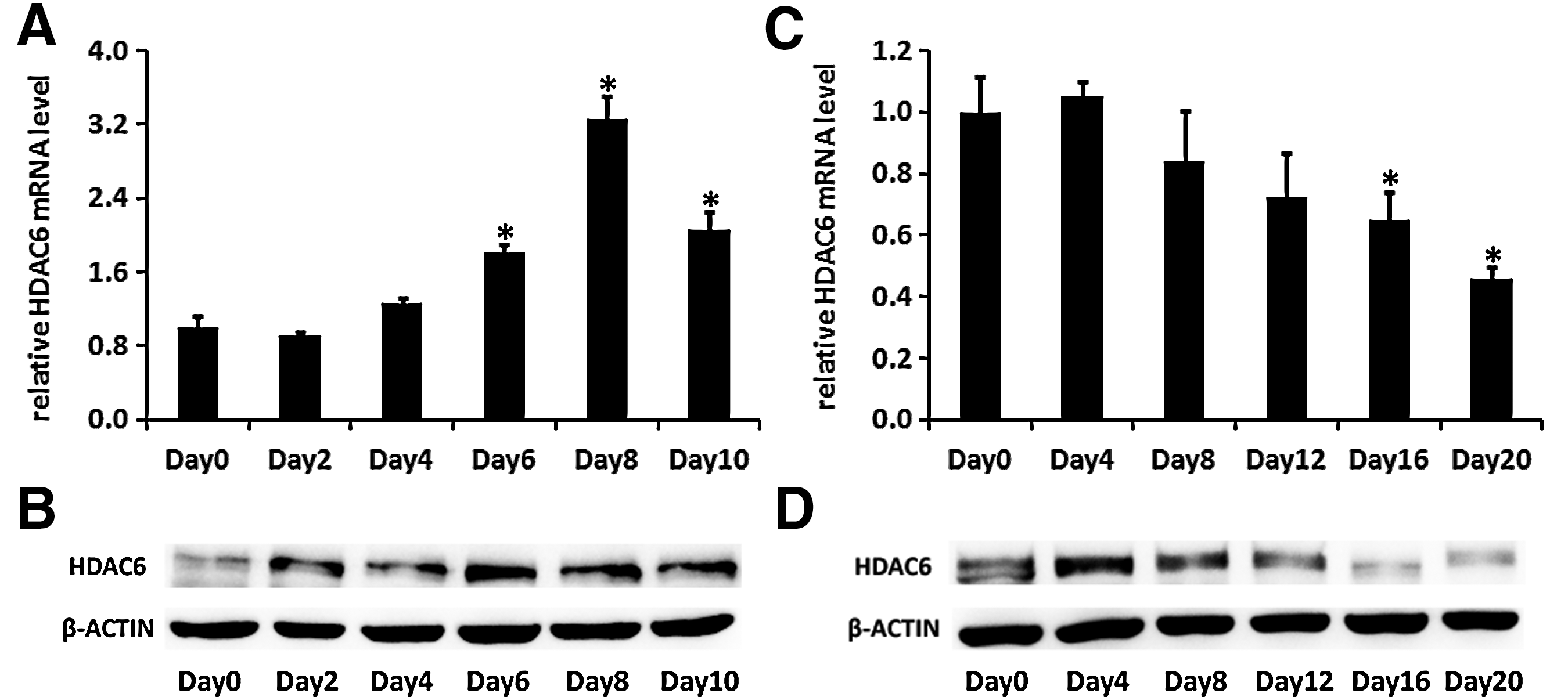
The expression of HDAC6 increases during adipogenic differentiation and decreases during osteogenic differentiation of hADMSCs.
Effect of HADC6 inhibition on adipogenic and osteogenic differentiation of hADMSCs
To investigate the functional effect of HDAC6 on adipogenic and osteogenic differentiation of hADMSCs, we suppressed the expression of HDAC6 by transfecting siRNA of HDAC6 into hADMSCs. As shown in Fig. 5A and B, both mRNA and protein expression levels of HDAC6 were downregulated due to transfection of HDAC6 siRNA. hADMSCs were then induced into adipogenic and osteogenic differentiation, respectively, after transfection with siHDAC6. Oil red O staining (Fig. 5C) and a real-time PCR analysis of adipo-specific markers (Fig. 5D) showed that adipogenic differentiation of hADMSCs was repressed as a result of HDAC6 inhibition. Conversely, the decrease of HDAC6 expression resulted in a significant increase in the ALP activity (Fig. 5E), matrix mineralization deposition (Fig. 5F), and mRNA expression of osteo-specific markers (Fig. 5G). Consistent with the role of miR-22 in the differentiation of hADMSCs, down regulation of HDAC6 inhibited adipogenic differentiation and promoted osteogenic differentiation, indicating that HDAC6 was a molecular target of miR-22 in the regulation of the balance between adipogenic and osteogenic differentiation of hADMSCs.

