
Abstract
Human somatic cells can be reprogrammed into induced pluripotent stem cells (iPSCs) by ectopic expression of key transcription factors. iPSCs have been generated from a variety of cell types. However, iPSC induction from human myoblasts has not yet been reported. Human primary skeletal myoblasts can be cultured from diagnostic muscle biopsy specimens, and thousands of lines are frozen and stored in biobanks, and are a valuable source for iPSC-based etiological and pathogenic studies. Our aim was to generate iPSCs from human skeletal myoblasts enriched from muscle biopsy samples. We used retro- or Sendai virus vector-mediated reprogramming of enriched human myoblasts from 7 donors. We show that stable iPSC lines can be generated from human myoblasts at efficiency similar to that of fibroblasts when appropriate media is used, and the efficiency of the feeder-free iPSC generation can be significantly improved by inhibitors of histone deacetylase (sodium butyrate) and TGF-β signaling (SB431542).
Introduction
Muscle satellite cells are progenitor cells located between the basal lamina and the plasma membrane of the multinucleated myofiber. They are normally quiescent in adult skeletal muscle. After an injury or during growth of an adult skeletal muscle, dormant satellite cells become activated and are involved in muscle growth and regeneration [19]. Myoblasts (activated satellite cells) are able to proliferate and fuse to form long multinucleated myotubes. Myoblasts are regularly obtained from patients with skeletal muscle disorders, and can be sorted from fibroblasts using antibody against neural cell adhesion molecule (NCAM). In the presence of mitogen-rich culture medium and a high concentration of serum, they are able to grow for a limited number of passages as an undifferentiated population expressing MYOD and MYF-5 [20]. Thousands of human myoblast lines are stored frozen in biobanks around the world. These cells could serve as a valuable source of iPSC for etiological and pathogenetic studies. However, to our knowledge the generation of iPSCs from human myoblasts has not been reported.
Reprogramming is in general a slow and inefficient process, which can be enhanced by the use of small molecule inhibitors [21]. These molecules are known to affect for example the epigenetic state of the cells, by modifying chromatin [22], or signaling context by targeting pathways, such as MAPK and TGF-β [23]. However, these inhibitors are likely to have divergent effects depending on the starting cell type and the method of reprogramming.
Here, we report efficient iPSC generation from human skeletal muscle biopsy-derived myoblasts and improvement of efficiency with the use of small molecule inhibitors.
Materials and Methods
Cell culture
Human fibroblasts and myoblasts (Supplementary Table S1; Supplementary Data are available online at
Preparation of retroviruses and transduction
293-GPG packaging cells [24] were transfected at the confluence of 80%–90% with 5 individual pMXs-cDNA vector (pMXs_Oct4, pMXs_Sox2, pMXs_Klf4, pMXs_cMyc, and pMXs_dsRed) or 2 tricistronic vectors (pMXs_Oct4_2A_Klf4_2A_GFP and pMXs_Sox2_2A_cMyc_2A_GFP) along with FugeneHD (Roche), according to the manufacturer's instructions. 24 h after transfection media was changed to the fibroblast growth medium. Viral supernatants were collected at days 4, 5, and 6, combined and filtered through 0.45 μm syringe filters (Millipore) prior to use.
Generation of iPSC
The Coordinating Ethics Committee of the Helsinki and Uusimaa Hospital District approved the generation of iPSC lines from the myoblasts and fibroblasts. Purified myoblasts and fibroblasts between passages 4 to 10 from the same donors were plated in 6-well plates at a density of 1×105 cells/well. Cells were transduced at days 0 and 1 with freshly collected virus supernatants containing equal volume amount of each of the 4 retroviruses. On day 3, transduced myoblast and fibroblast cells were washed with phosphate-buffered saline (PBS) and media changed to fibroblast growth medium. Six days after infections, cells were dissociated by TrypLE Select (Invitrogen) and replated on cell culture plates coated with mitomycin-treated MEFs. Next day, the medium was replaced with hES medium. The medium was changed every other day. Colonies were picked 24–30 days after transduction and transferred to 24-well plates on mitomycin-treated MEFs. This stage is counted as passage 1.
For generation of iPSCs using Sendai viruses (SeV, Cytotune; Life Technologies) we followed manufacturer's instructions. Briefly, purified myoblasts and fibroblasts were plated in 6-well plates at a density of 2×105 cells/well. At day 0, cells were transduced with SeVs in either myoblast or fibroblast growth medium. At day 6, the cells were dissociated using TrypLE select (Invitrogen) and replated to mitomycin C-treated MEF-coated plates. hES growth medium was changed every other day. iPSC colonies were picked up 18–26 days after transduction and transferred to 24-well plates on mitomycin-treated MEFs.
Inductions using small molecules in feeder-free conditions
For inductions using inhibitors, cells were re-plated on day 6 after transduction to Matrigel-coated wells and media was changed to hES medium (knockout DMEM; Gibco), supplemented with 20% KO-serum replacement (Gibco), 1% GlutaMAX (Gibco), 0.0915 mM β-mercaptoethanol, 1% nonessential amino acids (Gibco), and 6 ng/mL bFGF (Sigma), supplemented with inhibitors. The following factors were tested: 0.25 mM sodium butyrate (NaB; Sigma), 0.5 mM valproic acid (VPA; Sigma), 2 μM SB431542 (Selleck), 0.5 μM PD0325901 (Selleck), and 1 mM lithium chloride (Sigma).
