
Abstract
Although transplanted pluripotent stem cell-derived neurons can contribute to functional recovery in animal models of Parkinson's disease, the risk of tumor formation hinders clinical applications of this approach. Removing undifferentiated cells from the donor population is critical to reduce tumorigenesis. Moreover, immature neural progenitors in transplants can proliferate unpredictably, resulting in neural overgrowth and long-term risks of compressing the surrounding host tissue. Because Notch signaling plays a role in maintaining the multipotency and proliferative capacity of neural progenitors, we used γ-secretase inhibitors (GSIs) to dampen Notch signaling in human-induced pluripotent stem cell-derived neural progenitors before transplantation and examined the effects on the growth of proliferative grafts. We observed a marked reduction in the percentage of dividing cells and increased neuronal maturation in GSI-treated samples in vitro. Next, grafts were transplanted into the striata of nonobese diabetic/severe combined immune deficiency mice. Histological analyses performed 8 weeks after the operation showed that grafts pretreated with GSIs—N-[N-(3,5-difluorophenacetyl)-
Introduction
C
Recent efforts have focused on removing undifferentiated pluripotent cells from the donor population to avoid teratoma formation. Grafts containing proliferating neural progenitor cells (NPCs), however, may continuously grow instead of terminally differentiating into postmitotic neurons; this unpredictable graft growth—referred to as neural overgrowth—may adversely affect surrounding host tissue [13]. To use stem cell replacement therapy clinically, multidisciplinary strategies are needed to ensure patient safety and optimize outcomes. In this study, we examine the effects of small molecule γ-secretase inhibitors (GSIs) on graft growth after transplantation.
Recent studies have revealed remarkably diverse roles of Notch-related signaling during various aspects of embryonic development. During in vivo neural development, Notch first controls the induction of definitive neural stem cells, and then plays an important role in the maintenance of the cells' undifferentiated and highly proliferative state via such downstream effector genes as Hes1 and Hes5 [14
–17]. Hatakeyama et al. identified extensive defects in brain formation in Hes1/Hes5 double-mutant mouse embryos after E8.5 owing to insufficient expansion of NPCs and premature neuronal differentiation [18]. Indeed, when a sufficient number of NPCs have developed, inactivation of Notch signaling in NPCs drives neuronal differentiation via the expression of such proneural genes as Ngn2 and Mash1, expression of which is repressed by Hes1 and Hes5 when Notch signaling is active [16,17,19]. Similar molecular processes have been observed during in vitro neuronal differentiation of NPCs when Notch signaling was blocked with the small molecule GSI N-[N-(3,5-difluorophenacetyl)-
Because inhibiting Notch signaling induces proliferative NPCs to develop into a more mature state with limited ability to proliferate, we hypothesized that treating NPCs with GSIs before transplantation may prevent graft overgrowth in vivo by reducing overall proliferation. In this study, we first confirmed the in vitro effects of GSIs on the proliferation and differentiation of NPCs derived from human iPSCs. We then evaluated whether transient treatment with GSIs before transplantation reduced the growth and/or differentiation of NPC grafts in vivo.