HDAC6 inhibition by RNA interference suppresses adipogenic differentiation and promotes osteogenic differentiation of hADMSCs.
Discussion
MSCs have the capacity to differentiate into adipocytes and osteoblasts. In general, mechanisms that promote one cell fate actively suppress mechanisms that induce the alternative lineage [5]. In vivo, adipogenesis and osteogenesis of MSCs are competitively balanced. Once the homeostasis is disturbed, pathological disorders such as osteoporosis will occur. Thus, identification and thorough characterization of signaling pathways that influence or determine such balance are of great importance for us to understand the underlying mechanisms of disease initiation and progression. Despite intensive research efforts focusing on the individual differentiation pathways, little is known about the cross-talk between adipogenesis and osteogenesis. Recently, miRNA has emerged as an important regulator of gene expression in a variety of cellular processes. Studies reporting that miRNAs function as mediators of the balance between adipogenesis and osteogenesis of MSCs began to accumulate. miR-204, for instance, has been identified as a regulator between adipogenesis and osteogenesis of MSCs because miR-204 could promote adipogenesis but inhibits osteogenesis through Runx2 suppression [19]. Another study revealed that miR-637 was indispensable to maintain the balance between adipogenic and osteogenic differentiation by repressing adipocyte formation while promoting osteoblast formation through OSX, a critical regulator of bone formation [20].
Here, we found that miR-22 was decreased in adipocyte differentiation but increased in osteoblast differentiation of hADMSCs. Overexpression of miR-22 in hADMSCs enhanced osteoblast formation and suppressed adipocyte formation, indicating the role of miR-22 in regulating the balance between adipogenesis and osteogenesis of hADMSCs.
We then searched for downstream target genes of miR-22 through target prediction databases and found a 7-nt match site complementary to miR-22 in the 3′UTR of HDAC6 mRNA. Dual luciferase reporter assay identified HDAC6 as a direct target of miR-22. Transfection of miR-22 mimics into hADMSCs resulted in upregulation of HDAC6 protein expression without changing the mRNA expression of HDAC6, confirming that miR-22 suppressed HDAC6 expression at a post-transcription level.
HDACs could regulate differentiation through the modulating chromatin structure and suppressing the activity of transcription factors [24]. HDACs inhibition by various HDAC inhibitors could accelerate osteoblast maturation through partially increasing Runx2 activity [26]. HDACs have been reported to affect osteogenic differentiation in various aspects. For example, HDAC4 was found to physically interact with the Runt domain of Runx2, interfering with DNA binding by Runx2, and consequently suppressed activation of Runx2 target genes [27]. Also, HDAC4 could downregulate Runx2 expression and activity in the suppression of osteoblast differentiation by EGFR signaling [28]. In Kang's study, HDAC4 or HDAC5 acted as a co-repressor for TGF-β/Smad3-mediated transcriptional inhibition of Runx2 function in osteoblast differentiation [29]. Additionally, Jeon et al. showed that HDAC4 and HDAC5 could deacetylate Runx2 and promote Smurf-mediated degradation of Runx2 [30]. HDAC5 was also identified as a target of miR-2861, and was found to regulate osteoblast differentiation by enhancing Runx2 degradation [31]. HDAC3 was found to interact with Runx2 to repress the osteocalcin promoters and regulate osteoblast differentiation without effecting Runx2 expression levels [32]. Besides, HDAC3 conditional knockout mice exhibited lower bone density but more bone marrow adipocytes [33].
HDAC6, a member of class II of the HDAC family, has been found to play an important role in various biological processes, such as cell migration, cell–cell interactions, tumor growth, and development events [34 –36]. According to Westendorf's study, HDAC6 could interact with the carboxy terminus of Runx2 and act as a corepressor of Runx2 to inhibit the p21 promoter in mouse preosteoblasts MC-3T3 cells [21]. In our research, we found that the expression of HDAC6 was upregulated during adipocyte differentiation but downregulated during osteoblast differentiation. Moreover, silencing of HDAC6 by siRNAs could remarkably inhibit adipogenic differentiation and promote osteogenic differentiation, consistent with the effect of overexpression of miR-22 in hADMSCs, indicating HDAC6 as a promoter in adipogenesis while an inhibitor in osteogenesis.