Immunohistochemistry and alkaline phosphatase staining
Cells were fixed in 4% paraformaldehyde (PFA). To reveal nuclear antigens, the cells were permeabilized using 0.2% Triton X-100 (Sigma). Cells were then treated with ultra V block (Thermo). Immunohistochemical analysis of embryoid bodies (EB) was done from paraffin sections. To reveal antigenic sites, the rehydrated sections were treated in microwave oven in 1 mM EDTA, pH 8, for 3 min. Primary antibodies were against Tra1-60 (1:40, 90232; Millipore), SSEA4 (1:1,000, 90231; Millipore), OCT4 (1:1,000, sc-9081; Santa Cruz Biotechnology), TUJ1 (1:5,000, beta cell biology consortium), Brachyury (1:1,000, sc-20109; Santa Cruz Biotechnology), FOXA2 (1:500, sc-9187; Santa Cruz Biotechnology), Vimentin (1:2,000, M0725; Dako), NCAM (1:10), and Nanog (1:500, #4903; Cell Signaling). Secondary antibodies were AlexaFluor 488: donkey anti-goat (1:500, A11055; Invitrogen), donkey anti-mouse (1:500, A21202; Invitrogen), donkey anti-rabbit (1:500, A21206 and A21207; Invitrogen), and Horseradish peroxidase (1:200). Alkaline-phosphatase (AP) was detected using NBT/BCIP solution (Roche Applied Science). Nuclei were stained with DAPI (Vectashield, Vector).
Real-time polymerase chain reaction
Total RNAs were purified from cultured cells using NucleoSpin RNA II kit (Macherey-Nagel). DNA was digested in a separate reaction using DNaseI (Promega). Two micrograms of total RNA was used for reverse transcription reaction. Reverse transcription was performed using MMLV RTase (Promega), random hexamers (Promega), and oligoT (Promega) according to the manufacturer's instructions. 1 μl of cDNA was used for quantitative polymerase chain reaction (qPCR) and was performed with SYBR Green (Invitrogen, Molecular Probes S7563) in Corbett RotoGene. Primers for PCR are listed (Supplementary Table S2), and qPCR assays were performed in duplicates. Relative gene expression levels were normalized to GAPDH or Cyclophilin.
EB differentiation of iPSCs
iPSC colonies were dissociated using Collagenase IV into small pieces and transferred to 60 mm low attachment plates (Corning) in hES medium without bFGF. Dissociated cells were cultured in suspension for 14 days, with media changed every fourth day. EBs were fixed with 4% PFA for immunohistochemical analysis.
Flow cytometry
Cells were collected by TrypLE (Gibco) treatment, after which the cells were washed, counted, and suspended in FACS-buffer (5% fetal calf serum; Promocell, in PBS). The cells were suspended in PE Mouse Anti-human CXCR4 antibody (BD Pharmingen) or anti NCAM and incubated at room temperature for 30 min. After incubation, the cells were washed thrice with FACS-buffer and fixed with 0.1% PFA in FACS-buffer. Samples were run with FACSCalibur (BD Biosciences) by using CellQuestPro software (BD Biosciences).
Targeted differentiation into definitive endoderm and myotubes
iPSC lines were differentiated into definitive endoderm on mitomycin C-treated MEFs. At day 0, cells were washed with PBS and media changed to RPMI-1640 (Gibco) supplemented with Glutamax (Gibco), 2% B27, 100 ng/mL Activin A, 75 ng/mL Wnt3a, and 1 mM NaB. After that, medium was changed daily from day 1 to 7, RPMI-1640 supplemented with Glutamax, 2% B27, 100 ng/mL Activin A, 75 ng/mL Wnt3a, and 0.5 mM NaB.
To differentiate myoblasts into myotubes, myoblasts growth medium was replaced with DMEM (Lonza BE12-614F) supplemented with 2% horse serum and 1% Glutamax (Gibco). Media was changed every fourth day.
Teratoma formation
About 200,000 morphologically intact iPSCs were intratesticularly injected into male NMRI nude mice (Scanbur). The resulting tumors were collected 8 weeks after injection, fixed with 10% formalin, and hematoxylin and eosin stained. The experimental animal welfare committee of the District Government of Southern Finland approved the animal experiments.
Statistical analysis
Student's t-test was used for statistical evaluations of iPSC induction efficiency from myoblasts and fibroblast cells. Two-way ANOVA followed with Tukey's HSD post-hoc test was used to assess the efficiency of iPSC induction in the presence of small molecule inhibitors. A p-value<0.05 was considered statistically significant.
Results
Characterization of myoblasts enriched from skeletal muscle biopsy
Myoblasts were enriched from 7 skeletal muscle biopsies (Supplementary Table S1). Practically, all sorted cells expressed myoblast markers NCAM and Desmin and expressed PAX7 and myogenic determination factor genes MYF-5 and MYOD, as shown by qPCR (Fig. 1A–C and data not shown). The ability of enriched myoblasts to differentiate into myotubes was then tested with 2 myoblast lines (Myoblast 1 and 2). Both lines were found to differentiate into multinucleated myotubes and expressed NCAM (Fig. 1A, and data not shown).
