
Abstract
Embryonic stem (ES) cells have the potency to produce many cell types of the embryo and adult body. Upon transplantation into early host embryos, ES cells are able to differentiate into various specialized cells and contribute to host tissues and organs of all germ layers. Here we present data in the fish medaka (Oryzias latipes) that ES cells have a novel ability to form extra organs and even embryo-like miniatures. Upon transplantation as individual cells according to the standard procedure, ES cells distributed widely to various organ systems of 3 germ layers. Upon transplantation as aggregates, ES cells were able to form extra organs, including the hematopoietic organ and contracting heart. We show that localized ES cell transplantation often led to the formation of extra axes that comprised essentially of either host cells or donor ES cells. These extra axes were associated with the head region of the embryo proper or formed at ectopic sites on the yolk sac. Surprisingly, certain ectopic axes were even capable of forming embryo-like miniatures. We conclude that ES cells have the ability to form entire organs and even embryo-like miniatures under proper environmental conditions. This finding points to a new possibility to generate ES cell-derived axes and organs.
Introduction
The small laboratory fish medaka (Oryzias latipes), like the zebrafish, is an excellent model system for analyzing vertebrate development [6] and stem cell biology in lower vertebrates [7]. In this organism, we have derived several diploid ES cell lines [8,9], haploid ES cell lines [10], and a male germ stem cell line from adult spermatogonia [11]. One of the diploid ES cell lines, the medaka embryonic stem cell line 1 (MES1), has been characterized as being pluripotent in vitro and in vivo [8,9] and capable of directed differentiation into melanocytes by ectopic expression of the microphthalmia-associated transcription factor [12]. MES1 can activate the totipotency-specific mouse Oct4 promoter [20] and express 7 pluripotency genes examined [13]. MES1 retains the developmental pluripotency after stable gene transfer and long-term drug selection [15]. Upon transplantation into host embryos at the blastula stage, MES1 cells have a 100% efficiency of chimera formation and contribute into many functional cell types of various organs such as the heart [8,9,14 –16]. Remarkably, even in interordinal chimeras, MES1 cells can differentiate into various cell types by adopting the zebrafish host program [17].
Given the therapeutic promise of ES cells, this study was aimed at demonstrating the full potential of ES cells by using externally developing embryos of medaka for chimera formation. We show that medaka ES cells can form extra organ systems, axes, and even embryo-like miniatures in an environment of uncompromised diploid host embryos.
Materials and Methods
Fish
Work with fish was carried out in strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Advisory Committee for Laboratory Animal Research in Singapore and approved by this committee (Permit Number: 27/09). Medaka strains af and orange were maintained under an artificial photoperiod of 14-h light to 10-h darkness at 26°C [8,14,17]. Embryogenesis was staged according to Iwamatsu [18].
Plasmids
Plasmid DNA was prepared using the Qiagen Midi plasmid kit (Qiagen). Plasmids pCVpf and pCVpr were described [19], with the former expressing the fusion PF between P (the puromycin acetyltransferase; Pac) and F (green fluorescent protein; GFP), and the latter the fusion PR between Pac and R (the red fluorescent protein; RFP).
Cell culture
The medaka ES cell line MES1 was used as the transplantation donor throughout. It was maintained in ESM4 on gelatin-coated substrata as described [8]. MES1 cells transgenic for GOF18geo (GOF18geo-MES1) were produced and maintained in the presence of G418 (200 μg/mL) to inhibit spontaneous differentiation as described [15]. To label GOF18geo-MES1 cells genetically, they were transfected with pCVpf or pCVpr and selected in the presence of both G418 (400 μg/mL) and puromycin (0.5 μg/mL) [15,20]. The cell line iC6 was derived from the adult medaka testis by using the procedure described for SG3 [11], which consisted of differentiated somatic cells.
For use as a transplantation donor, cells were harvested by trypsinization and stained with fluorescent vital dyes PHK26 (red; Sigma) or PKH67 (green; Sigma) for 5 min at 2×10−6 M with 1×107 cells/mL in Diluent C (Sigma). After stopping the labeling reaction by adding an equal volume of serum, cells were harvested by centrifugation and washed 3 times with phosphate-buffered saline (PBS) to remove the unbound dye. PKH67 and PKH26 staining has little adverse effect on cell viability and phenotype, as single cells after staining maintained the capability of adherent growth.