Materials and Methods
Cell culture
Two lines of human iPSCs were used for in vitro studies; 253G-4 cells were generated following retroviral transfection of 4 genes (OCT3/4, SOX2, KLF4, and c-MYC), whereas 404C-2 cells were reprogrammed with episomal plasmid vectors containing 5 genes (OCT3/4, SOX2, KLF4, LIN28, and L-MYC) and sequence encoding p53-specific shRNA [21,22]. Cells were maintained on mitomycin C-inactivated mouse STO feeder cells in the Dulbecco's modified Eagle's medium (DMEM)/F-12 medium (Wako) supplemented with 20% knockout serum replacement (KSR; Invitrogen), 2 mM
Derivation of dopaminergic progenitors from human iPSCs
To induce dopaminergic neurons, we used the previously described serum-free culture of embryoid body-like aggregates quick method with some modifications (Fig. 1) [23]. Briefly, human iPSCs were pretreated with 10 μM Y27632 (ROCK inhibitor; Wako) overnight to prevent apoptosis [24], dissociated into single cells using Accumax (Innovate Cell Technologies), and seeded onto 96-well low-adhesion plates (Lipidure-Coat Plate A-U96; NOF Corporation) at a density of 9,000 cells/well in DMEM/F-12 supplemented with 5% KSR, 2 mM
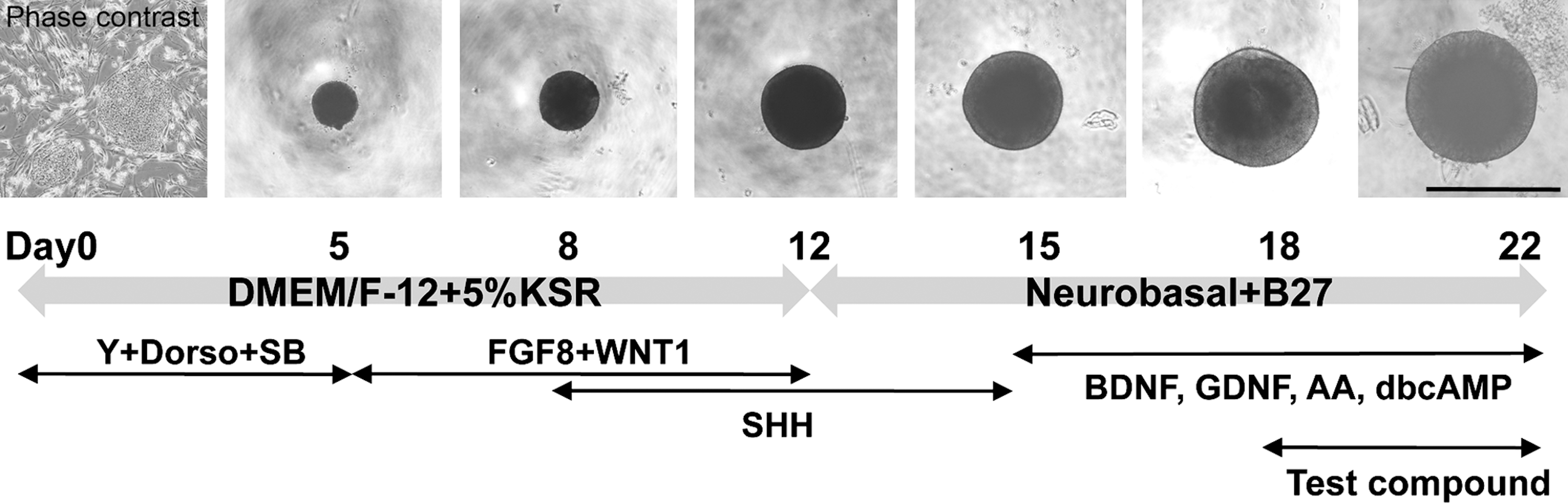
Inducing dopaminergic progenitor development from human iPSCs in vitro. Pluripotent human iPSCs were dissociated and seeded onto U-bottomed 96-well plates to induce rapid reaggregation on day 0. The aggregated cells were induced to differentiate into dopaminergic neurons using various factors during the 22-day incubation period. Test compounds, including GSIs, were added to the culture during the last 4 days. Scale bar 1 mm. Dorso, dorsomorphin; SB, SB431542; Y, Y27632; iPSCs, induced pluripotent stem cells; GSI, γ-secretase inhibitor.
Treatment of neuronal progenitors with small molecule GSIs and δ-opioid peptide
DAPT (Sigma) and compound E (Santa Cruz Biotechnology) are potent nontransition-state analogue inhibitors of γ-secretase that are thought to interact with the same active site between presenilin-1 heterodimers in γ-secretase complexes [26]. The agents were dissolved in dimethylsulfoxide (DMSO), and used at a final concentration of 10 μM, the highest concentration at which they effectively inhibited division of NPCs without precipitating in culture or producing toxic effects in preliminary experiments. The vehicle control was 0.1% DMSO. Differentiated dopaminergic progenitors on day 18 were treated with GSIs for 4 days before transplantation on day 22. The δ-opioid peptide [D-Ala(2), D-Leu(5)]-enkephalin (DADLE) was also used for flow cytometric analysis of the cell cycle because the peptide potently inhibits the growth of NPCs of mesencephalic origin [27]. DADLE was used alone at a final concentration of 1 or 10 nM, and at 1 nM in combination with the opioid antagonist naltrexone (0.1 nM).
