
Abstract
Androgen-independent prostate cancer (AIPC) is difficult to treat. Present study is to explore the inhibitory effect of a cytokine environment on AIPC and its mechanism. We utilized nerve growth factor (NGF)/γ-interferon (γ-IFN) to change the cytokine environment. Animal models and 2 androgen receptor (AR)-negative prostate cancer cell lines were used to evaluate the effect of NGF/γ-IFN. Flow cytometry, immunocytochemistry, western blotting, Tunel assay, colony formation efficiency, gene microarray, and in vivo bioluminescence were used to discern the mechanisms within NGF/γ-IFN that effect the environment. In vitro, NGF/γ-IFN effectively inhibited the proliferation of AIPC cell lines and promoted the apoptosis of the cancer cells. In vivo, NGF/γ-IFN suppressed the growth and metastasis of a tumor mass that arose from the AIPC cell line. After NGF/γ-IFN treatment, the AR-negative cell lines re-expressed AR and were then able to respond to the androgen. Contrary to expectations, the proliferation of cells was inhibited after dihydrotestosterone was added, and the results indicated that NGF/γ-IFN decreased the proportion of cancer stem cells. NGF/γ-IFN worked mainly through the downregulation of fibroblast growth factor receptor 2.
Introduction
P
ADT can stimulate the formation of androgen-independent prostate cancer (AIPC). Many studies have confirmed that the decline of testosterone disturbs the level of certain cytokines that may influence the growth of tumors by regulating mitotic division and differentiation. A rebalance of the cytokine level may be a new therapy for AIPC. Through screening, nerve growth factor (NGF) and γ-IFN were found to inhibit prostate cancer cell lines, Du145 and PC-3M.
NGF and γ-IFN are notably influenced by testosterone, and the decline of testosterone decreased the levels of NGF and γ-IFN. In this study, our results indicated that NGF and γ-IFN inhibited AIPC cell lines Du145 and PC-3M in vivo and in vitro. By the dose–effect curve, 50 ng/mL of NGF and 1,000 IU of γ-IFN will present the optimal effect.
With NGF/γ-IFN treatment, previously negative androgen receptor (AR) was re-expressed in Du145 and PC-3M. The re-expressed AR can be activated by dihydrotestosterone (DHT). The proportion of the integrineα2β1+CD44+ cells decreased in the 2 cell lines. In previous reports, integrineα2β1+CD44+ cells proved to be prostate cancer stem cells (CSCs) in prostate cancer tissue and cell lines. Thus, the formation of AIPC may be due to the selective growth of prostate CSCs, which do not express an AR. CSCs are a small population of cells in a tumor that have the ability to self-renew and infinitely proliferate. Non-CSCs represent the remainder of the cells in a tumor and possess limited proliferation potential.
CSCs resist radiotherapy [6], chemotherapy and hormone therapy due to their unique characteristics [7 –11]. Conventional therapy could eliminate most of the non-CSCs in tumors, whereas CSCs survive and become the roots of recurrence and metastasis. Even so, CSCs need stem cell niches to regulate self-renewal and differentiation. A stem cell niche is defined as the microenvironment surrounding the stem cells.
Cytokines form a special microenvironment that strongly influences the proliferation and differentiation of stem cells [12]. NGF is expressed in prostate tissues and carcinoma [13,14], and promotes the differentiation of various tumors [15 –17]. Importantly, NGF is closely correlated with androgen levels [18,19]. One study showed that Du145 cells treated with NGF re-expressed AR, decreasing cell invasion [20]. The level of γ-IFN is also regulated by ADT [21], and it has been shown that γ-IFN stimulates the differentiation of prostate cancer [22]. In the nervous system, γ-IFN upregulates the trkA receptor (one of the NGF receptors) in neuroblastoma cells and coordinates with NGF to induce the differentiation of these cells [23].
In the present study, NGF and γ-IFN were used either solely or jointly to treat the AIPC cell lines, Du145 and PC-3M. The combination of NGF and γ-IFN (NGF/γ-IFN) effectively inhibited tumor proliferation in vivo and in vitro, and functional AR was re-expressed in the cell lines. Contrary to expectations, AR induced by NGF/γ-IFN had entirely different functions. This activation re-expressed AR and suppressed the proliferation of NGF/γ-IFN-treated cells. AR activation had the promoting proliferation effect in common.
After the NGF/γ-IFN treatment, a decline in the proportion of CSCs and lower expression of fibroblast growth factor receptor 2 (FGFR2) was detected in the 2 cell lines. The results suggested that NGF/γ-IFN may play a role through the downregulation of CSCs and FGFR2. This study provides a new insight into the treatment of AIPC and the role of AR in various cytokine environments and in differentiated states of cells.