To analyze the donor cell fate in chimeric embryos in more detail, cells and cell aggregates were dissociated from 4-day-old chimeras. To this, chimeric embryos were washed extensively in PBS-containing antibiotics and prickled a hole by using fine tweezers on the yolk sac to release yolk material. The embryos were transferred into a 1.5-mL Eppendorf tube containing cold trypsin solution. After treatment on ice for 15∼30 min, the embryos were incubated in 100 μL of trypsin solution for 2 min at room temperature for enzymatic cell dissociation. Trypsinization was stopped by adding 100 μL of ESM2, and single cells and cell aggregates were prepared by vigorously pipetting the embryos without producing air bubbles. After centrifugation for 2 min at 6000 rpm (Centrifuge 5415C; Eppendorf), the cells and tissues were collected, pipetted in ESM2, and seeded onto gelatin-coated 24-well plates. During 24 h of culture at 28°C in ESM2 containing puromycin (1 μg/mL), many cells were killed by the drug, and the remainder survived and attached to the substrata. The cells were washed in PBS with 3–5 changes so that any unattached aggregate is washed away, and further cultured for 2 h for recovery before observation and photography.
Cell transplantation
Recipient embryos were collected at the 2-cell stage, dechorionated, and transplanted at the midblastula stage [10], with a minor modification. Briefly, donor cells were loaded at a higher density and injected quickly as single cells (type I transplantation) or gently as an aggregate (type II transplantation) or a thread-like mass into localized areas along the blastoderm (type III transplantation), at 200–500 cells per host embryo.
Microscopy
Results
Parameters of chimera formation
We and others have revealed strain differences in ES cell derivation and chimera formation in mouse [21] and medaka [14,16], and the effect of ablating donor ES cell differentiation on the efficiency of chimera production in mouse [22] and medaka [9,16]. To understand in more detail the capacity and behavior of ES cells in vivo, we further analyzed parameters of chimera formation by testing different conditions of donor ES cell transplantation (Fig. 1A). Type I results in wide distribution of 100–200 cells, (Fig. 1B) which has routinely been used to test the potential of ES cells for differentiation in vivo [9,14,16,17]. Type II and type III deliver a high density of ES cells into a round (Fig. 1C) and thread-like area (Fig. 1D).