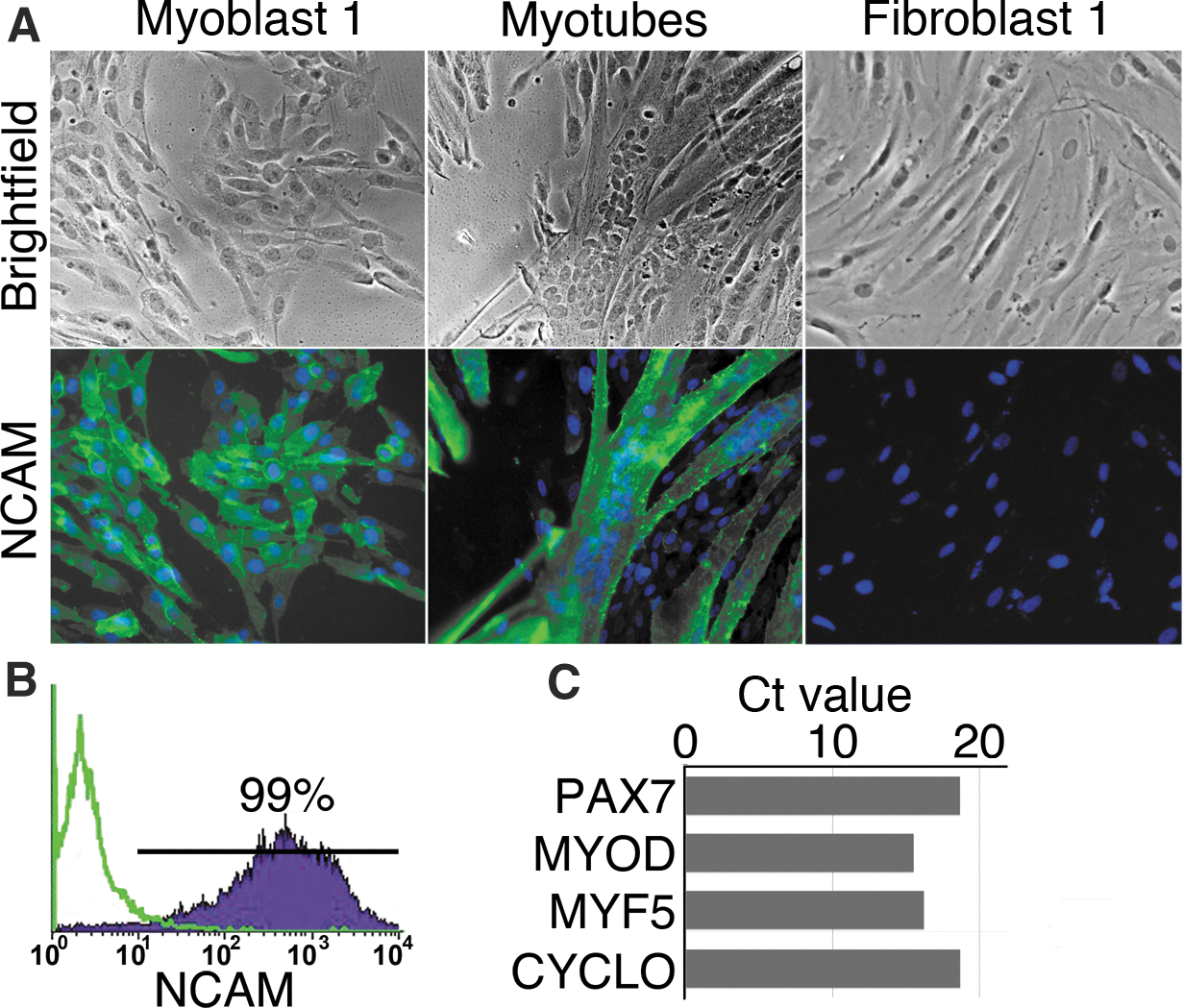
Characterization of myoblasts.
Reprogramming of skeletal myoblasts using retroviruses
We have observed that amphotrophic retroviruses routinely yield high transduction efficacy in human foreskin fibroblast (HFF) cells [25]. The transduction efficiency was tested with myoblasts to see if the retroviral vectors were also able to transduce skeletal myoblasts. Transduction of skeletal myoblasts and fibroblasts derived from the same donor, with retrovirally delivered vector-expressing dsRed, resulted in more than 60% of transduced cells expressing dsRed. We concluded that adult primary myoblasts and fibroblasts were transduced with similar efficacy (data not shown).
Retroviruses containing the “Yamanaka factors” OCT4, SOX2, KLF4, and c-Myc were used to reprogram isolated myoblasts and fibroblast cells. Schematic representation of the iPSC induction is shown (Fig. 2B). Myoblast-derived iPSC colonies started to appear 14–18 days after transduction and by day 23, iPSC colonies with a morphology similar to human embryonic stem cells (hESC) were picked and further propagated (Fig. 2A). AP activity and hESC marker Tra1-60 expression were seen in most of the colonies (Supplementary Fig. S1A).
