
Abstract
Recent discovery of a method for derivation and culture of germline-competent rat pluripotent stem cells (PSCs) enables generation of transgenic rats or knock-out rats via genetic modification of such PSCs. This opens the way to use rats, as is routine in mice, for analyses of gene functions or physiological features. In mouse or human, one widely used technique to express a gene of interest stably and ubiquitously is to insert that gene into the Rosa26 locus via gene targeting of PSCs. Rosa26 knock-in mice conditionally expressing a reporter or a toxin gene have contributed to tracing or ablation of specific cell lineages. We successfully identified a rat orthologue of the mouse Rosa26 locus. Insertion of tdTomato, a variant of red fluorescent protein, into the Rosa26 locus of PSCs of various rat strains allows ubiquitous expression of tdTomato. Through germline transmission of one Rosa26-tdTomato knock-in embryonic stem cell line, we also obtained tdTomato knock-in rats. These expressed tdTomato ubiquitously throughout their bodies, which indicates that the rat Rosa26 locus conserves functions of its orthologues in mouse and human. The new tools described here (targeting vectors, knock-in PSCs, and rats) should be useful for a variety of research using rats.
Introduction
U
To generate a genetically modified animal with stable and ubiquitous expression of a gene of interest is essential for current research. One widely used method is to insert that gene into the Rosa26 locus on mouse chromosome 6, identified by random retroviral gene-trap screening using mouse embryonic stem cells (ESCs) [8]. Rosa26 is ubiquitously expressed in embryonic as well as adult tissue, and gene targeting at this locus in ES cells is highly efficient. Insertion of a gene of interest or a loxP-flanked stop codon with a reporter or a toxin gene into Rosa26 thus has been widely used to trace specific cell lineages or, by mating with mice expressing Cre recombinase under the control of specific promoters, to ablate specific cell lineages [9,10].
As in the mouse, so in man: A human ROSA26 locus has also been identified by homology search with mouse Rosa26 sequences [11]. Insertion of sequences encoding a red fluorescent protein (RFP) into the Rosa26 locus of human ESCs allows ubiquitous expression of RFP in both undifferentiated and differentiated states [11]. Features of the Rosa26 locus may be conserved in a variety of species.
Here, as a third model, we report identification of a Rosa26 locus in the rat. Furthermore, we generated rat PSCs expressing tdTomato, a variant of RFP, by gene targeting into the Rosa26 locus, and generated a knock-in rat line via germline transmission of such PSCs.
Materials and Methods
Animals
C57BL/6NCrSlc, BDF1, and ICR mice and Wistar and DA rats were purchased from SLC Japan (Shizuoka, Japan). All experiments were performed in accordance with the animal care and use committee guidelines of the Institute of Medical Science, the University of Tokyo, and of the National Institute for Physiological Sciences.
Culture of PSCs
In this study, 3 ESC lines and 2 induced PSC (iPSC) lines were used (see Table 1). Their culture conditions were as described [12]. In brief, undifferentiated rat PSCs were maintained on mitomycin-C - treated mouse embryonic fibroblasts in an N2B27 medium [1] containing 1 μM PD0325901 (Axon, Groeningen, The Netherlands), 3 μM CHIR99021 (Axon), and 1,000 U/mL of rat leukemia inhibitory factor (Millipore, Bedford, MA).
Judged by PCR.
ESC, embryonic stem cell; iPSC, induced pluripotent stem cell; PCR, polymerase chain reaction.
DA rat-derived ESC (DA3i-1) and iPSC (DAT3-1) lines were newly established. DA3i-1 ESCs were derived from DA rat blastocysts as described [13]. DAT3-1 iPSC were generated from DA rat-derived tail-tip fibroblasts by introducing 3 mouse factors (Oct3/4, Klf4, and Sox2) in one retroviral vector (data not shown).
Construction of vectors and gene targeting
Homology arms were amplified from genomic DNA of DA strain rats by polymerase chain reaction (PCR) using PrimeSTAR or PrimeSTAR GXL DNA polymerase (Takara Bio, Otsu, Japan), according to the manufacturer's protocol. These arms, with an additional NheI site and an MC1-promoter driven DTA cassette amplified from an MC1-DTA vector, a kind gift from Dr. T. Yagi (Osaka University, Osaka, Japan), were inserted into pBluescript KS(+) (Stratagene, La Jolla, CA) with an infusion cloning kit (Takara Bio) (prRosa26-1 in Fig. 1A). A splice acceptor sequence amplified from a pSAβ-geo vector, a kind gift from Dr. P. Soriano (Fred Hutchinson Cancer Research Center, Seattle, WA); tdTomato amplified from ptdTomatoN1 (Clontech, Palo Alto, CA); and IRES-Puror-pA amplified from pCAG-Cre-IRES-Puror-pA, a kind gift from Dr. J. Miyazaki (Osaka University), were inserted into the NheI site of prRosa26-1.