Materials and Methods
Cell lines and dose–effect experiment
The human AIPC cell lines, Du145 and PC-3M, were purchased from the Chinese Academy of Sciences (originally obtained from ATCC). All cells were cultured in the DMEM (Hyclone) supplemented with 10% of fetal bovine serum (Hyclone) at 37°C in a 5% CO2 atmosphere. Once they reached 80% confluence, the cells were detached with ACCUTASE (Millipore), seeded into a 96-well plate, and cultured in the DMEM supplemented with 10% fetal bovine serum at 37°C with 5% CO2. After 24 h, cells were grouped, and a combination of NGF and γ-INF was added. According to the previous study (20), the concentration of NGF was set to 50 ng/mL. To explore the optimum concentration of γ-INF, 6 concentration gradients were set. Cells that added 50 ng/mL of NGF served as the control. Other than 50 ng/mL of NGF, other gradients, respectively, added were 200 IU, 500 IU, 1,000 IU, 2,000 IU, and 4,000 IU of γ-INF. Following 7 days of treatment, the cell number was measured using CellTiter-BlueTM reagent (Promega), according to the manufacture's instruction. Briefly, cells were rinsed with PBS, and 100 μL of Keratinocyte SFM was added into each well. Then, 20 μL of CellTiter blue reagent was added. The cells were incubated for 4 h in the dark at 37°C for 6 h. The intensity of fluorescence in each well was detected using a fluorometer (Infinite 200), and the cell number was calculated according to the standard curve.
Cytokines treatment
According to the results of the dose–effect experiment, the human AIPC cell lines, Du145 and PC-3M, were plated into 25-cm2 flasks at a concentration of 0.5×105 cells/mL, followed by the addition of 50 ng/mL of NGF (Promega), 1,000 U/mL of γ-INF (Peprotech, Inc.) or each 24 h later. The medium and the cytokines were renewed every 2 days, and cells were harvested after 7 days of treatment.
Proliferation assay
The Du145 and PC-3M cells (150 cells/200 μL) were seeded into a 96-well plate and cultured in the DMEM supplemented with 10% of fetal bovine serum at 37°C with 5% CO2. After 24 h, cells were grouped and either 50 ng/mL of NGF, or 1,000 U/mL of γ-INF, or both were added, while untreated cells served as the control. Following 7 days of treatment, the cell number was measured with the above method.
Immunocytochemistry and in situ cell death detection
Cells (200 cells/well) were seeded onto 8-mm coverslips and placed in a 24-well plate. A previous cytokine environment was used to culture the cells for 7 days, and then cells were rinsed with PBS and fixed with 4% of paraformaldehyde. The slides were rinsed again and treated with 1× permeabilization buffer (eBiosience). Next, they were washed and labeled with proliferating cell nuclear antigen (PCNA), CK5, CK18, or CK19 monoclonal antibodies (Abcam) for 2 h at 37°C, followed by incubation with a PE-labeled secondary antibody. The slides were then incubated with DAPI (300 nM; Wako pure chemical industries, LTD.) for 2 min and washed. We used the in situ cell death detection kit fluorescein (Roche Diagnostic) to detect cell apoptosis, according to the manufacturer's protocol. An antifade solution (Millipore) was used to seal the slides, and they were examined using a fluorescent microscope.
Test of androgen sensitivity
The cytokine-pretreated cells (24 wells/group) were seeded into a 96-well-plate at a density of 500 cells per well. To exclude the interference of other cytokines and hormones, cells were cultured in 200 μL of a keratinocyte serum-free medium without bovine pituitary extract, epidermal growth factor (EGF), leukemia inhibitory factor, NGF, or γ-INF.
In each group, 8 wells of cells were incubated with 20 nM DHT (Sigma), and another 8 wells were added with vehicle. The remaining 8 wells contained 20 nM DHT and 20 nM of flutamide, and on the seventh day, the cell number was measured using Cell Titer Blue™ reagent.
Flow cytometry
The treated cells were detached using ACCUTASE and washed with PBS, then re-suspended, and incubated in the dark with 1:200 of anti-CD44 antibody-PE (Novus Biologicals, LLC.) and 1:200 of anti-integrin α2β1 antibody-FITC (Life Span Bioscience) for 10 min. For NGF/γ-INF-treated cells, 1:500 of anti-AR antibody (Abcam) and 1:500 of PE-labeled secondary antibody were then added. The cells were incubated and rinsed for 3 times, followed by the addition of 1:200 of anti-integrin α2β1 antibody-FITC (Life Span Bioscience) for 10 min. The labeled cell suspension was centrifuged and washed with PBS in triplicate. The centrifuged cells were re-suspended in 500 μL of PBS and were subjected to flow cytometry (FACSCalibur; Becton Dickinson), and then the percentage of positive events in the gated population was measured.
Assay of colony formation efficiency
The untreated and cytokine-treated cells were plated in a 24-well plate with 200 cells per well in 500 μL of a keratinocyte SFM (keratinocyte serum-free medium containing 25 μg/mL of bovine pituitary extract, 10 ng/mL of EGF, and 2 ng/mL of leukemia inhibitory factor; GIBCO). The medium was renewed every 3 days, and after 30 days, we counted the colonies that contain 32 cells.
Quantitative real-time PCR array
The total RNA was isolated with TRIzol reagents. Quantitative real-time PCR was carried out using Signal Transduction PathwayFinder PCR Array and Cancer PathwayFinder PCR Array (SA Bioscience Corporation) to simultaneously examine the mRNA levels of 168 genes involved in 24 signal transduction pathways. The experiments were performed in triplicate according to the protocol of the manufacturer. The gene expression profiles of the untreated and treated cells were compared, and the data were normalized for either GAPDH or β-actin levels by the ΔΔCt method. Following our reported methods [24], quantitative real-time PCR was used to examine the expression of 2 isoforms of the FGFR gene (FGFR2), FGFR2-IIIb (forward primer: 5′-TGCTGGCTCTGTTCAATGTG-3′; reverse primer: 5′-GGCGATTAAGAAGACCCCTA-3′) and FGFR2-IIIc (forward primer: 5′-ACACCACGGACAAAGAGATT-3′; reverse primer: 5′-GGCGATTAAGAAGACCCCTA-3′).