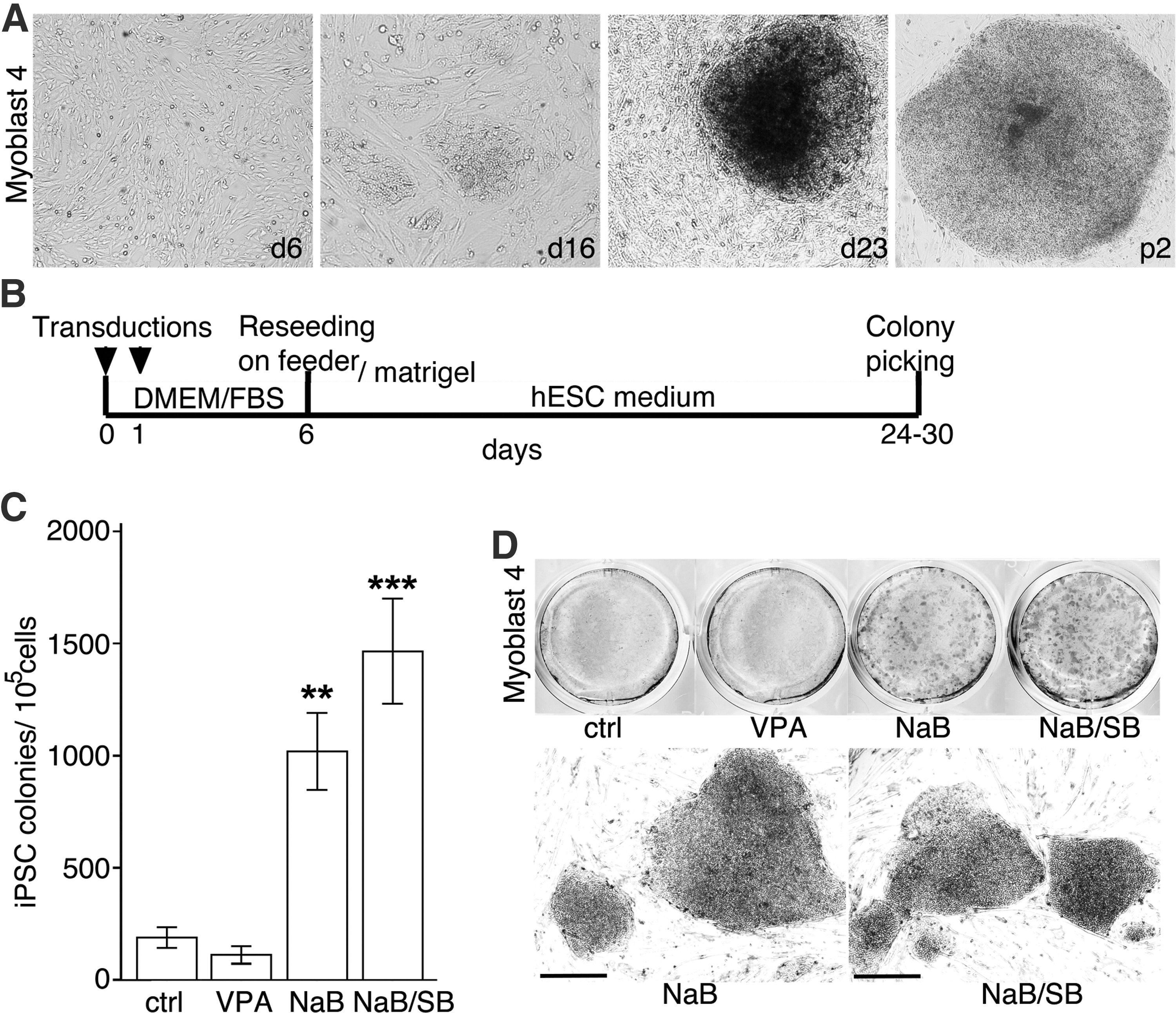
Retroviral reprogramming of myoblasts.
To compare iPSC induction efficiency we reprogrammed myoblast and fibroblast cells obtained from the same donor. iPSC colonies, with clear hESC morphology and positive for AP staining were counted. No significant difference (P=0.2) in reprogramming efficacy between fibroblasts (0.016%, n=4) and myoblasts (0.014%, n=4) cells was detected (Supplementary Fig. S1B).
Small molecule inhibitors significantly enhance the induction efficiency in feeder-free conditions
Histone deacetylase (HDAC) inhibitor, NaB (sodium butyrate) can be used to enhance the efficiency of retrovirally mediated iPSC induction from fibroblast cells [22]. To see if NaB has the same effect on the iPSC induction from skeletal myoblasts, induction efficiencies from the individual myoblast and fibroblast cell lines derived from the same donor were compared. Addition of NaB to myoblasts enhanced iPSC induction efficiency by more than 100-fold compared with induction without NaB (Supplementary Fig. S1B). However, no significant difference (P=0.29) was observed in the iPSC induction efficiency between myoblast (1.2%; n=10) and fibroblast (1.7%; n=10) cell lines (Supplementary Fig. S1B).
To see if the addition of other small molecule inhibitors improves the efficacy of iPSC induction from myoblast cells in feeder-free conditions, ALK4/5/7 inhibitor SB431542 (SB), MEK inhibitor PD0325901 (PD), nonspecific GSK3 inhibitor lithium chloride (LiCl), and HDAC inhibitor VPA, all reported to enhance the efficiency of iPSC inductions [22,26,27], were tested in combination with NaB (Fig. 2C, D, and Supplementary Fig. S2). Addition of SB to the NaB-containing medium enhanced the induction efficiency compared with NaB only. Further addition of PD or LiCl to the mixture did not further improve the efficiency. Addition of the HDAC inhibitor VPA instead of NaB did not increase the induction efficacy.