Identification of Rat Rosa26 locus and gene targeting of rat pluripotent stem cells by inserting a cassette containing tdTomato into Rosa26 locus.
Electroporation for gene targeting was carried out as described [5]. In brief, 2.5∼5×106 rat PSCs suspended in PBS were mixed with 20 μg linearized targeting vector digested by the Sall1 restriction enzyme and were transferred to a Gene Pulser cuvette (Bio-Rad, Richmond, CA). Electroporation was carried out at 800 V, 10 μF in Gene Pulser equipment (Bio-Rad). After electroporation, PSCs were seeded onto mitomycin-C - treated puromycin-resistant mouse embryonic fibroblasts made in house, and 24 h later, 1.5 μg/mL puromycin (Invitrogen, Carlsbad, CA) was added to the culture medium.
Reverse transcription–PCR and quantitative real-time PCR analysis
cDNA synthesized using the ThermoScript™ reverse transcription (RT)–PCR System (Invitrogen, Carlsbad, CA) from extracts of BLK2i-1 rat ESCs was used for the RT-PCR analysis of Rosa26 noncoding RNA. PCR primers are shown in Supplementary Fig. S3 (Supplementary Data are available online at
3′ and 5′ rapid amplification of cDNA ends analysis and identification of full-length noncoding RNA
3′ and 5′ rapid amplification of cDNA ends (RACE) was performed using the SMARTer™ RACE cDNA Amplification Kit (Clontech), according to the manufacturer's protocol. Synthesized cDNAs were amplified using an universal primer A mix with 3′ or 5′ RACE primers (primers shown in Supplementary Fig. S3). Amplified cDNAs were cloned into the pCR-Blunt IITOPO vector (Invitrogen), and their sequences were confirmed.
Genotyping and Southern blotting
DNA was extracted using QIAamp DNA Mini Kits (Qiagen, Germantown, MD) from picked-up PSCs. For genotyping, PCR primers for amplification of the Rosa26 knock-in locus were Fw, 5′-CAGAAAAGGCGGAGCGAGCCCAAG-3′, and Rv, 5′- GGGCCCTCACATTGCCAAAAGACGG-3′. For the Southern blot analysis, genomic DNA extracted from PSCs was digested by the ApaI restriction enzyme and hybridized with a DNA probe cloned from the upstream region of the rat Rosa26 5′-arm.
Embryo manipulation
Rat and mouse embryos were prepared using published protocols [12,14]. In brief, rat blastocysts were collected in the HER medium [15] containing 18% fetal bovine serum (Invitrogen) from the oviduct and the uterus of rats 4.5 days post coitum (dpc). These embryos were transferred into the mR1ECM medium [16] containing 80 mM NaCl and 0.01% polyvinyl alcohol (Sigma-Aldrich Co., St. Louis, MO) and were cultured for about 1 h until injection. Mouse 8-cell/morula stage embryos were collected in the M2 medium (Millipore) from the oviduct and the uterus of BDF1×C57BL/6 mice 2.5 dpc. These embryos were transferred into the KSOM-AA medium (Millipore) and were cultured for 24 h before blastocyst injection.
For micromanipulation, PSCs were trypsinized and suspended in the PSC culture medium. A piezo-driven micromanipulator (Prime Tech, Tokyo, Japan) was used to drill zona pellucida and trophectoderm under the microscope, and 10 PSCs were introduced into blastocyst cavities near the inner cell mass. After blastocyst injection, embryos underwent follow-up culture for 1–2 h. Rat blastocysts were transferred into the uteri of pseudopregnant recipient Wistar rats (3.5 dpc), and mouse blastocysts were transferred into the uteri of pseudopregnant recipient ICR mice (2.5 dpc).
Flow cytometry analysis
To analyze chimerism of chimeric rats at the adult stage, we used peripheral blood cells obtained from the retro-orbital venous plexus. Leukocytes isolated by osmotic lysis of erythrocytes were stained with APC-conjugated mouse anti-rat CD45 antibody (BD Biosciences, San Diego, CA).