Western blotting analysis
Cells were lyzed, and western blotting analysis was performed as previously described [20]. Every analyzed sample contained 40 μg of protein. The dilution of anti-AR, anti-FGFR2, anti-telomerase reverse transcriptase (TERT), anti-actin, anti-GAPDF (Abcam), anti-AKT, and anti-p-AKT antibody (Cell Signaling Technology, Inc.) was 1:1,000.
FGFR2 signaling blockage using a neutralizing antibody
Du145 and PC-3M cells were plated into 25-cm2 flasks at a concentration of 0.5×105 cells/mL in the DMEM. The medium was supplemented with 10% of fetal bovine serum and anti-FGFR2-neutralizing antibody (1: 300; Abcam) at 37°C in a 5% CO2 atmosphere. After 7 days, gene expression changes were measured using the PCR Array. Meanwhile, the cell number, AR expression, and proportion of CSCs were measured using a proliferation assay, immunocytochemistry, and flow cytometry.
In vivo tumorigenicity and metastasis in NOD/SCID mice
The PC-3M cells that stably expressed the luciferase gene (PC-3M-luc-C6 cells) were purchased from Xenogen. Cells were resuspended in PBS at a concentration of 1×106 cells/0.2 mL. NOD/SCID mice (n=24) were injected with firefly luciferin (150 mg/kg) by intraperitoneal injection. After 7 to 8 min, mice were anesthetized with 3% isoflurane, and then 1×106 PC-3M-luc-C6 cells were injected subcutaneously into the right dorsal flanks of each mouse. The images of tumor cells were examined using a luminometer (NightOWL IILB 981; Berthold Technologies). After 7 days, the mice were imaged and divided into 4 groups (n=6). The different groups were subcutaneously injected with PBS, NGF (0.1 μg/g body weight), γ-interferon (γ-IFN) (2,000 U/g body weight), or both once a day for 10 days. Afterward, the tumors were imaged every 10 days.
Statistical analysis
Significance was determined by a paired t-test in paired designing experiments and 1-way analysis of variance in multiparameter experiments. A value of P<0.05 was used considered statistically significant.
Results
Optimum concentration of NGF/γ-IFN in inhibiting AIPC cell line
Using the dose–effect experiment, the optimum concentration of NGF/γ-IFN was determined. According to the previous report, the optimum concentration of NGF is 50 ng/mL; therefore, the dose of γ-IFN was adjusted gradually. The results indicated that a combination of 50 ng/mL NGF and 1,000 IU γ-IFN maximized the inhibition of Du145 and PC-3M (Fig. 1A). When the concentration was increased to 50 ng/mL NGF and 4,000 IU γ-IFN, the effect of inhibition was not promoted.

NGF/γ-IFN inhibits the proliferation of AIPC cell lines.
NGF/γ-IFN inhibits the proliferative ability and colony formation of AIPC cell lines, Du145 and PC-3M
After incubation with NGF/γ-IFN for 7 days, the cell counts of Du145 and PC-3M decreased by 62.8%±5.14% and 69.0%±2.03%, respectively (Fig. 1B), and the colony forming efficiency (CFE) decreased by 55.0%±10.35% and 42.6%±6.20%, respectively, compared to the untreated cells (Table 1). The number of proliferating cell nuclear antigen (PCNA)+ cells decreased by 31.6% and 23.0%, respectively (Fig. 1C), and apoptotic cells increased by 46.6% and 37.8% in NGF/γ-IFN-treated Du145 and PC-3M cells (Fig. 2A).