Generation of myoblast-derived iPSCs using integration-free method
Generation of integration-free iPSCs is essential to reduce the risk of mutagenicity caused through integration of reprogramming factors and to reduce potential alterations in differentiation potential caused by the integrated factors. SeV-mediated factor delivery can be used to efficiently generate integration-free iPSCs from fibroblast cells [6]. To see if integration-free skeletal myoblast-derived iPSCs can be generated using the same method, myoblasts were transduced with SeV and cultured either in myoblast- or fibroblast growth medium for the first 6 days, followed by change of the medium to hESC growth medium for the rest of the induction protocol (Fig. 3B). When the myoblast medium was used for the first 6 days of reprogramming, observed iPSC generation efficiency was low, 0.04% (n=3). However, when myoblasts were maintained in fibroblast growth medium for the first 6 days after transduction, the induction efficiency increased to 0.75% (n=3) and was comparable to that of fibroblast reprogramming observed in our laboratory (Fig. 3C). iPSC-like colonies appeared already by day 12 and were picked 18–24 days after transduction for further passaging.
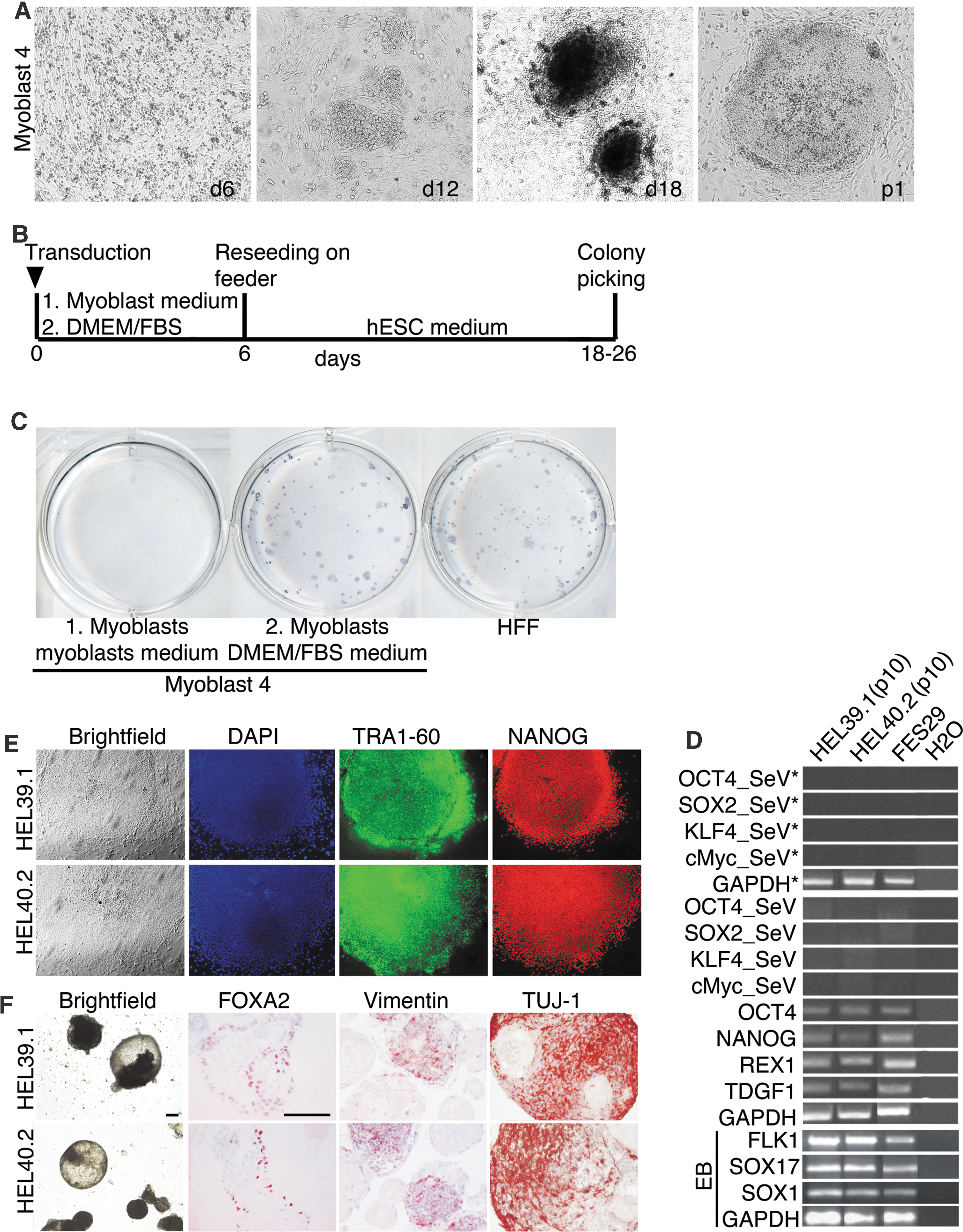
Characterization of integration-free iPSC derived by SeV vectors.