Quantification of neurite extension
Sterilized axon investigation system (AXIS) devices (Millipore) were placed onto tissue culture dishes precoated with poly-
Transplantation
Animals were cared for and handled according to the Guidelines for Animal Experiments of Kyoto University and the National Institute of Health Guide for the Care and Use of Laboratory Animals from the Institute of Laboratory Animal Resources. For in vivo studies, a donor population was prepared from the 253G-4 human iPSC line. Male nonobese diabetic/severe combined immune deficiency mice (5–6-weeks old; CLEA Japan) were anesthetized and injected stereotactically with day-22 NPCs in the midstriatum (from the bregma: A+1.0 mm; L+1.8 mm; V+2.5 mm). Suspension cultures of NPCs were treated overnight with 10 μM Y27632 to minimize the damage caused by transplantation, and spheres were transplanted using 22-gauge needles without enzymatic dissociation. Eight weeks after transplantation, animals were deeply anesthetized and intracardially perfused with 4% paraformaldehyde.
Flow cytometry
Following overnight treatment with 10 μM Y27632, day-22 cultures pretreated with GSIs, DADLE, or DADLE and naltrexone were incubated with 10 μM bromodeoxyuridine (BrdU; BD Pharmingen) for 3 h at 37°C. Subsequently, cells were dissociated in a 5-min incubation with Accumax, fixed, permeabilized, and stained with fluorochrome-conjugated anti-BrdU antibodies and 7AAD using APC BrdU Flow Kits (BD Biosciences). Flow cytometry and data analysis were performed on an LSR1 system (BD Biosciences).
Immunofluorescence and terminal deoxynucleotidyl transferase dUTP nick end labeling
For in vitro experiments, GSI-treated spheres at day 22 were fixed in 4% paraformaldehyde, frozen, and sliced into 10-μm-thick slices with a microtome for immunocytochemistry. Following in vivo experiments, brains were removed and refixed overnight in 4% paraformaldehyde, cryopreserved in 30% sucrose for 2 days, frozen, and sliced into 40-μm-thick slices with a microtome for immunohistochemistry. Frozen sections of the spheres and brains were permeabilized and blocked for 1 h at room temperature with 0.3% Triton X-100 and 4% donkey serum in PBS, followed by overnight incubation with primary antibodies at 4°C. Primary antibodies used in the study included mouse anti-Ki67 (1,000 ×; Novocastra), mouse anti-MAP2ab (500×; Sigma), rabbit anti-PAX6 (200×; Covance), mouse anti-Tubβ3 (1,000×; Covance), rabbit anti-TH (500×; Millipore), and mouse anti-human nuclear antigen (HNA, 400×; Millipore). After 3 washes with 0.05% Tween-20 in PBS, samples were incubated for 1 h with Alexa Fluor-conjugated secondary antibodies at room temperature. Following 3 additional washes with 0.05% Tween-20 in PBS, samples were incubated with 4′,6-diamidino-2-phenylindole for nuclear staining and mounted with Permafluor (Dako).
Detection of apoptosis in GSI-treated spheres at day 22 was combined with immunocytochemistry for Ki67, and performed using an In Situ Cell Death Detection Kit with tetramethylrhodamine red (Roche). Briefly, immunostained samples were incubated with a mixture of terminal deoxynucleotidyl transferase enzyme and nucleotides for 1 h at 37°C before mounting. Only frozen sections spanning the center of the spheres were used for imaging and subsequent analysis. Fluorescent images were obtained with a FV1000D laser-scanning confocal microscope (Olympus) and a BioRevo fluorescence microscope (Keyence). Ki67+ cells and TUNEL+ cells were quantified by averaging the percentages of labeled cells in 3 different spheres using a TissueFAXS cell analysis system (Novel Science). Percentages of PAX6+, MAP2ab+, TH+, and Tubβ3+cells were manually counted.