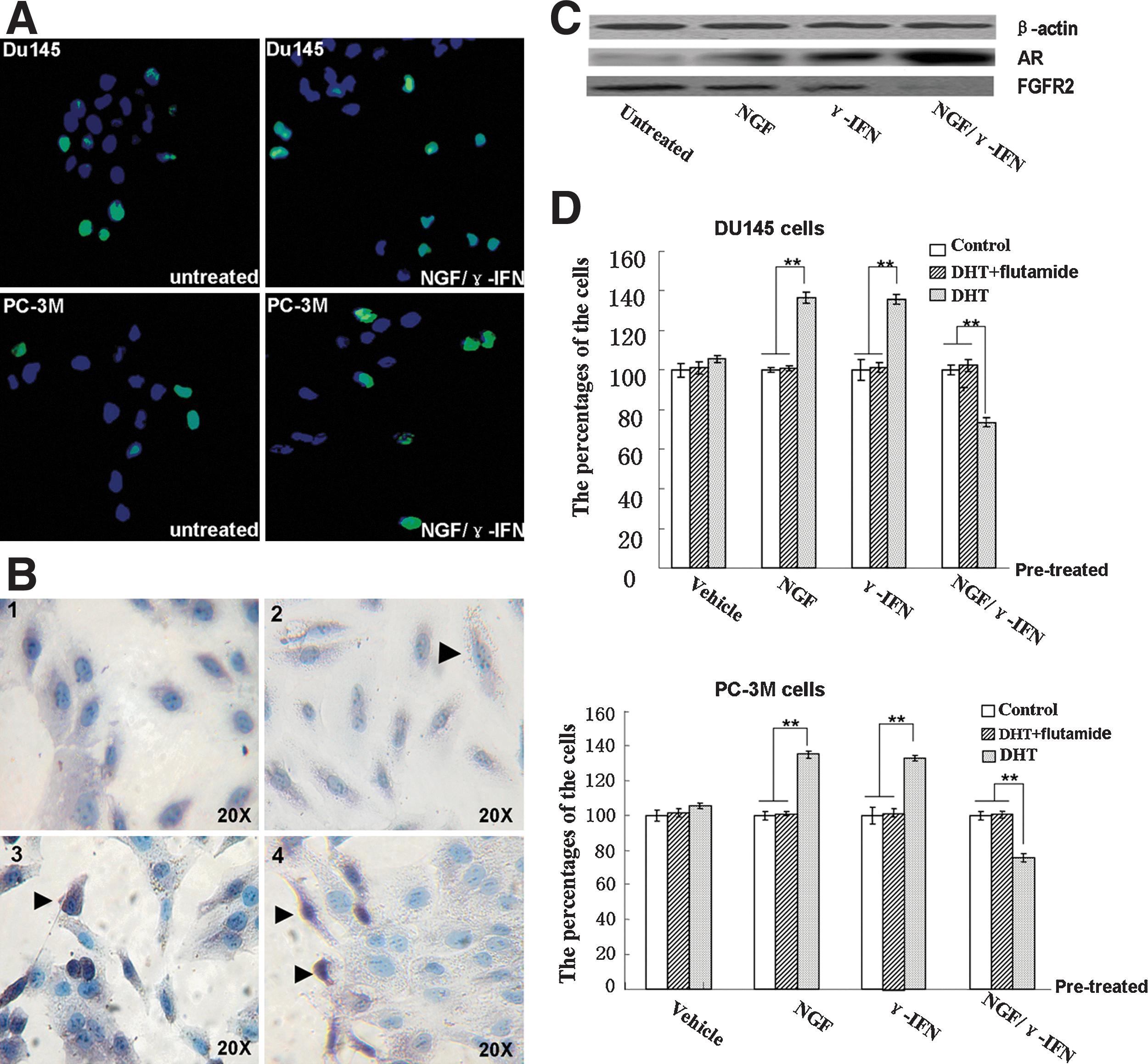
NGF/γ-IFN treatment affected cell apoptosis, AR expression and AR function in AIPC cell lines.
NGF, nerve growth factor; γ-IFN, γ-interferon.
NGF/γ-IFN treatment leads to re-expressed AR
After 7 days of treatment with NGF and γ-IFN, AR expression in Du145 and PC-3M was determined using immunohistochemical staining and western blotting. Less than 2% of untreated Du145 and PC-3M cells expressed AR (Fig. 2B2). After treatment with NGF/γ-IFN, the percentage of AR+ cells remarkably increased to 37.4%±4.6% and 31.2%±5.7% in Du145 (Fig. 2B3) and PC-3M cells, respectively., Treatment with NGF alone led to 14.3%±3.7% and 11.0%±5.6% of AR+ Du145 and PC-3M cells, respectively, whereas γ-IFN treatment caused 21.3%±2.2% of Du145 cells and 15.1%±5.2% PC-3M to express AR. Western blot analysis confirmed the higher expression of AR in NGF/γ-IFN-treated Du145 cells (Fig. 2C).
AR induced by NGF/γ-IFN has anticancer function
Cytokine-treated cells were pretreated with vehicle, DHT, and a combination of DHT and flutamide for 7 days, and then the proliferation rate was detected. The number of vehicle-treated cells was set at 100%.
With the DHT addition, the pretreatment with NGF alone increased the number of Du145 and PC-3M cells by 36.41% and 34.81%, respectively. The number of γ-IFN pretreated Du145 and PC-3M cells also increased by 35.67% and 32.93%, respectively. Surprisingly, the activation of NGF/γ-IFN-induced AR by DHT suppressed proliferation of the tumor cells. Compared with vehicle-treated cells, the number of DHT-treated cells decreased by 26.62% and 24.28% in Du145 and PC-3M cells after 7 days of DHT treatment (Fig. 2D); however, the effect of DHT could be blocked by flutamide (Fig. 2D). Immunohistochemical staining showed that re-expressed AR was located in the cytoplasm and the nucleolus, and after exposure to DHT, the re-expressed AR translocated to the nucleoli (Fig 2. B4).
NGF/γ-IFN decreased the proportion of CSCs
After incubation with NGF/γ-IFN for 7 days, the percentage of CD44+integrinα2β1+ decreased by 43.97%±8.20% (Fig. 3A) and 37.69%±6.62% (Fig. 3B) in Du145 and PC-3M cells. The immunofluorescence results indicated that integrinα2β1+ cells decreased by 34%±2.3% in NGF/γ-IFN-treated Du145 cells (Fig. 3C). CK5 expression is also downregulated in the 2 cell lines (date not shown). Untreated cells can partly form sphere-like colonies, but NGF/γ-IFN-treated cells rarely arrange this figure (Fig. 3D). The FCS results indicated that AR expressed on integrinα2β1- cells (Fig. 4A). As suggested in the results, the most of remaining cells may not be CSCs after NGF/γ-IFN treatment. Of them, parts of cells are more differentiated and can response to androgen.