To analyze chimerism of interspecific chimeras at the fetal stage, embryonic fibroblasts were stained with biotin-conjugated mouse anti-rat CD54 antibody (ICAM-1, 1A29), Alexa Fluor 647-conjugated goat anti-mouse IgG antibody, and FITC-conjugated rat anti-mouse CD54 antibody (all BD Biosciences).
All stained cells were analyzed by FACSCanto II (BD Bioscience).
Result and Discussion
Given that the transgenic rat carrying a mouse Rosa26 promoter-driven EGFP construct revealed ubiquitous expression of EGFP [17], we inferred that the promoter region of the Rosa26 locus contained a highly conserved sequence. To identify a Rosa26 locus in rat genome, we searched for similar sequences in the UCSC Genome Browser database (
To see if this region, like the Rosa26/ROSA26 locus in mouse or human, actually allowed ubiquitous expression of an inserted gene, we cloned homology arms from genomic DNA of the DA rat and constructed a targeting vector to insert a splice acceptor with the tdTomato-IRES-Puror-pA sequence (Fig. 1C). We transduced the linearized targeting vector into 3 ESC lines and 2 iPSC lines derived from various strains and tissues. Although only small numbers of colonies were observed after electroporation and drug selection, on average about 30% of picked-up clones demonstrated correct targeting judged by PCR using genomic DNA (Table 1). Gene targeting was also confirmed by Southern blotting (Fig. 1D). As expected, all positive clones ubiquitously expressed tdTomato (Fig. 1E). Survival rates of PSCs after drug selection were low compared with mouse PSCs subjected to conventional targeting. This might result from low transduction efficiency of the targeting vector or from high sensitivity to antibiotics in rat PSCs [7].
We injected rat ESCs from one of the Rosa26-tdTomato knock-in lines, BLK-RT2, into rat blastocysts. Rat ESCs of the original cell line, BLK2i-1, are germline-competent. The line was established from blastocysts of rats with black coats derived from F1 crosses of Wistar (white-coated) and DA (black-coated) rats back-crossed with Wistar rats for at least 3 generations [18]. A total of 116 injected blastocysts were transferred into uteri of pseudo-pregnant rats and at weaning 27 out of 36 pups proved chimeras. Neonatal chimeric rats expressed tdTomato throughout their bodies (Fig. 2A). They grew into adults normally and showed coat color chimerism (black, donor ESC-derived; white, host embryo Wistar strain-derived) (Fig. 2B). To estimate their chimerism, we analyzed tdTomato expression in peripheral blood of male chimeric rats by flow cytometry. Although chimerism varied individually, most chimeras showed over 50% chimerism in CD45-positive peripheral blood mononuclear cells (Fig. 2C).