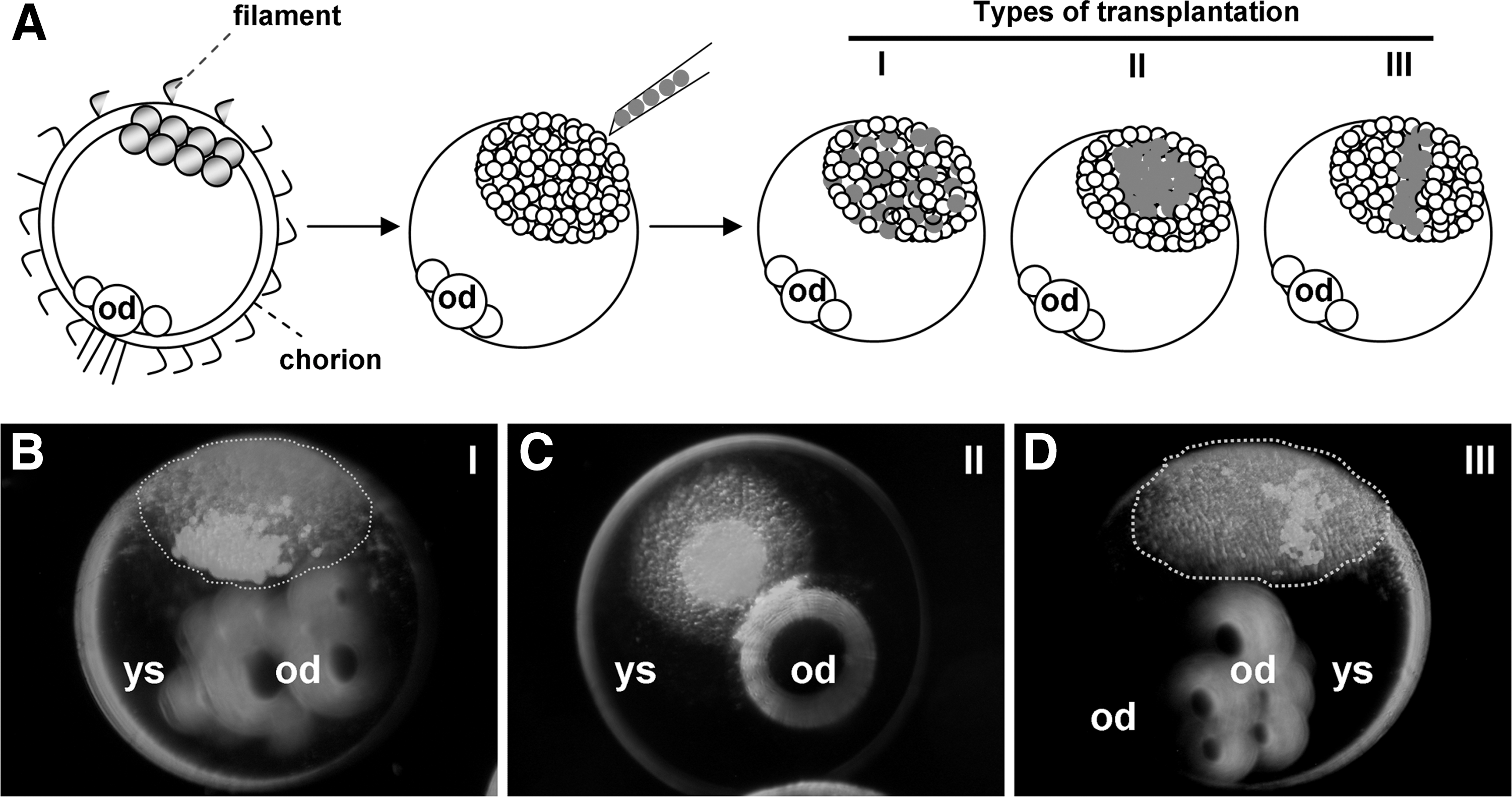
Types of embryonic stem (ES) cell transplantation.
Wide distribution of ES cells to host organs
Transplantation of ∼200 MES1 cells per host led to a 100% efficiency of chimera formation and normal and near-normal development in the overwhelmingly majority of chimeras, conforming to previous reports [9,14,16]. Upon type I transplantation, these ES cells most frequently exhibited wide distribution in various embryonic compartments and organs such as the eye, heart, trunk, gonad, blood islands, and yolk sac (Fig. 2A), essentially as described previously [9,14]. A total of 51 chimeras from type II transplantation, ∼200 MES1 cells were observed in detail. Up to 90% (n=46) exhibited normal development, in which the bulk of donor cells was concentrated in one area such as the brain, with the remainder being dispersed in various areas and organs (Fig. 2B). Contribution of transplanted MES1 cells to terminally differentiated, functional cells was apparent in several organ systems. Specifically, MES1-derived cardiomyocytes were clearly seen as GFP-positive contracting cells in the chimeric atrium and ventricle of the beating heart (Supplementary Movie S1; Supplementary Data are available online at
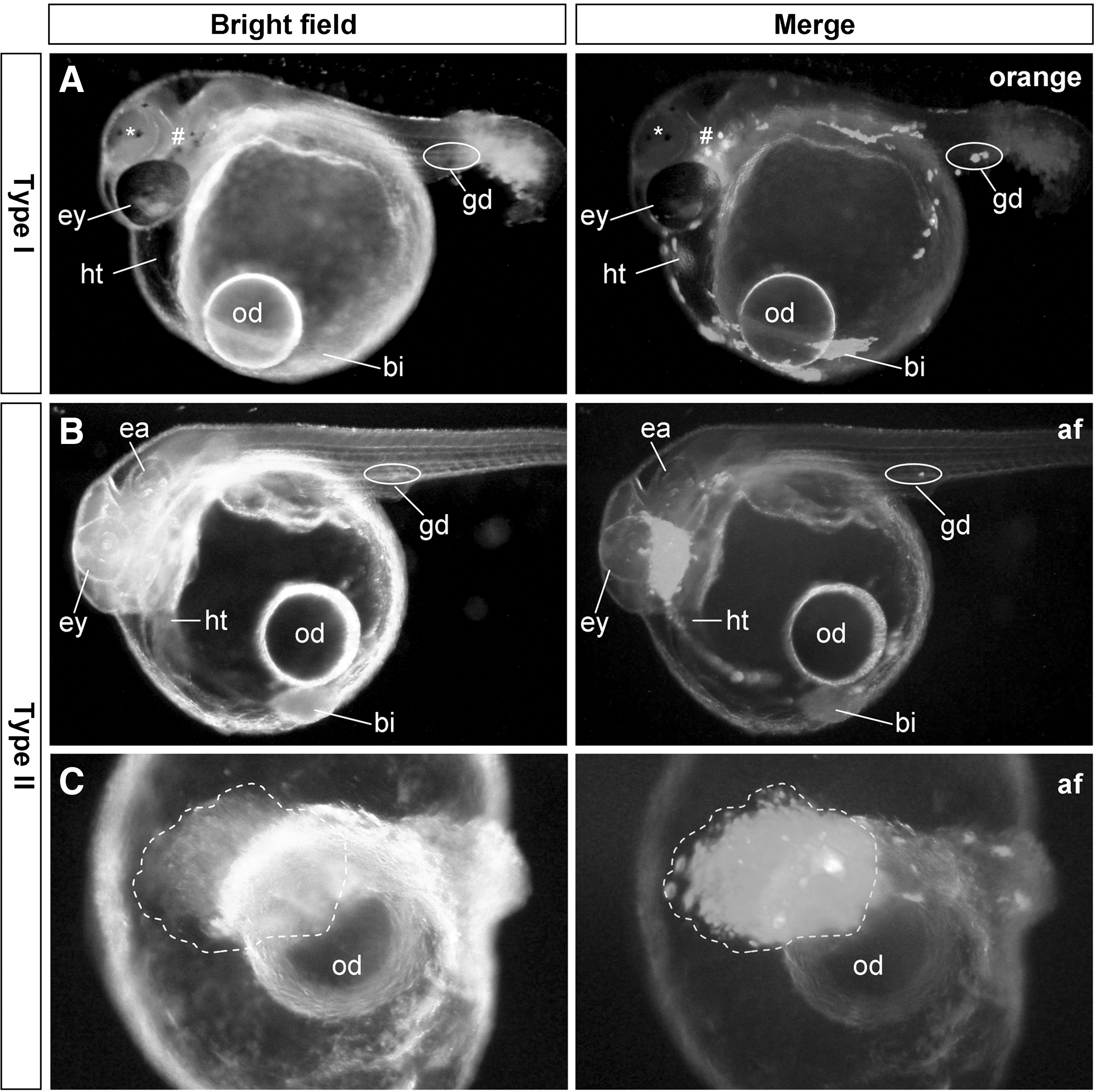
Chimera formation from 2 transplantation strategies.
MES1 derivatives in chimeric embryos
Wide distribution of MES1 cells and their functional contribution to several easily identifiable organs/lineages of the developing chimeric embryo provoked us to examine the fate of MES1 derivative in more detail. To this end, single cells or cell aggregates were dissociated from chimeras at 3–7 days postfertilization (dpf). At 24 h of culture, heterogenic cell populations of GFP-positive and negative cells are visible. MES1 derivatives from 4-day-old chimeric embryos exhibit