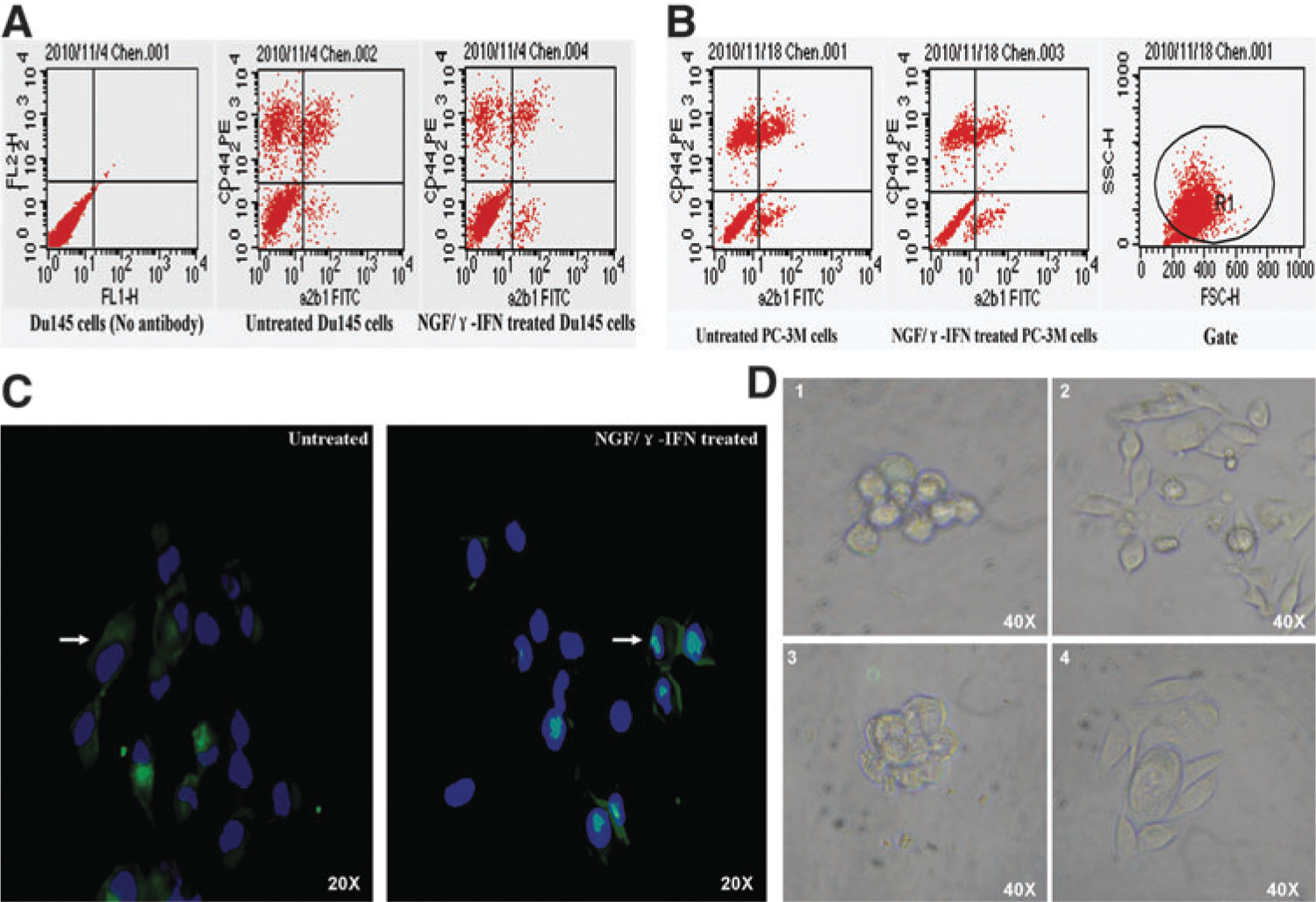
NGF/γ-IFN decreased the proportion of cancer stem cells.
