
Abstract
We sought to determine whether neural stem cells (NSCs) can be isolated from the amniotic fluid in the setting of neural tube defects (NTDs), as a prerequisite for eventual autologous perinatal therapies. Pregnant Sprague-Dawley dams (n=62) were divided into experimental (n=42) and control (n=20) groups, depending on prenatal exposure to retinoic acid for the induction of fetal NTDs. Animals were killed before term for analysis (n=685 fetuses). Amniotic fluid samples from both groups underwent epigenetic selection for NSCs, followed by exposure to neural differentiation media. Representative cell samples underwent multiple morphological and phenotypical analyses at different time points. No control fetus (n=267) had any structural abnormality, whereas at least one type of NTD developed in 52% (217/418) of the experimental fetuses (namely, isolated spina bifida, n=144; isolated exencephaly, n=24; or a combination of the two, n=49). Only amniotic samples from fetuses with a NTD yielded cells with typical neural progenitor morphology and robust expression of both Nestin and Sox-2, primary markers of NSCs. These cells responded to differentiation media by displaying typical morphological changes, along with expression of beta-tubulin III, glial fibrillary acidic protein, and/or O4, markers for immature neurons, astrocytes, and oligodendrocytes, respectively. This was concurrent with downregulation of Nestin and Sox-2. We conclude that the amniotic fluid can harbor disease-specific stem cells, for example, NSCs in the setting of experimental NTDs. The amniotic fluid may be a practical source of autologous NSCs applicable to novel forms of therapies for spina bifida.
Introduction
N
Spina bifida leads to significant spinal cord damage at and below the lesion and is often accompanied by hydrocephalus. Common manifestations include paraplegia, sensory loss, bowel, bladder, and sexual dysfunction, and secondary musculoskeletal deformities [6,7]. It is among the most morbid survivable congenital anomalies; so much so, that it has prompted the first ever prospective randomized control trial of prenatal surgical repair of a nonlife threatening anomaly, widely known as the Management of Myelomeningocele Study (MOMS) [8]. This sentinel undertaking has shown that, compared with the conventional postnatal coverage of the lesion, prenatal repair does offer some benefits, but to a limited extent and only in a select subset of fetuses, not without notable maternal risks [8].
Using a large animal model, we have previously introduced the notion of therapeutic NSC delivery to the spinal cord in fetuses with spina bifida, as a potential means of promoting actual neural tissue repair, rather than solely preventing further damage by simple coverage, as originally aimed in the MOMS [9]. In that initial study, donor NSCs were necessarily of a xenologous origin. Certainly, for the eventual clinical translation of this therapeutic concept to be rendered viable, an autologous source of NSCs would be preferred. Neural cells have long been known to be present in the amniotic fluid in the setting of NTDs [10 –13]. In this study, we aimed at testing our hypothesis that a population of NSCs is also present therein in this setting. This would be a prerequisite for the development of minimally invasive autologous perinatal NSC-based strategies for the treatment of these anomalies.
Materials and Methods
This study was approved by the Children's Hospital Boston's Institutional Animal Care and Use Committee under protocol # 09-05-1353.
Sixty-two time-dated pregnant Sprague-Dawley dams (Charles River Laboratories, Inc., Wilmington, MA) were housed under a standard dark/light cycle and fed a normal diet ad libitum. Animals were divided into 2 groups. One group, control (n=20), underwent no manipulations. The other group, experimental (n=42), was exposed to retinoic acid at a specific time in gestation for the creation of fetal NTDs.
Fetal NTD model
Fetal NTDs were induced as previously described [14]. Briefly, after brief exposure to isoflurane (Abbot Laboratories, North Chicago, IL), chamber inhaled at 2%–4% in 100% oxygen, the dams received 60 mg/kg of all-trans retinoic acid (Sigma-Aldrich Chemical, St. Louis, MO) dissolved in olive oil at 10 mg/mL, through gavage, on embryonic day ten (E10), between 6:00 and 8:00 p.m.