a phenotype of differentiated cells, (Supplementary Fig. S2) which is distinct from that of undifferentiated MES1 before transplantation. Many cells were live as evidenced by their attachment to the culture plates. Derivatives from type I transplantation chimera displayed wide distribution (Supplementary Fig. S2A), whereas those of a type-II transplant mainly resided in a cell cluster (Supplementary Fig. S2B). Overall, MES1 derivatives show viability and differentiation in the host.
Production of ectopic organs by ES cells
Our previous studies have shown that the medaka midblastula embryo is permissive for transplantation of ∼100 MES1 cells [9,16]. This permissive number can be increased to ∼200 when the donor ES cells were maintained in the presence of G418 to suppress spontaneous differentiation [15,20]. Transplantation of 200–500 these ES cells into a localized area was compatible with a largely normal embryogenesis. In the first series of experiments, we introduced ∼200 MES1 cells as individual aggregates per host. Of 321 transplants, 74 survived to the organogenesis stage at 3 dpf. Eighteen had MES1-derived hematopoietic islands that were easily identified by the red color of hemoglobin, location on the yolk sac (the position of embryonic hematopoietic islands), and more importantly, the production and release of red blood cells into circulation (Fig. 3A and A’). Those blood islands consisted predominantly of either MES1 derivatives (bi1) or host cells (bi2; Fig. 3B). Furthermore, 2 of the embryos had ectopic hearts consisting predominantly of MES1 derivatives (Fig. 3C–F). In both embryos, the extra heart had blood vessels that were connected to the major heart, and it was similar in size to the major heart below the head region (Fig. 3C and E). Besides a shared pool of blood cells, the extra and major hearts displayed synchronous contraction of rhythmical waves (78 pulses/min), demonstrating that the ES cell-derived extra heart functions similarly as the major heart. Taken together, these observations show that ES cells are capable of producing entire organ systems in developing embryos when transplanted at high cell numbers.