Skeletal myoblast-derived iPSCs express hESC markers
Eventually, iPSC lines from 7 skeletal myoblasts were derived (Table 1). All iPSC colonies were assessed for short tandem repeats and matched to parental lines (Supplementary Fig. S3A). Karyotype analysis of iPSC lines 9A1 (passage 16) and 9A2 (passage 12) showed a normal karyotype (46,XX). Line 9A3 (passage 14) was mosaic for a balanced Robertsonian translocation between chromosomes 13 and 21 (2 cells of 20; Supplementary Fig. S3B).
Basic characterization of iPSC clones including hESC-like morphology, passaging >10 passages, pluripotency marker analysis, transgene silencing, and capability to differentiate into all 3 germ layers (embryoid body assay).
Ter., teratoma; Kar., karyotype; iPSC, induced pluripotent stem cell; hESC, human embryonic stem cell; SeV, Sendai-virus.
All iPSC lines were cultured for at least 10 passages. During that time, the cells exhibited growth properties and morphology similar to that of hESC, characterized by high nucleus to cytoplasm ratio, large nucleoli, and formation of tightly packed and flat colonies. All iPSC lines expressed cell surface markers typical for hESC colonies and had a nuclear localization of a transcription factor OCT4 and NANOG (Figs. 3E and 4A).
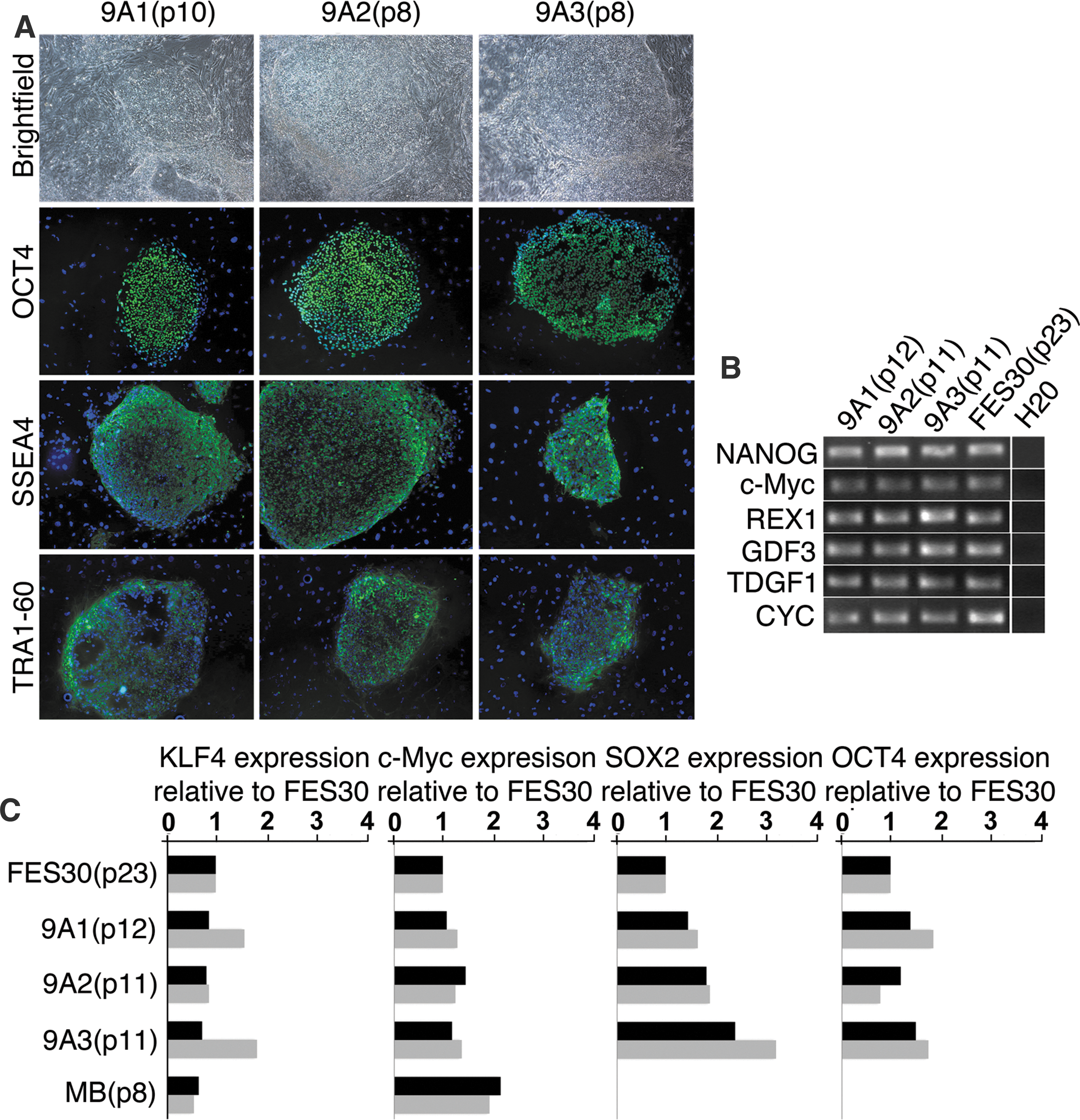
Characterization of pluripotency and transgene silencing in retrovirally derived iPSC.
q-PCR showed that all of the iPSC lines expressed undifferentiated hESC marker genes OCT4, SOX2, c-Myc, and KLF4 and had silenced their transgenes (Fig. 4C). Also, other genes typically characterizing human pluripotent stem cells, including REX1, TDGF1, GDF3, OCT4, and NANOG, were expressed in all iPSC lines derived using SeV or retroviruses vectors (Figs. 3D and 4B). At passage 10, SeV-derived iPSC colonies were integration- and transgene free (Fig. 3D).