Amniotic fluid procurement
In both groups, amniotic fluid samples from all fetuses (n=685) were obtained between E19 and E21 (term=E22). To that end, the dams were euthanized with chamber inhaled CO2. A large midline laparotomy was made and the bicornuate uterus eviscerated. Both horns were evaluated to determine the total number of fetuses. A separate set of 30G needle (Becton Dickinson, Franklin Lakes, NJ) on a 1-mL syringe (Becton Dickinson) was introduced into each amniotic cavity upon the ventral aspect of the fetus, through which amniotic fluid was procured. After procurement, the uterus and gestational membranes were incised and the fetus examined for the presence or absence of a NTD, namely, an isolated spina bifida, an isolated exencephaly, or a combination of the two.
Amniotic fluid processing
Amniotic fluid samples were divided into 2 groups, depending on the presence or absence of a NTD. Samples from all fetuses were processed. Whenever necessary, samples from more than one fetus were combined to create an aliquot of at least 1 mL before processing, which consisted of epigenetic selection for the presence of NSCs, followed by exposure to neural differentiation media. To that end, aliquots were centrifuged at 3,000 rpm for 3 min, and the pellets removed and resuspended in a commercially available rat NSC expansion medium (Millipore, Billerica, MA), supplemented with 20 ng/mL fibroblast growth factor (FGF)-2 (Promega, Madison, WI); 5 μg/mL amphotericin (X-Gen Pharmaceuticals, Big Flats, NY); the Dulbecco's modified Eagle's medium/F12, 1× B27 chemically defined medium, 2 mM L-glutamine; 1× penicillin; and 1× streptomycin (all from Millipore). The cell suspensions were plated in poly-L-ornithine/laminin-coated 6-well plates (BD Biosciences, Bedford, MA) and placed in a 95% humidified, 5% CO2 incubator at 37°C. The culture medium was changed every other day. Cells were maintained in culture for 6–8 weeks with 2–3 passages using an accutase solution (Millipore), after which time, a subset of cells derived from each aliquot underwent morphological and phenotypical analyses as specified below. The remaining cells were then exposed to the neural differentiation medium, which consisted of the rat NSC expansion medium described above (Millipore) without FGF-2 supplementation, also changed every other day. Cells were maintained in the differentiation medium for 5–7 days before morphological and phenotypical analyses.
Morphological and phenotypical analyses
Throughout processing, cells were inspected for morphological changes with an inverted microscope (Carl Zeiss, Jena, Germany). Upon completion of exposure to the NSC expansion and differentiation media, as described above, representative cell samples underwent immunostaining for the undifferentiated neural cell markers, Nestin and Sox-2, as well as markers for differentiation into the 3 major neural cell lineages, namely, the immature neuronal marker, β-III tubulin, the astroglial marker, glial fibrillary acidic protein (GFAP), and the oligodendrocyte maker, O4, as previously described [15]. Briefly, cells were fixed with 4% paraformaldehyde at room temperature for 10 min, and then washed 3 times with phosphate-buffered saline. After nonspecific antibody reactions were blocked with 10% goat serum (Sigma Chemical, St Louis, MO) and 0.1% triton X (Sigma) for 30 min, cells were incubated with primary antibodies directed against Nestin (5 μg/mL; Beckton Dickinson and Company); Sox-2 (1:50; Santa Cruz Biotechnology, Santa Cruz, CA); GFAP (1:80; Sigma); β-III tubulin (25 μg/mL; R&D Systems, Minneapolis, MN); and O4 (20 μg/mL; Millipore) overnight at 4°C. After 3 washes, cells were incubated with Dylight 488- or 549-conjugated goat secondary antibodies (Jackson Immunoresearch, West Grove, PA) for 1 h at room temperature. Adult rat hippocampal NSCs (Millipore) were used as positive controls. Amniotic mesenchymal stem cells obtained from the pregnant dams that were not exposed to retinoic acid were used as negative controls.
Results
None of the fetuses in the control up (n=267) had a structural abnormality, whereas at least one NTD was present in 52% (217/418) of the fetuses in the group exposed to retinoic acid. Such NTDs were distributed as follows: isolated spina bifida, n=144; isolated exencephaly, n=24; and a combination of the two, n=49.