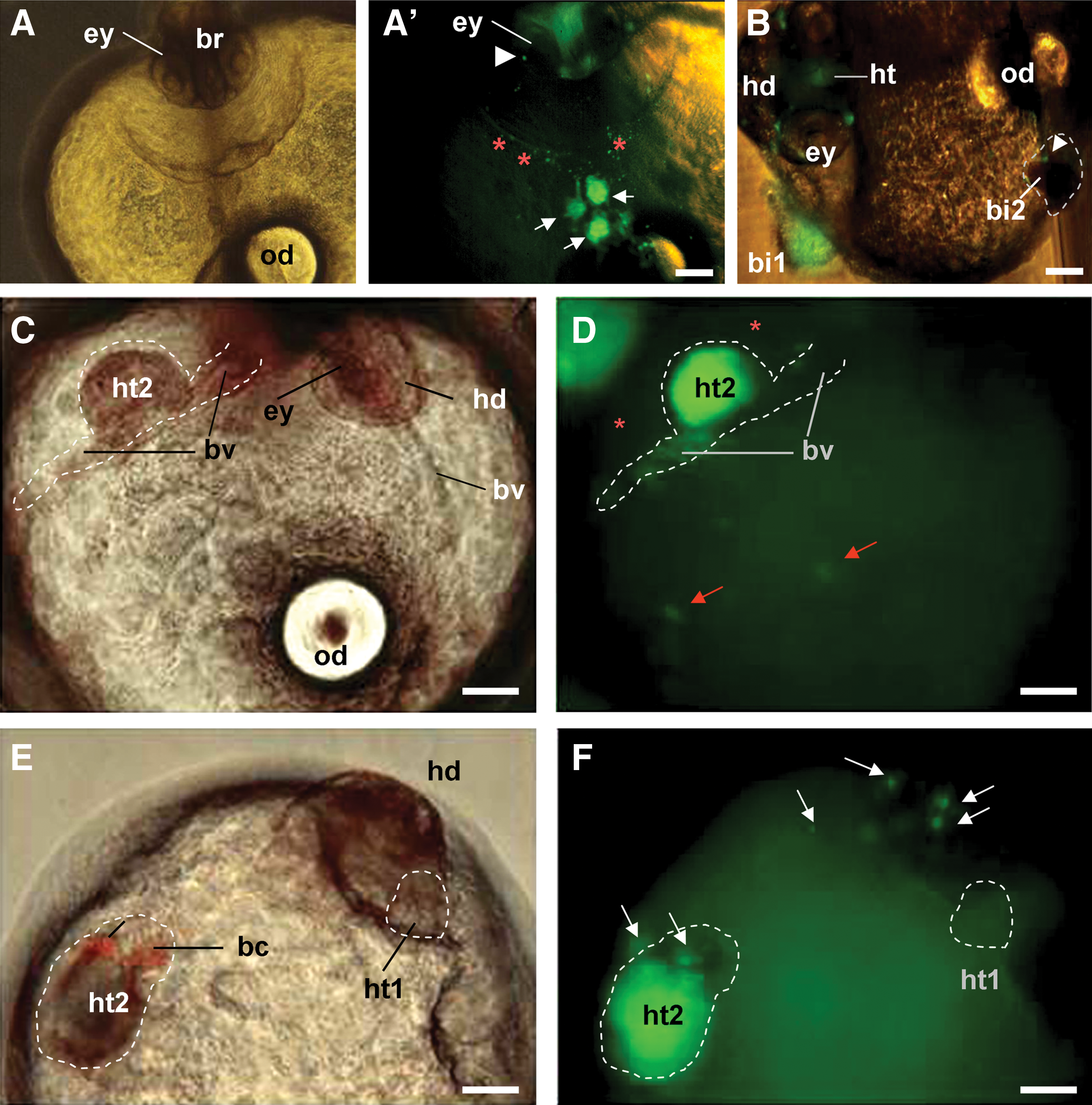
Generation of ectopic organ systems.