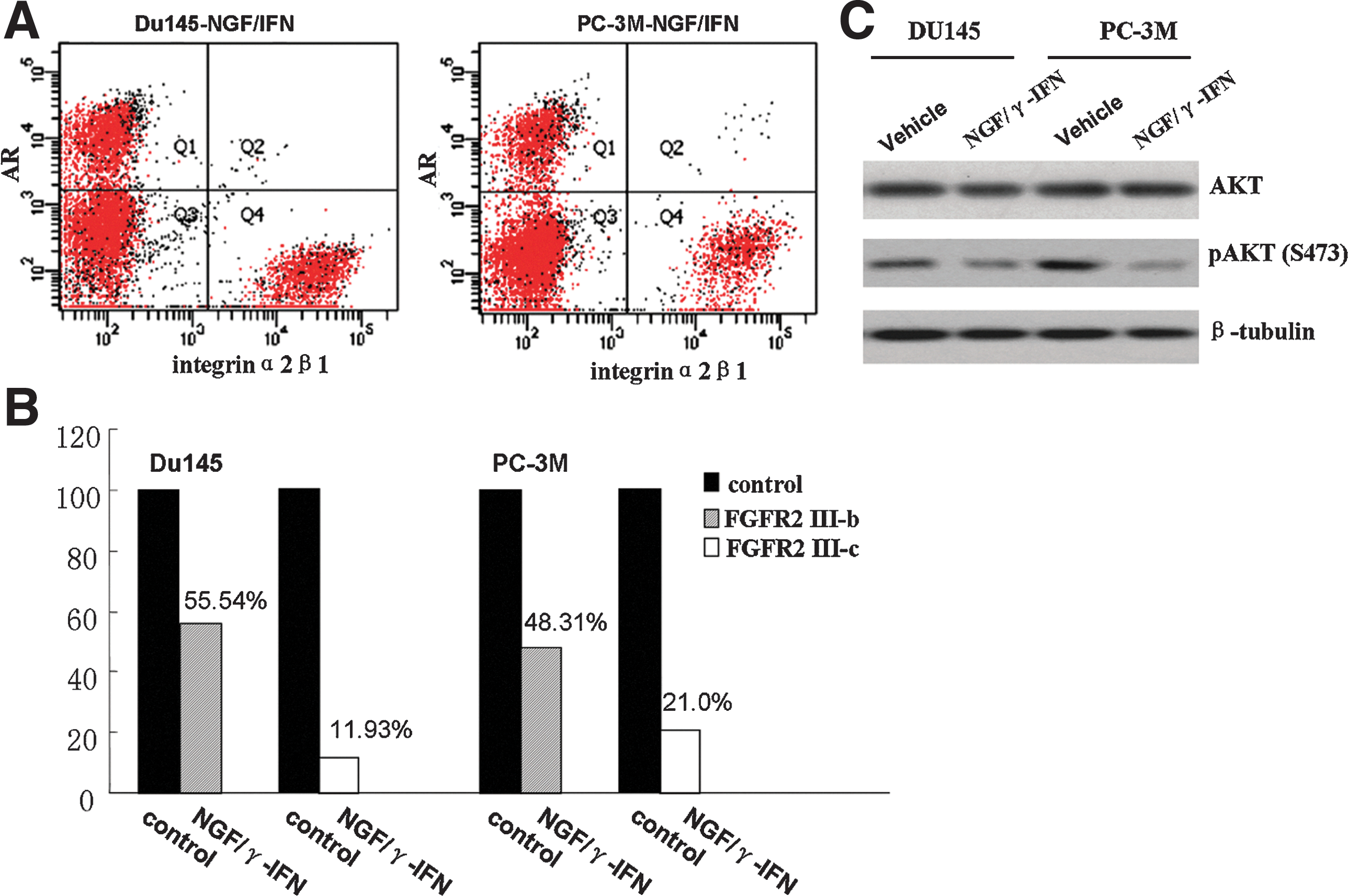
NGF/γ-IFN induced AR expression in Intergrinα2β1- cells. The cytokines mainly decreased the expression of FGFR2 IIIc and resulted in a reduction of phosphorylated AKT at Serine 473.
FGFR2, TERT, and Wnt are reduced by NGF/γ-IFN treatment
The genes with altered expression in response to NGF and/or γ-IFN treatment are listed in Table 2. Among these genes, the downregulation of FGFR2 expression was in agreement with the differentiation of the prostate CSCs. In NGF/γ-IFN-treated cells, FGFR2 was downregulated by 9.42 (Du145) and 7.15 (PC-3M) times compared with untreated cells. Western blot analysis confirmed the decreased expression of FGFR2 in NGF/γ-IFN-treated cells (Fig. 2C). In all of the real-time PCR super-array microarrays, TERT, wingless-type MMTV integration site family member 1 (Wnt1) and Wnt2 were found to be remarkably downregulated in NGF/γ-IFN-treated cells (Table 2).
3.7. Blockade of FGFR2 signaling showed a similar effect as NGF/γ-IFN treatment
Compared with controls at 7 days of incubation, the number of neutralizing FGFR2 antibody-treated cells decreased by 54.6%±6.2% and 47.9%±6.8% in Du145 and PC-3M cells, respectively. The CFE was decreased by 46.2%±9.6% and 38.7%±11.0%, respectively. CD44+ integrinα2β1+ cells decreased by 32.7%±6.5% and 27.6%±5.1%, respectively, and AR+ cells were 7.4%±1.6% and 4.6%±0.8% in Du145 and PC-3M cell lines, respectively. After the addition of DHT, the proliferation rate of the cells increased by 7.4%±2.7% and 5.7%±3.4% in both cell lines.
NGF/γ-IFN downregulates FGFR2 III-c and inhibits AKT signaling
The expression of FGFR2 isoforms was examined using real-time PCR. After NGF/γ-IFN treatment, FGFR2 III-b and FGFR2 III-c were downregulated by 0.8-fold and 7.4-fold in Du145 and 1.06-fold and 3.76-fold in a PC-3M cell line, respectively (Fig. 4B). With NGF/γ-IFN treatment, the phosphorylation of AKT was inhibited at Serine 473 while total AKT expression was not significantly altered. (Fig. 4C).