During exposure to the NSC medium, cells derived from amniotic samples procured from fetuses with a NTD underwent cell division and proliferation in culture, exhibiting clusters commonly referred to as neurospheres, the morphological hallmark of neural progenitors (Fig. 1) [16]. Such cells were immunoreactive for the neural progenitor markers, Nestin and Sox-2 (Fig. 2). Upon exposure to the neural differentiation media, these cells became increasingly heterogeneous morphologically, in arrangements indistinguishable from what would be expected of differentiated neural cells of diverse lineages, many displaying neurites (Fig. 3). For this proposed initial nature of qualitative assessment, further differentiation between dendrites and axons was deemed unnecessary. Assorted subpopulations of these cells stained positively for any of the 3 major neural cell lineages tested; namely, β-III tubulin, GFAP, and O4 (Fig. 4), concurrently with downregulation of Nestin and Sox-2. Although we did not measure the exact proportion of neurons, astrocytes, or oligodendrocytes, no particular cell type proved dominant, with all appearing fairly evenly distributed.
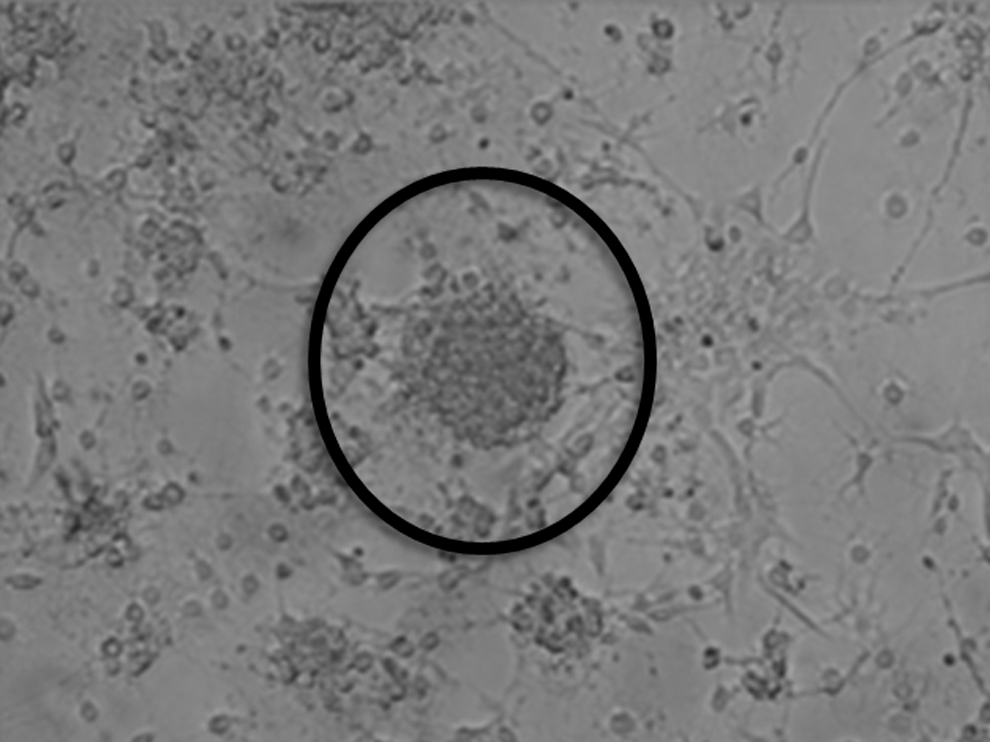
Phase-contrast microscopy of amniotic fluid-derived neural stem cells in culture forming neurospheres typical of undifferentiated neural progenitors. (Magnification 200×).