Production of extra axes by ES cells
In the second series of experiments, ∼500 MES1 cells were introduced as several aggregates into a small area within the blastoderm (Fig. 1B). This led to a high frequency of extra axis formation. Of 177 transplants, 107 survived to 1 dpf, of which 25 (24%) had secondary axes. Two major types of axes were observed. The majority of extra axes were physically connected to the embryo proper (type 1). They were similar to (Fig. 4A), or smaller than (Fig. 4B,C), the major axis in size (Fig. 4E). These extra axes consisted of posterior trunk beginning from the hindbrain, followed by the otic vesicle (Fig. 4B,D). Type-2 axes were ectopic, much smaller than the embryo proper, and made up predominantly of ES cell derivatives (Fig. 4F,G). All extra axes appeared to be at the same developmental stages as the major embryo.

Generation of extra axes and embryo-like miniatures from MES1 cells.
Production of embryo-like miniatures by ES cells
In the third series of experiments, ∼500 MES1 cells were introduced as a thread-like aggregate along the blastoderm (Fig. 1C). This transplantation resulted in severely disturbed gastrulation and thus massive death at early stages. From a total of 198 transplants, only 17 survived beyond 3 dpf. Five of them had extra axes, 3 of which were ectopic. Intriguingly, 2 of the 3 ectopic axes developed into embryo-like miniatures. One miniature-bearing embryo survived until 7 dpf. The ectopic miniature showed many features of an entire embryo (Fig. 4H–J), as evidenced by apparent anterior–posterior patterning, dorsoventral patterning, and left–right symmetry, and all the major embryonic compartments, namely the head, trunk, and tail. More importantly, the miniature had many well-developed cells, tissues, and organs. These included muscles, somites, the neural tube, notochord, pigment cells, and eye-like structure consisting of the seemingly retina and lens. We found that the tail was capable of movement, indicating the functionality of the nervous cells and muscles. We were not able to identify internal organs. Notably, the embryo-like miniature exhibited wild-type (black) pigmentation in the body and eye-like structures (Fig. 4I–J), demonstrating its origin from transplanted ES cell donors. In summary, MES1 is apparently capable of forming many organ systems and even whole-embryo-like miniatures in vivo.
Blastula transplantation of differentiated cells
For a comparison as differentiated cells, the cell line iC6 derived from the medaka adult testis was used for blastula transplantation at passage 67. This cell line consists of a apparently differentiated phenotype, including a large size and flat shape (Supplementary Fig. S3A). Staining with the red fluorescent dye PHK26 had little adverse effect on survival and growth, as stained cells retained the ability for attachment and growth (Supplementary Fig. S3B). A similar result was obtained also with the green dye PKH67 (data not shown). The labeled cells were subjected to type I and type II transplantation. In the first experiment, a total of 67 embryos survived type II transplantation (Supplementary Fig. S3C and D). Of 54 survivors at 1 dpf, 37 (68%) had a secondary axis. Until 4 dpf, 43 survived with or without extra axes (Supplementary Fig. S3E). In the second experiment, a total of 75 embryos survived type I transplantation (Supplementary Fig. S4A), and 58 transplants survived at 1 dpf. The majority (86%) of these chimeras apparently showed gastrulation arrest and defects, including epigastrulation (Supplementary Fig. S4B) and incomplete gastrulation (Supplementary Fig. S4C). Only 7 (9%) of the type I transplants survived at 2 dpf, and none of them appeared normal, because they were much smaller than normal embryos due to yolk collapse (Supplementary Fig. S4D). No ectopic axis and embryo-like miniatures were obtained by transplantation of differentiated cells. It follows that transplantation of differentiated cells is nonpermissive for normal development.