For evaluations of in vivo results, only grafts that showed good cell viability (HNA+ cells >500) were quantitatively analyzed. Graft volume for each animal was determined by identifying HNA+ areas in every third 40-μm-thick section using BZ-II Analyzer software (Keyence), and totaling the volumes of all 0.12-mm-tall cylinders according to Cavalieri's principle. PAX6+, Ki67+, and Tubβ3+ cells in the grafts were manually counted in 3 randomly selected fields of view, each containing more than 500 HNA+ cells, where possible, and the average percentages of positive cells were calculated for each animal. TH+ neurons were counted manually in every third section, and the total number in each graft was estimated using the Abercrombie correction [28].
Quantitative reverse transcription-polymerase chain reactions
Total RNA was extracted from iPSC-derived NPCs using an RNeasy Plus Mini kit (Qiagen), and reverse transcribed with the Super Script III First-Strand Synthesis System (Invitrogen). Quantitative polymerase chain reactions (PCRs) were performed using Power Sybr (Applied Biosystems) according to the manufacturer's instructions. The expression levels of each gene were normalized to that of GAPDH using the ΔCT method. Primers used for the reactions are shown in Table 1.
Statistical analysis
Statistical analyses were performed using GraphPad Prism software Ver. 5.0b (GraphPad Software). More than 30 linear neurites that were TH+ in the channels were measured for each group to examine axon lengths, and one-way analysis of variance (ANOVA) and Kruskal–Wallis tests were performed to statistically analyze the results (Fig. 3I). Quantitative data obtained using flow cytometry, immunofluorescence, and terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL), quantitative PCRs, and immunohistochemistry were statistically analyzed using one-way ANOVA and Newman–Keuls post-tests (Figs. 2B, D, 3A, C, D, F, G, 4B, and 5B, D, F, H, I). Differences were considered statistically significant when P<0.05. All quantitative data are presented mean values±standard error of the mean. All in vitro data were obtained from at least 3 independent experiments.