Besides, we found that both overexpression of miR-22 and inhibition of HDAC6 promoted osteoblast formation without affecting Runx2 expression levels (data not shown). These findings provided evidence that HDAC6 might regulate osteogenic differentiation by directly interacting with the Runx2 protein to suppress osteogenic-related genes, which could be considered as one of the mechanisms underlying miR-22-mediated regulation of osteogenic differentiation. Since Runx2 was identified as an inhibitor in adipocyte formation [25] and a modulator of the balance between adipogenesis and osteogenesis [19], suppressed Runx2 activity by HDAC6 interaction might be also partially responsible for the positive regulation in adipogenesis. Hence, it is possible that one mechanism of HDAC6 regulation in the balance of adipogenesis and osteogenesis in hADMSCs is partially mediated by Runx2.
In addition, Lee's study suggested that HDAC6 was required for growth factor-induced activation of the PI3K/AKT-signaling cascade [35]. The PI3K/AKT-signaling pathway was reported to promote adipogenic differentiation by increasing the expression and transcriptional activity of PPARγ [37,38]. PPARγ was also considered as a potential regulator between adipogenesis and osteogenesis, since activation of PPARγ could inhibit Runx2 in mouse MC-3T3 cells [39], and its sufficiency could increase bone mass by promoting osteoblast formation from bone marrow progenitors [40]. Due to above studies, HDAC6 might also regulate adipogenic and osteogenic differentiation of hADMSCs partially through PI3K/AKT-mediated PPARγ regulation.
Moreover, distinguished from other members in class II of the HDAC family, HDAC6 contains 2 catalytic domains with the deacetylase activity and a specific ubiquitin-binding motif, the BUZ finger, at its C-terminus and plays roles in both deacetylse-related and ubiquitin-related regulations [41]. Therefore, the effect of HDAC6-mediated ubiquitination or deacetylation on regulatory factors involved in hADMSCs differentiation might also be considered as a possible mechanism for HDAC6-modulated adipo/osteogenic differentiation in hADMSCs. However, further studies still need to be performed to reveal the precise molecular mechanisms underlying HDAC6 regulation of the balance between adipogenic and osteogenic differentiation.
In this study, we showed that transfection of miR-22 mimics could inhibit HDAC6 protein levels without affecting its mRNA levels in hADMSCs (Fig. 3C, D), but we also found that mRNA expression of HDAC6 was changed during osteogenic and adipogenic differentiation of hADMSCs (Fig. 4A, C), indicating that besides the post-transcriptional inhibition by miR-22, there might be other upstream molecules contributing to the regulation of HDAC6 expression at the mRNA/transcription level directly or indirectly in hADMSCs. A few studies have reported the regulation of HDAC6 expression at the mRNA level in cancers so far. According to the latest study by Hsieh et al., the upstream region of the HDAC6 promoter contained a consensus CREB1 response element and could be transactivated by cAMP/PKA/CREB1-signaling cascade in breast cancer [42]. Another newly published research by Birdsey et al. revealed that the erythroblast transformation specific (ETS) transcription factor (Erg) drives HDAC6 transcription directly in endothelial cells and angiogenesis, and the ETS binding sites were identified in the HDAC6 promoter [43]. In addition to these recent studies, the oncogenic Ras and HER2 were reported, respectively, to increase HDAC6 mRNA expression in tumorigenesis [35,44]. Besides, knockout of TDP-43, the transactive response DNA-binding protein, could significantly reduce the mRNA level of HDAC6 in HEK293E cells and neuronal SH-SY5Y cells and Drosophila melanogaster [45]. However, few studies have been reported on the upstream regulation of HDAC6 mRNA expression in stem-cell differentiation. The regulatory mechanisms mentioned above might have impacts on HDAC6 expression in similar pathways during the differentiation of hADMSCs, but the potential molecular mechanisms still need to be investigated in future studies.
In conclusion, we identified miR-22 and its downstream target HDAC6 as important regulators of the balance between adipogenic and osteogenic differentiation of hADMSCs. In our study, overexpression of miR-22 significantly inhibited adipocyte formation but promoted osteoblast differentiation by directly suppressing HDAC6. These results indicated that miR-22 and HDAC6 might not only be useful in illustrating the pathology of bone-related diseases but also could be developed as potential targets in the therapy of those diseases.
Footnotes
Acknowledgments
This study was supported by grants from the “863 Projects” of Ministry of Science and Technology of the People's Republic of China (No. 2011AA020100), National Natural Science Foundation of China (No. 30700321), the National Key Scientific Program of China (No. 2011CB964901), and Program for Cheung Kong Scholars and Innovative Research Team in University-PCSIRT (No. IRT0909).
Author Disclosure Statement
All authors have no conflict of interest.