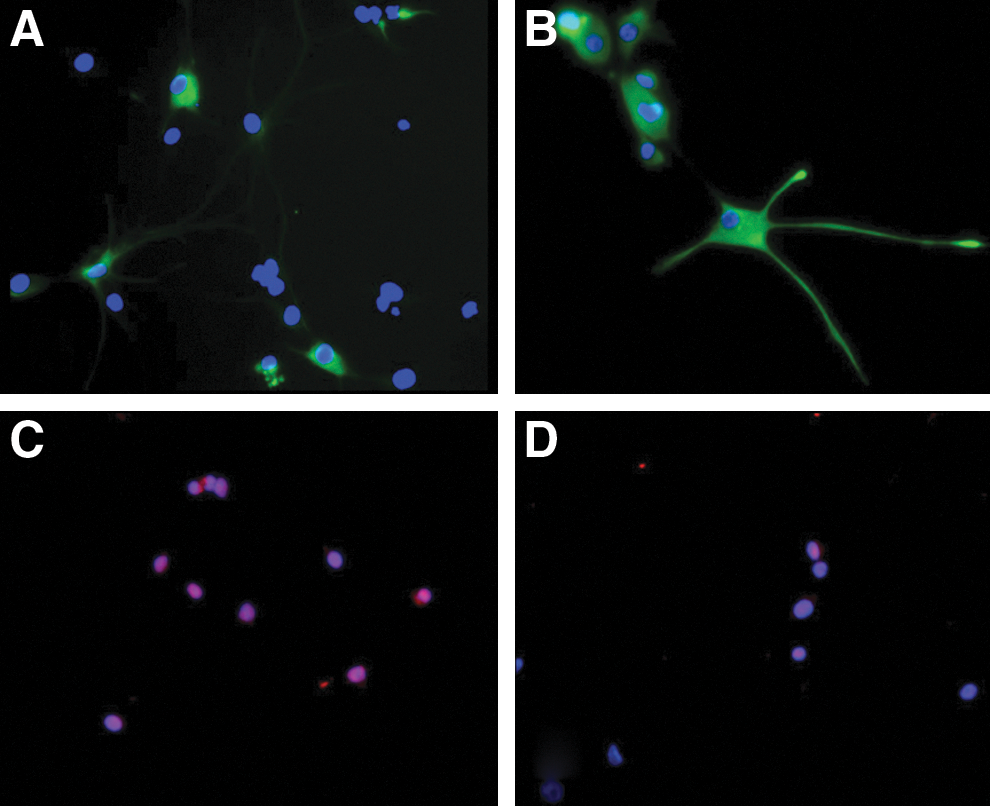
Undifferentiated amniotic fluid-derived neural stem cells
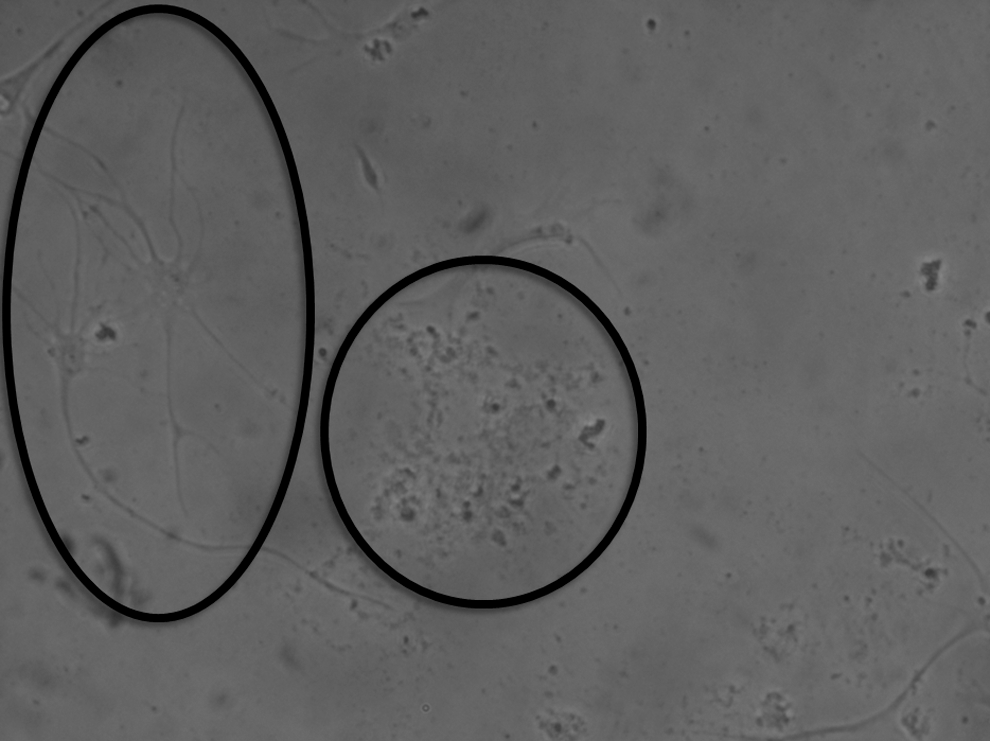
Phase-contrast microscopy of amniotic fluid-derived neural stem cells in neural differentiation medium exhibiting heterogeneous morphological changes expected of neural cells of diverse lineages, including neurites. (Magnification 200×).