Evaluation of pluripotency of iPSCs
The pluripotent differentiation potential of the skeletal myoblast-derived iPSCs was first analyzed by in vitro EB differentiation assay. Round cell aggregates were apparent after 4–6 days in suspension culture (Figs. 3F and 5A and data not shown) The EBs were maintained in cell suspension for additional 8 days. Cells were positive for all 3 germ layers, Vimentin (mesoderm and parietal endoderm), βIII-tubulin (ectoderm), Brachyury (mesoderm), and FOXA2 (endoderm) were found in sectioned EBs (Figs. 3F and 5A, and data not shown).
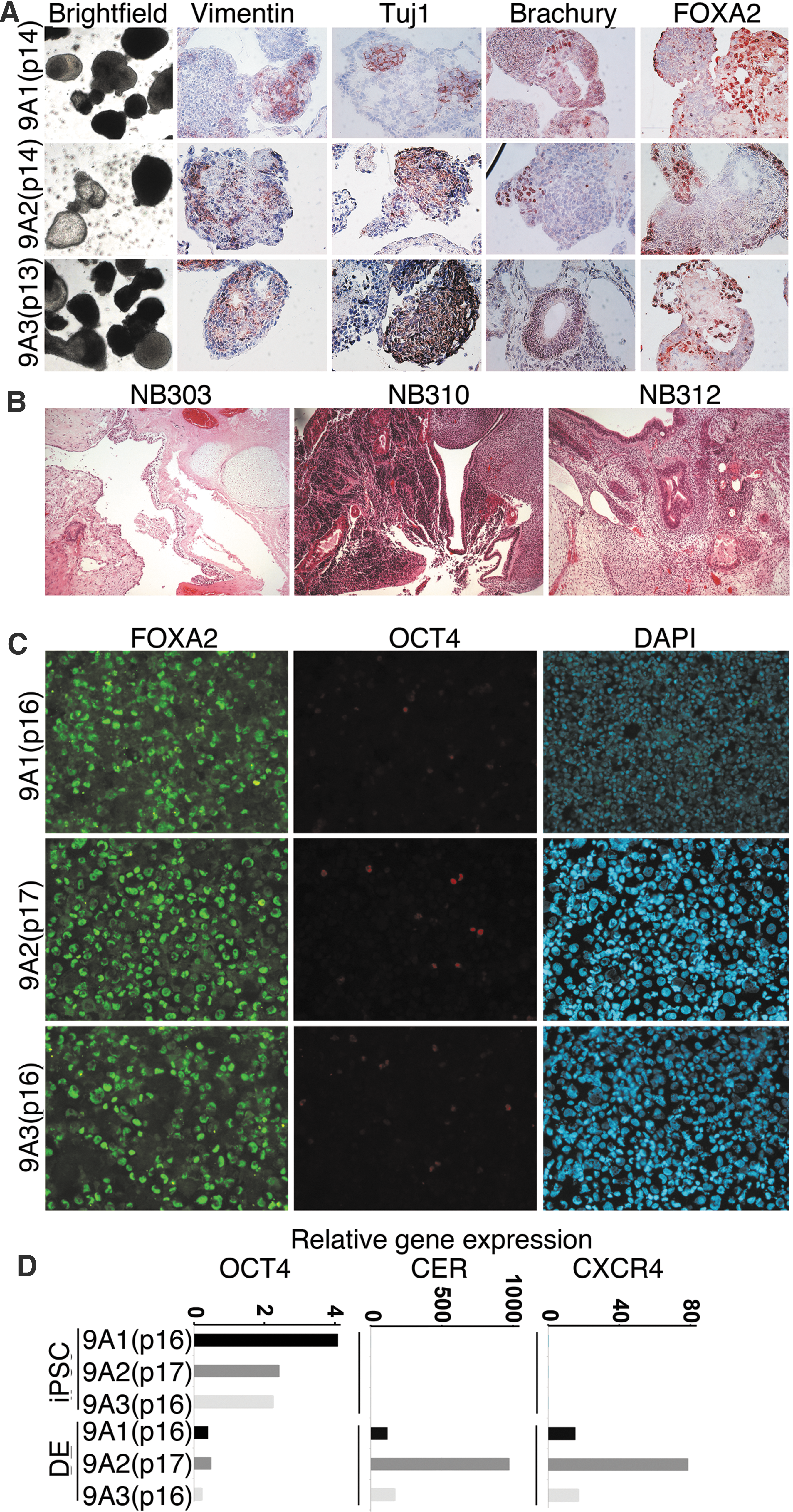
Differentiation of retrovirally derived iPSC.
For teratoma formation, iPSC lines 9A1 (passage 19), 9A2 (passage 18), and 9A3 (passage 17) were injected into the testis of male immune-deficient NMRI nu/nu mice. After 8 weeks teratomas were harvested. Histological examination showed that the tumors contained multiple differentiated tissues, derived from all germ layers (Fig. 5B).