Discussion
In this study, we have used externally developing embryos of medaka to assess the developmental potential of ES cells. We show the participation of ES cells in a wide variety of embryonic compartments and organ systems of 3 germ layers, conforming to our previous observations [9,10,14,16,17]. We reveal that ES cells have the unusual ability to form ectopic organs and even whole-embryo-like miniatures. Our analyses of the ability of ES cells to produce ectopic organs has focused on the hematopoietic island, blood vessels, and heart, because they are easily identified during early organogenesis stages by their unique structures, locations, and functions (e.g., circulation and contraction). We cannot rule out that these ES cells could also form other extra organs. We reveal that certain ectopic organs and ectopic axes are substantially of donor ES cell origin, as evidenced by donor-specific GFP expression. Some of the extra organs of donor ES cell origin appear to be functional. For example, the ectopic heart can contract with one and the same rhythm of the normal heart. More convincingly, the extra embryo-like miniature developing on an albino host background exhibits wild-type pigmentation, which demonstrates a functional contribution of wild-type donor ES cells. Therefore, we conclude that ES cells are capable of participating in the formation of extra organs and entire embryo-like miniatures in medaka.
In lower vertebrates, β-Catenin and Nodal play an essential role for axis development. For example, extra axes can be induced by injection of β-Catenin antibodies into ventral blastomeres in Xenopus [23], and overexpression of β-Catenin [24] and Nodal [25] in zebrafish and β-Catenin overexpression in medaka [26]. Most recently, Nanog-like is able to activate the Nodal pathway to induce the endoderm in zebrafish [27]. In medaka, Nanog is essential for stem cell proliferation [28], and MES1 and even haploid ES cell lines express a detectable level of Nanog RNA [10,13]. It remains to be determined whether the ability of transplanted MES1 cells to produce extra axes is due to Nanog expression, as it is unclear at present that if Nanog could induce the Nodal signaling for axis duplication in this organism. Alternatively, extra axis formation may be a consequence of the inducing activity of differentiated cells. In support of this idea is the observation that transplantation of fully differentiated cells results in even a higher frequency of extra axis formation. In this study, the ES cells used for transplantation were free of apparent differentiation by drug ablation of terminally differentiated cells on the basis of the mouse Oct4-driven expression of drug resistance [20]. However, this approach does not ablate early-differentiating ES cells, because residual drug resistance will allow them to escape from drug selection.
Cell transplantation is a powerful tool in medicine (e.g., blood and bone marrow transplantation) and basic research. Transplantation into organs of growing and adult animals has been reported for both progenitors such as mesoangioblasts [29,30] and differentiated cells such islet cells [31]. For transplantation into early developing embryos, stem cells of various origins are usually used as donors. In this study, we reveal that in contrast to ES cell transplantation that is compatible with normal development, transplantation of a differentiated cell line is nonpermissive for normal embryogenesis in medaka. Future work is needed to examine whether incompatibility is common to transplantation of various differentiated cell types.
We have previously shown that MES1 is highly efficient for production of somatic chimeras [9,14,16]. The present study corroborates and extends these observations by demonstrating the ability of MES1 to form entire organs and embryos. It has been reported that mammalian ES cells can differentiate into various cells of all germ layers in vitro and in vivo [1], and can form organ-like structures such as eye and gut under defined culture conditions [2 –4]. Our work substantiates these reports by demonstrating that medaka ES cells in externally developing embryos are able to generate extra organs such as the heart that is similar to the major heart in gross morphology and function. Our striking finding that ES cells are able to generate entire organs, extra axes, and even embryo-like miniatures in the environment of the host embryo has an important implication for transplantation medicine. At present, it is not known what environmental conditions and mechanisms govern the formation of entire organs from ES cells. However, our results points to new possibility for ES cell-based organ formation as a novel resource of organs for transplantation medicine. Although it remains to be determined whether mammalian ES cells also have the ability to form organs or even full embryos, it has been reported that ES cells alone are able to support fetal development in the mouse [32,33].
Conclusion
We reveal that normal embryogenesis after blastula transplantation is compatible with pluripotent stem cells such as ES cells, but nonpermissive to differentiated cells. Furthermore, our results show that the way of cell transplantation has a profound effect on the distribution and behaviors of donor cells. Most importantly, we demonstrate the ability of ES cell transplantation to produce extra organs, extra axes, and even embryo-like miniatures by using externally developing medaka embryos as the host. These results will shed new insight into chimera biology and potential of ES cells potential for organ production.
Footnotes
Acknowledgments
We thank J. Deng for fish breeding and CM. Foong for laboratory management. This work was supported by the National Research Foundation Singapore under its Competitive Research Program (CRP award no. NRF-CRP7-2010-03 and NRF-CRP 002-082).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.