NGF/γ-IFN inhibits prostate cancer in vivo
PC-3M-luc-C6 cells (1×106), which are stably expressed luciferase, were subcutaneously transplanted into NOD/SCID mice (n=24). Bioluminescence imaging was used to detect tumor growth after cell transplantation. In the PBS- (n=6) and NGF- (n=6) treated groups, the pulmonary metastasis of cancer cells was detected on day 57 post-transplantation, while in the NGF/γ-IFN- (n=6) treated group (Fig. 5), only small-size tumors were observed (Fig. 5). This indicated that NGF/γ-IFN administration inhibited prostate tumor growth.
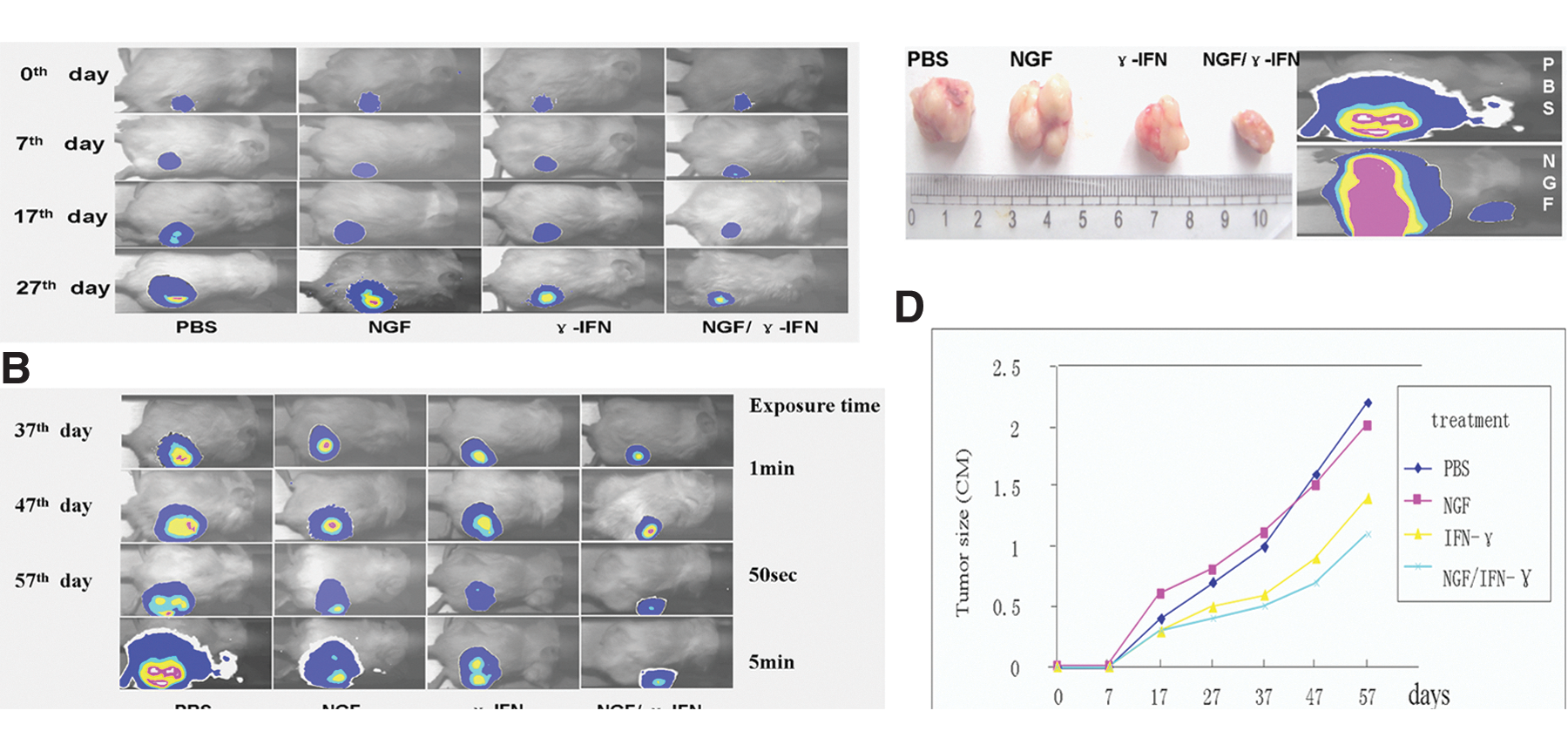
Bioluminescence imaging of transplanted prostate cancer in vivo.
Discussion
The mechanism responsible for changing prostate cancer into an androgen-independent type remains unclear. The treatment of AIPC remains a challenge for urologists. One possible explanation for its formation is that the original minority of CSCs becomes dominant after a long time of exposure with ADT. Prostatic CSCs neither express AR nor respond to androgen [25,26], but mature luminal cells express AR and can be stimulated by androgen. It is possible that castration and ADT can eliminate highly differentiated cells and CSCs that remain existent. AIPC may be the cell population that contains the majority of undifferentiated prostate CSCs. The blockage of CSC differentiation may be a cause of AIPC formation; therefore, once the differentiation pathway is recovered, CSCs in AIPC would be differentiated, and AIPC may be reverted to an androgen-sensitive type and be easily treated. Prostate CSC markers CD44 and integrinα2β1 were highly expressed in the AIPC cancer cell lines PC3 and DU145 [27]. In our previous report [24], the proportion of cancer stem-like cells in DU145 is higher than that of primary cancer, suggesting that AIPC is associated with CSCs.