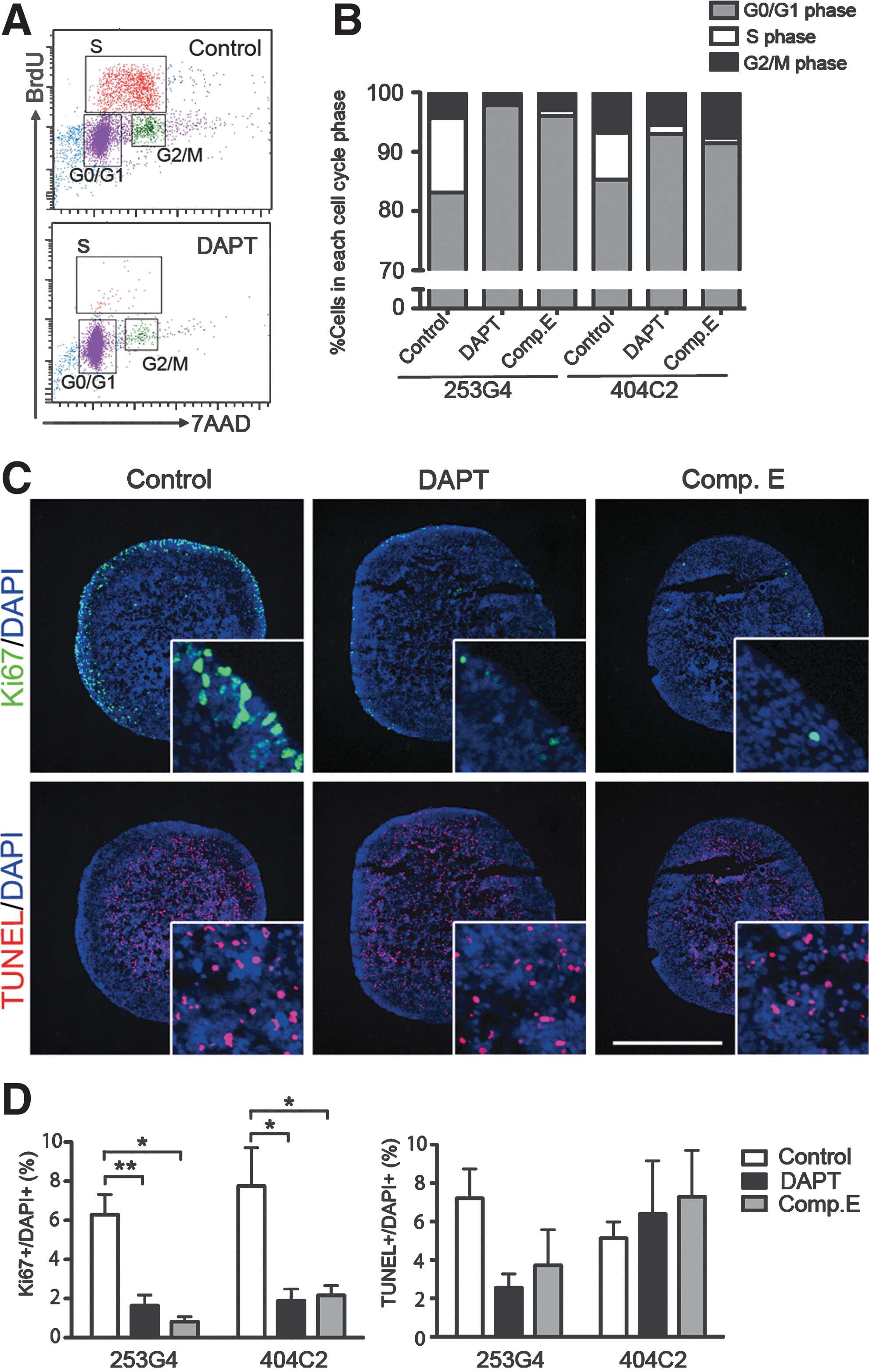
GSIs significantly reduced cell proliferation.

Characterization of cells treated with GSIs.
Results
GSIs reduced the proliferation of human iPSC-derived NPCs
We first determined whether substances that have been previously reported to slow cell-cycle progression in NPCs exerted similar effects on the heterogeneous population of human iPSC-derived NPCs [20,27]. Two human iPSC lines—253G-4 and 404C-2 cells—were induced to differentiate into dopaminergic neurons using a floating culture system and a combination of growth factors (Fig. 1).
To analyze the effects of GSIs, we first determined the optimal timing to inhibit Notch signaling in our system. Because Notch signaling is required for pluripotent stem cells to assume a neural fate [29], subject population would ideally contain NPCs that are free of pluripotent cells, but still able to proliferate quickly and, most importantly, Notch signaling should be active in the cells. Quantitative reverse transcription-polymerase chain reaction (RT-PCR) analysis revealed that the expression of the pluripotency marker, OCT3/4, was downregulated by day 8 (Supplementary Fig. S1; Supplementary Data are available online at
Following 4 days of treatment with DMSO (control) or test compounds, day-22 cultures were subjected to BrdU uptake assays and flow cytometry to assess the various phases of the cell cycle. In control cultures containing 253G-4 cells, 83.1%±2.1% of the viable cells were in the G0/G1 phase, whereas 12.6%±2.0% and 4.3%±0.3% were in the S and G2/M phases, respectively, indicating that the cells were normally progressing through the cell cycle (Fig. 2A, B). Inhibiting Notch signaling with DAPT significantly decreased the percentage of cells in the S phase (0.37%±0.11%) and increased the percentage of cells in the G0/G1 phase (97.9%±0.4%) (n=3). Similar effects were observed with the compound E, and with the 404C-2 cell line (n=3 for all conditions). DADLE alone or in combination with naltrexone, however, failed to slow cell-cycle progression in our protocol (Supplementary Fig. S2). Therefore, we used the GSIs DAPT and compound E for the subsequent analyses.