The iPSC lines 9A1 (passage 16 and 18), 9A2 (passage 17 and 28), and 9A3 (passage 16) were tested for their capacity to differentiate into definitive endoderm. At the end of the differentiation protocol, all 3 lines were shown to express definitive endoderm specific markers FOXA2, CER, and CXCR4 (Fig. 5C, D). FACS analysis at day 7 of definitive endoderm differentiation showed that all iPSC lines expressed at least 60% of definitive endoderm marker CXCR4 (data not shown).
Discussion
We have reported 3 major findings. First, our study demonstrates efficient iPSC generation from human skeletal muscle biopsy-enriched myoblasts using integrative or integration-free methods with the same factors (Oct4, Sox2, Klf4, and c-Myc) that have been used to reprogram fibroblasts. The myoblast-derived iPSCs display characteristics similar to hESC, express key pluripotency markers, differentiate into cell types of the 3 germ layers, and can be target differentiated into definitive endoderm. This adds primary myoblasts to the growing list of human somatic cell types that are amenable for pluripotent reprogramming [8,10,28,29]. Although c-Myc provides an advantage in enhancing reprogramming success, it is a potent oncogen and its reactivation has been shown to increase tumorigenicity in chimeric mice and their progeny [30]. Replacing c-Myc with related proteins or small molecules would likely be required for clinical application of myoblast-iPSCs. In addition, the use of nonintegrating reprogramming methods eliminates the risk of transgene reactivation.
Second, we found that the efficiency of iPSC induction using Oct4, Sox2, Klf4, and c-Myc, from human myoblast cells was similar to that of fibroblasts. However, as mouse myoblasts have previously been reported to reprogram with very poor efficiency [31,32], it was surprising to notice no significant difference in reprogramming efficiencies between human myoblasts and fibroblasts. The media composition seems to play an important role in this context, as changing the reprogramming medium from fibroblast to myoblast culture medium for the first days of reprogramming reduced the amount of reprogrammed colonies. This implies that the myoblast culture medium may have an inhibitory effect on the reprogramming efficiency of human myoblasts by some yet undefined mechanism. It has been shown that both MyoD and Oct4 can direct the binding of Smad3, downstream factor in TFG-beta signaling, to different genomic sites [33], and this kind of effect could potentially also happen with other signaling factors that contribute to the maintenance of myoblast cell state. It is thus possible that the signaling provided by the myoblast culture medium, which maintains the myoblast phenotype, may be guided by myogenic transcription factors to provide a nonpermissive state for reprogramming, and that the short time in nonmyoblast medium could provide a window for the transgenic reprogramming factors to initiate proper reprogramming process. Interestingly, the effect of media change on colony-formation efficiency seems to also happen in a fairly similar time scale as the epigenetic changes in the MyoD promoter in fibroblasts mediated by overexpression of either MyoD or Oct4 [34].
Third, we found that the addition of HDAC inhibitor (NaB) significantly enhanced the efficiency of iPSC generation from myoblasts to similar extent as from the fibroblasts in feeder-free conditions. In addition to butyrate, the ALK4/5/7 inhibitor (SB431542) had a positive effect on the efficiency of iPSC reprogramming. Previously, it has been shown that small molecule inhibitors enhance the reprogramming efficiency of human fibroblasts, adipose tissue-derived cells, and cord blood cells [22,35]. The use of small molecules is an attractive way to scale up efficacy of iPSC generation and can be a valuable tool for production of iPSCs for biobanks. However, it is important to study what the impact of this is on the genetic integrity of iPSCs, since the standard methods are known to lead to genomic instability [25].
The possibility to purify human myoblast cells and their capability to proliferate and fuse into multinucleated fibers in tissue culture offers an experimental model to study mechanisms of muscle disorders. However, pluripotent reprogramming provides the possibility to generate any other cell types from long-term frozen myoblasts, which often may represent the only available cell type from a specific patient. Previous reports have shown that iPSCs can retain epigenetic memory and differentiate preferentially along the origins of a parental cell line [31,36,37]. It is possible that myoblast-derived iPSC lines have increased ability to differentiate toward muscle lineages. Generation of myoblast-iPSCs provides valuable tools for the understanding of disease pathogenesis and for the identification of novel therapeutic approaches.
Conclusions
We have successfully derived iPSCs from enriched human myoblasts using integrating and integration-free methods. Myoblasts are reprogrammed with the same efficiency as fibroblast cells when appropriate media is used. Addition of small molecule inhibitors, HDAC inhibitor (NaB) and ALK4/5/7 inhibitor (SB431542) to the cocktail significantly enhances the efficiency of retroviral myoblast reprogramming in a feeder-free condition.
Footnotes
Acknowledgments
We thank Jarkko Ustinov, Jaan Palgi, and Cia Olsson for skilled assistance in characterization of human pluripotent stem cells, as well as Johanna Pispa and Christian Unger for critical comments on the article. This study was supported by grants from The Academy of Finland, Sigrid Jusélius Foundation, Helsinki University Hospital Research Funds (for T.O. and A.S.), Biocenter Finland (for T.O.), Jane and Aatos Erkko Foundation, and the European Research Council (for A.S.)
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.