As the type I AIPC cell lines, Du145 and PC-3M cannot entirely represent AIPC in patients due to a lack in an AR, but they are the appropriate model since the cell lines scarcely react to androgen.
After Du145 and PC-3M cells are chronically exposed to the NGF/γ-IFN environment for 1 week, the number of CD44+ α2β1+ cells dramatically decreased along with CK5 +cells, suggesting that CSCs were either differentiated or induced to apoptotic death. Meanwhile, both the proliferative potential and CFE declined, and AR was re-expressed in each cell line. Noticeably, AR was only expressed in α2β1- cells, confirming that AR was only expressed in more differentiated cells instead of stem cells or progenitor cells. When androgen was added into NGF/γ-IFN-treated cells, the proliferation of cells was inhibited rather than increased, a highly unexpected result. These results suggest that the inhibition of cell proliferation was due to the activation of AR, which plays an antiproliferation role in the particular cytokine environment. However, the function of AR induced by either NGF or γ-IFN alone is apparently different than with AR induced by NGF/γ-IFN. The mechanisms are unclear, and there is a possibility that some molecules associated with AR are altered by the treatment of NGF/γ-IFN, but not by either NGF or γ-IFN alone. Nevertheless, this finding demonstrates that the different cytokine environments may induce divergent functions of AR, and may provide a clue for the mechanisms presented when young males rarely have either benign prostatic hyperplasia or prostate cancer. The cytokine environment in a young man is different from that in aged males [28]. With aging, changes in the cytokine environment may influence differentiation of the prostate and stimulate the function of AR. NGF and IFN are associated with age and androgen levels [19,21]; thus, NGF/γ-IFN may be an important portion of the cytokine environment.
Our in vivo experiments showed that NGF/γ-IFN inhibited tumor growth in NOD/SCID mice, which are immunodeficient animals without T, B, and NK cells. The inhibitory effect of NGF/γ-IFN is unlikely due to a manipulation of the host's acquired immune system. In this case, a cytokine environment may play a more important role in inhibiting tumor growth.
NGF/γ-IFN altered the expression levels of certain genes, making TERT, Wnt1, Wnt2, and FGFR2 noticeable. TERT repairs telomeres to keep the proliferative potential, and is also a pluripotency-associated gene and a stem cell marker [29,30]. Wnt1 and Wnt2 are members of the wingless-type MMTV integration-site family, which is associated with the maintenance of an undifferentiated status of cells. Activation of Wnt1 induces a complex signaling cascade that ultimately leads to the increased expression of over 50 genes. Wnt and TERT are able to regulate each other [31], and the downregulation of Wnt and TERT may be the result of differentiation of the prostate CSCs in the AIPC cell lines.
The downregulation of FGFR2 expression is in agreement with the differentiation of the prostate CSCs, indicating that NGF/γ-IFN worked through the downregulation of FGFR2. FGFR2 is a member of the tyrosine kinase receptor family, which is expressed on the membrane and binds with FGF4, FGF7, FGF8, or FGF10. It plays a crucial role in signal transduction and development, and either promotes or inhibits differentiation in tumors [32 –35]. To examine the FGFR2 pathway, we used an anti-FG.FR2-neutralizing antibody to block the function of the FGFR2 receptor. Compared with controls, the FGFR2-blocked cells were further differentiated and had a lower proliferative potential, similar to the NGF/γ-IFN-treated cells. However, the gene expression and cell nature of FGFR2-blocked cells were not always consistent with that of the NGF/γ-IFN-treated cells. AR re-expressed in the FGFR2-blocked cells showed no anticancer function, suggesting that other mechanisms may also be involved in the NGF/γ-IFN pathway. Nevertheless, this study suggested that NGF/γ-IFN induced differentiation of CSCs mainly through inhibiting FGFR2 signaling. On the contrary, one study found that in primary prostate cancer cells, FGFR2 plays the role as a proliferation inhibitor and as a differentiation inductor [36]. We further investigated the isoforms of FGFR2: the FGFR2 IIIb and IIIc. Results indicated that NGF/γ-IFN mainly decreased the expression of FGFR2 IIIc and resulted in a reduction of phosphorylated AKT at Serine 473. These data suggest that the FGFR2 IIIc isoform affects phosphorylation of AKT and plays a role as a proliferation promoter and as a differentiation inhibitor. Furthermore, the anticancer functions of NGF/γ-IFN may result from multiple gene expression changes, including FGFR2, p-AKT, TERT, Wnt1, and Wnt2 that are induced by cytokines.
Aging [37] and ADT interfere with the androgen level and internal cytokine environment, and may lead to the proliferation of CSCs. The correction of the internal cytokine imbalances and the induction of differentiation of CSCs may offer new therapeutic designs for the treatment of AIPC.
Footnotes
Acknowledgments
We thank Prof. Lian-hua Yin, Prof. Si-feng Chen, and Dr. Xiao-bo Li for giving critical advice to our study. This research was supported by the NSFC (Natural Science Foundation of China, no. 30901705) and the Doctoral Fund of Ministry of Education of China (no. 20090071120024).
Author Disclosure Statement
The authors declare that there is no conflict of interest that could be perceived as prejudicing the impartiality of the research reported.