Next, we assessed the effects of GSIs on the proliferation and viability of iPSC-derived populations containing NPCs. Immunofluorescent staining of cell aggregates revealed the presence of Ki67+ proliferating cells in the outer layer, and TUNEL+ apoptotic cells in the sample centers (Fig. 2C). DAPT and compound E significantly reduced the percentages of Ki67+ cells in the aggregates of 253G-4 cells (control, 6.3%±1.0%; DAPT, 1.6%±0.5%; compound E, 0.8%±0.3%), whereas the percentage of TUNEL+ cells was unaffected at approximately 5% in all samples, indicating that the GSIs suppressed proliferation without causing apoptosis (Fig. 2D).
GSI treatment accelerated neuronal maturation of NPCs in vitro
Previous studies demonstrated that inhibiting Notch signaling promotes neuronal differentiation from NPCs [13,20,30]. To determine if the reduced proliferation of GSI-treated NPCs was accompanied by increased differentiation into neuronal-lineage cells, we analyzed the expression of neural markers in day-22 samples. Because the 253G-4 and 404C-2 cell lines produced similar results in the experiments described above, we used the 253G-4 cell line for the following analyses. Quantitative RT-PCRs showed that the expression of HES5 was almost abolished by the GSIs, indicating that Notch signaling was efficiently inhibited (Fig. 3A). Expression of early neural markers, such as SOX1 and PAX6, were also downregulated, whereas expression levels of the mature neuronal marker, MAP2ab, and the dopaminergic neuron marker, TH, were upregulated by the GSIs (Fig. 3A). These effects were confirmed in immunofluorescence analyses. The outer layer of control spheres was densely packed with PAX6+ NPCs, which were much less prominent in spheres treated with DAPT or compound E (Fig. 3B, C). Because this layer also contained Ki67+ cells (Fig. 2C), the populations were proliferating NPCs in day-22 control spheres. In contrast, the percentage of MAP2ab+ postmitotic neurons significantly increased in spheres treated with DAPT or compound E (control, 3.3%±1.9%; DAPT, 57.7%±8.3%; compound E, 72.7%±5.6%) (Fig. 3B, D). Similarly, GSI treatment significantly increased the percentage of cells positive for the neuronal marker, Tubβ3, indicating that GSIs promoted neuronal maturation of human iPSC-derived NPCs in day-22 cultures (Fig. 3E, F). Among the Tubβ3+ population, some TH+ dopaminergic neurons were observed on day 22 (Fig. 3E). More TH+ neurons were detected in GSI-treated spheres, although the percentages of TH+ cells among Tubβ3+ neurons were similar under all culture conditions (control, 9.0%±3.3%; DAPT, 12.8%±4.0%; compound E, 13.1%±2.8%; Fig. 3G), implying that GSI treatment promoted neuronal maturation of NPCs without affecting the development of specific neuronal subtypes.
To further assess neuronal maturation, we analyzed neurite lengths in neurons using a culture chamber with 900-μm-long slits (Fig. 3H). Day-22 samples pretreated with GSIs or DMSO for 4 days were dissociated and seeded onto the chambers. Three days later, the cells were subjected to immunofluorescence staining for Tubβ3 and TH. In all samples, most Tubβ3+ neurites in the slits were also positive for TH (Fig. 3H). On average, TH+ neurites extending from control cells were 203.2±24.2 μm in length, while neurites extending from those treated with DAPT or compound E were significantly longer (384.4±37.2 μm or 496.3±48.5 μm, respectively; Fig. 3H, I).
Pretreating NPCs with GSIs reduced the graft size
Our in vitro results suggested that pretreating NPCs with GSIs may reduce the growth of the grafts by inhibiting cell proliferation. To confirm this hypothesis, we treated human NPCs with GSIs for 4 days, and grafted the resulting samples (2×105 cells in 1 μL) into the striata of nonobese diabetic/severe combined immune deficiency mice. At 8 weeks post-transplantation, brain slices were subjected to immunofluorescence staining for HNA. Cells that were not pretreated with GSIs extensively proliferated in the grafts to form numerous neural rosette-like structures of variable size (1.00±0.13 mm3; Fig. 4A and Table 2). In contrast, transplants containing DAPT- or compound E-treated cells resulted in smaller grafts with no sign of active proliferation (0.14±0.05 mm3 or 0.06±0.05 mm3, respectively). The grafts were rod-shaped with no or only a few rosette-like structures. The grafted cells did not survive well in some of the animals (<500 cells in 3/7 and 1/7 DAPT- and compound E-treated recipients, respectively; Table 2). These animals were excluded from further analyses. The volumes of the grafts derived from GSI-treated cells were significantly smaller than those of grafts derived from control spheres (Fig. 4B).