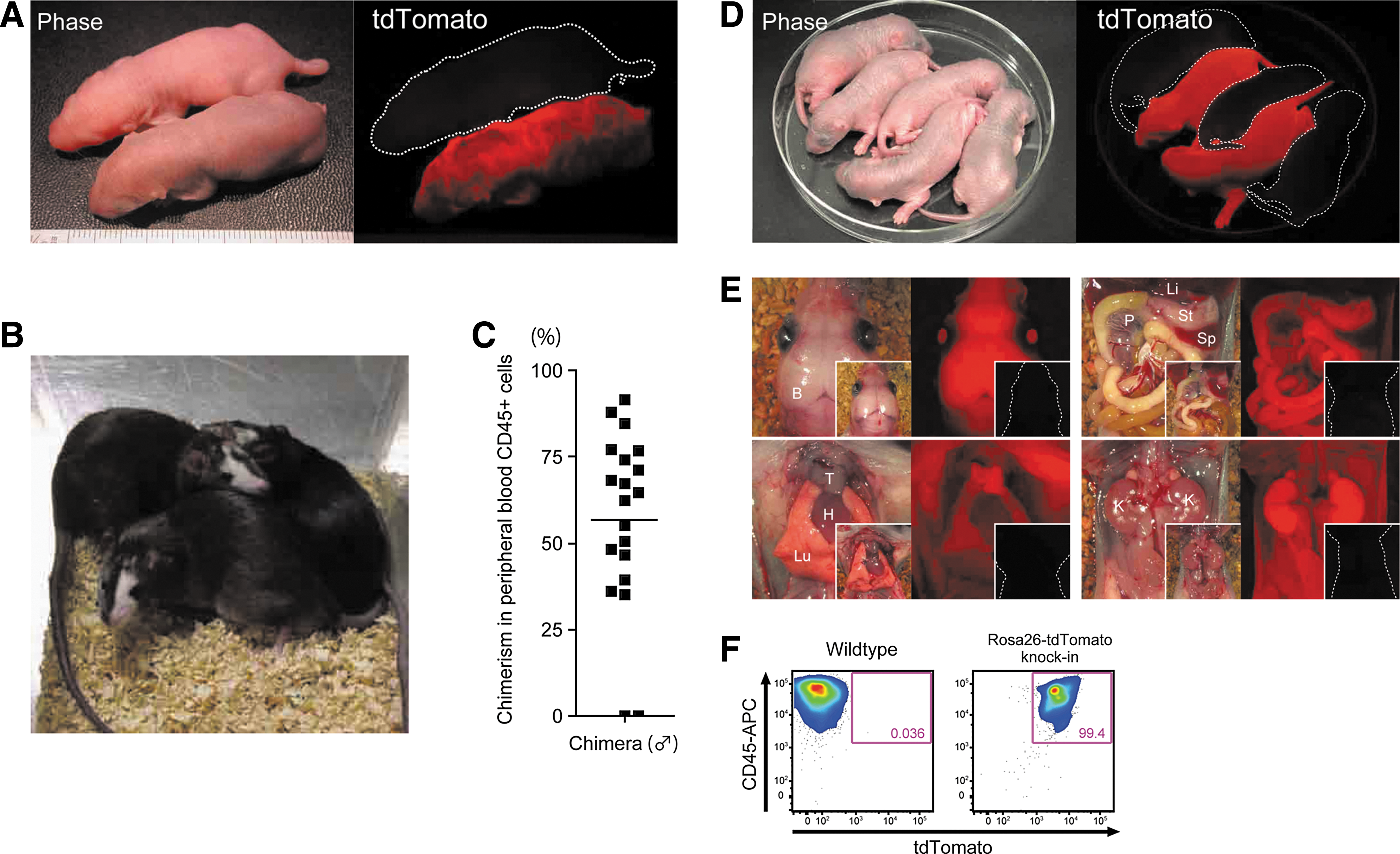
Generation of chimeric rats by injection of Rosa26-tdTomato knock-in rat ESCs into rat blastocysts.
Next, we mated 3 male rats exhibiting relatively higher chimerism with wild-type females to see if BLK-RT2 rat ESCs were capable of germline transmission. Offspring of 2 chimeras expressed tdTomato ubiquitously throughout their bodies, indicating successful germline transmission (Fig. 2D). Expression of tdTomato was detected in all main organs at the neonatal stage (Fig. 2E) and in CD45-positive blood cells at the adult stage (Fig. 2F). These data provided strong evidence for functional conservation of the Rosa26/ROSA26 locus among rat, mouse, and human.
We also injected BLK-RT2 rat ESCs into mouse blastocysts. A total of 32 injected blastocysts were transferred into uteri of pseudo-pregnant mice, and 13 out of 16 fetuses proved interspecific chimeras (Fig. 3A). To distinguish host- and donor-derived cells, we cultured embryonic fibroblasts from these chimeras and stained them with a specific antibody against mouse or rat CD54. Only cells expressing rat CD54 also expressed tdTomato (Fig. 3B). Therefore, Rosa26 knock-in rat PSCs can also be used for interspecific blastocyst complementation to make rat organs in mice [12].

Generation of interspecific chimeras by injection of Rosa26-tdTomato knock-in rat ESCs into mouse blastocysts.
Via gene targeting of PSCs, we have successfully generated Rosa26 knock-in rats that express tdTomato ubiquitously. These PSCs and rats will provide useful tools for cellular or organ transplantation experiments, and the targeting vector constructed here will be useful for expressing genes of interest stably and ubiquitously in a variety of rat PSCs. These powerful tools should contribute to progress in a variety of research using rats.
Footnotes
Acknowledgment
We thank Dr. A. Knisely for critical reading of the article. This work was supported by grants from the Japan Science and Technology Agency (JST), KAKENHI (23700507) Grant-in-Aid for Young Scientists (B) from Japan Society for the Promotion of Science (JSPS), and the Ministry of Education, Culture, Sport, Science, and Technology (MEXT).
Author Disclosure Statement
Hiromitsu Nakauchi is a founder and shareholder of ReproCELL, Inc. There is no conflict of interest to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.