Histologic analyses of human iPSCs derived grafts with or without GSI pretreatment.
HNA, human nuclear antigen; DAPT, N-[N-(3,5-difluorophenacetyl)-
Grafts containing GSI-treated cells contained a more mature neuronal population
Based on our in vitro results showing that GSIs promoted the neuronal maturation of NPCs, we compared neural phenotypes in grafts obtained using iPSC-derived samples with or without GSI pretreatment. In control grafts, 28.9%±4.8% of HNA+ grafted cells were also positive for the early NPC marker PAX6; the double-positive cells were frequently located in and nearby neural rosette-like structures (Fig. 5A, B). In contrast, significantly fewer PAX6+ cells were identified in grafts containing DAPT- or compound E-treated cells (9.2%±1.2% or 8.2%±2.6%, respectively). In control grafts, 15.7%±2.2% of cells were Ki67+, with many positive cells located at the center of the rosettes, whereas the percentages of Ki67+ cells in the grafts containing GSI-treated cells were significantly lower (Fig. 5C, D). In contrast, examination of postmitotic neurons revealed that 72.0%±16.0% and 82.0%±4.4% of cells in the grafts containing DAPT- and compound E-treated cells were Tubβ3+, respectively, percentages that were significantly higher than that observed in control grafts (17.0%±8.0%; Fig. 5E, F). For dopaminergic neurons, TH+ cells were found in most viable grafts except for one in the control group, although the estimated number of TH+ cells varied between 0 and 216 cells in the different animals (Fig. 5G, H). On average, the percentages of TH+ cells were higher in grafts containing GSI-treated cells (DAPT, 0.9%±0.5%; compound E, 1.6%±0.8%); however, no significant differences were noted in comparisons with control grafts (0.01%±0.006%; Fig. 5I). These observations suggested that control grafts contained immature and proliferating NPCs, whereas grafts pretreated with GSIs contained a postmitotic neuronal population. These data indicate that pretreating iPSC-derived NPCs with GSIs promoted neuronal maturation in vivo, and that the difference in the compositions of the grafts contributed to the difference in graft volumes.

Characterization of viable grafts pretreated with GSIs. Graft cells positive for PAX6, Ki67, Tubβ3, and TH. Control grafts contained numerous PAX6+and Ki67+ NPCs, especially near neural rosettes
Discussion
In this study, we examined the effects of GSIs on the proliferation and neuronal differentiation of human iPSC-derived NPCs. Treating day-22 samples containing proliferative NPCs for 4 days with GSIs reduced the percentage of dividing cells in vitro. Based on changes observed in neural marker expression, gene expression profiles, and neurite outgrowth, the GSIs likely promoted the transition of immature progenitors into committed neuronal precursors. We also demonstrated that pretreating donor cells with GSIs significantly reduced graft volume 8 weeks after transplantation, and that pretreated grafts contained more mature neurons compared with control samples, indicating that the GSI treatment produced similar results in vivo.
Previous studies have highlighted the critical role of Notch signaling in the maintenance of progenitor status for NPCs in vitro [13,20] and in vivo during development [15 –18]. Our in vitro experiments confirmed that inhibiting Notch signaling promoted neuronal differentiation of NPCs derived from 2 lines of human iPSCs. The effects of the GSIs were more prominent than those reported in a previous study by Borghese et al. [20]. The difference in these data may have resulted from the use of FGF2 and epidermal growth factor in the previous study to expand the NPC population, which has been shown to maintain NPCs in an undifferentiated state [31].
The duration of GSI treatment is an important parameter to consider. In the developing neural tube, expression levels of genes downstream of Notch signaling, including Hes1, Ngn2, and Dll1, have been shown to oscillate in individual NPCs such that neighboring NPCs activate each other to maintain a constant level of Notch signaling in the pool of proliferating NPCs [32]. After a period of proliferation, sustained repression of Hes gene expression causes sustained expression of Ngn2 and Dll1, leading to the initiation of neurogenesis. Using E4.5 embryonic chick retinal explants, Nelson et al. showed that NPCs should be treated with DAPT for at least 6 h to commit the cells to neuronal differentiation, even though Hes5 expression was reduced after 3 h [30]. In our preliminary transplantation experiment, pretreating day-18 population with compound E for 3 days or less only slightly reduced the sizes of resulting grafts, and neural rosettes still formed (data not shown). This result indicated that insufficient inhibition of Notch signaling may leave NPCs in an undifferentiated state that would allow continued proliferation after transplantation. Treating the donor cells with a GSI for 4 days markedly reduced the number of proliferative cells in the donor population, although cell viability was also reduced in some cases. Techniques to modulate the host brain environment to support mature neurons should be investigated to improve the survival of vulnerable transplants.
Alternatively, delivering GSIs in vivo after immature NPCs are transplanted may result in better cell survival in the grafts. When using this iPSC technology clinically, however, systemic administration of GSIs may be problematic because Notch inhibition in vivo is associated with a wide range of serious adverse effects, including diarrhea, altered lymphocyte development, and thymus atrophy, to name a few [33,34]. Moreover, a recent large clinical trial of the GSI semagacestat for Alzheimer's disease showed an increased risk of skin cancer in those treated with the drug [35].
Other than controlling graft growth, treating the donor population with GSIs resulted in more mature and homogenous grafts compared with control grafts, which were large with numerous neural rosettes. During embryonic neurogenesis, lateral inhibition mediated by Notch signaling is crucial for preventing NPCs from undergoing synchronized neuronal differentiation and reserving an undifferentiated population to generate glia and more neurons. This type of local cell −cell signaling, however, is problematic if the goal is to obtain a homogenous population of neurons in vitro. To commit all NPCs to neurons at once, it would be beneficial to inhibit Notch signaling simultaneously in the population rather than allowing the signaling to wane during the course of differentiation. Indeed, pretreating the donor population with GSIs provided the advantage of achieving better homogeneity in the grafts.
Although GSI-treated grafts yielded a higher number of TH+ neurons compared with control samples, the percentage was not satisfactory considering the frequency obtained in vitro. It is possible that terminally differentiated TH+ dopaminergic neurons would be too fragile to survive the transplantation procedure. Instead, dopaminergic precursor cells that are just becoming or just have become postmitotic may survive and terminally differentiate into TH+ neurons in the host brain. We believe that transiently treating the donor population with GSIs may have resulted in this type of population. To improve the yield of TH+ neurons after transplantation, strategies should be sought to modify the host brain environment to support the terminal differentiation of grafted dopaminergic progenitors.
In conclusion, treating iPSC-derived NPCs with GSIs before transplantation can control the growth of a potentially proliferative cell population in vivo. Other strategies to prevent overgrowth of grafted stem cells include prolonged differentiation, selection of more mature cells using fluorescence-activated or magnetic-activated cell sorting, introduction of suicide genes, and irradiation [36]; these approaches, however, require complicated procedures or long periods to prepare donor cells. Compared with these techniques, GSI treatment is a simple and inexpensive procedure that can be used in combination with other promising options, such as cell sorting, to optimize the safety of cell replacement therapies using iPSCs.
Footnotes
Acknowledgments
We thank Dr. M. Nakagawa and Dr. K. Okita for providing human iPSCs. We also thank Mr. K. Kubota for technical assistance and other members of our laboratory. This study was supported by a grant to the Kobe Cluster from the Ministry of Education, Culture, Sports, Science and Technology (MEXT) in Japan.
Author Disclosure Statement
The authors have no conflicts of interest to report.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.