Amniotic fluid-derived neural stem cells
Cells derived from amniotic samples procured in the control group did not proliferate, did not form neurospheres, nor exhibit any of the morphological changes observed during differentiation of the cells from the group with NTDs, as described above. These cells were negative for all of the neural markers tested.
Discussion
Spina bifida leads to spinal cord damage by both primary and secondary mechanisms. The former stems from the abnormal spinal cord development associated with the incomplete closure of the neural tube, while the latter is a result of spinal cord exposure to both the amniotic fluid (chemical insult) and local trauma (mechanical insult) within the uterus. Several studies point to the secondary component as being the most relevant clinically. Analyses performed in stillborns have shown that embryos, or very young fetuses, with spina bifida actually have little damage to the neural tube or spinal cord [17 –20]. Imaging studies have shown that fetal leg movement tends to be normal in fetuses with spina bifida early on in gestation, only to worsen as pregnancy progresses [21,22]. In addition, delivery of these fetuses by cesarean section is associated with improved neurological outcome, when compared with vaginal birth [23,24]. Animal models have further underlined the significance of the secondary insult to the spinal cord in this disease, also by demonstrating neurological improvement after experimental prenatal closure of the defect [25 –27]. All these data have justified fetal repair clinically, as performed at the MOMS.
Following transplantation from an exogenous source, NSCs have been shown to mediate repair in a multitude of central nervous system injury or disease models, including in the spinal cord, often by invoking developmental mechanisms [15,28 –35]. In essentially all of these conditions, however, NSCs have been administered after birth. In our previous study, we introduced the notion of prenatal delivery of NSCs as a potential means of enhancing fetal surgical coverage of spina bifida, which is fundamentally a developmental problem [9]. Our fetal approach was predicated on the hypothesis that the regenerative/reparative impact of NSCs could be maximized if applied to the developing, rather than the more mature, spinal cord. Necessarily, however, our early work involved a well-characterized NSC clone xenologous to the host. At this time, any realistic hopes of clinical translation of this therapeutic concept would necessarily hinge on the consistent availability of a heterologous, or, preferably, an autologous prenatal source of NSCs.
While neural cells have long been known to exist in amniotic fluid in the presence of NTDs, and, in fact, often aid in the diagnosis of these anomalies, the existence of an undifferentiated, more primitive population of neural cells within the amniotic cavity had yet to be demonstrated [10 –13,36]. In this study, we have isolated a population of NSCs in the setting of experimental NTDs. Due to the fact that, in this rodent model, samples from more than one fetus with the same type of NTD, at times, had to be combined to generate an aliquot sizeable enough for processing, we could not quantify the likelihood of such an isolation being viable in each fetus. On the other hand, all of the samples from fetuses with NTDs with at least 1 mL yielded cells. Also, such quantitative data in rodents would arguably be of limited translational significance. Previous data on the abundant presence of neural cells in human amniotic fluid in the setting of fetal NTDs would suggest that a comparable isolation of NSCs may well be practical clinically [36]. Such a demonstration is one of our current endeavors.
We have previously established experimentally that the mesenchymal stem cells long known to be naturally occurring in the amniotic fluid can be used in tissue engineering strategies for the repair of a variety of congenital anomalies in the perinatal period, and that these cells can be manufactured within regulatory guidelines for clinical use [37 –45]. Translated clinically, from a simple amniocentesis, different grafts could be engineered, in parallel, to the remainder of gestation, so that an infant could benefit from having autologous, expanded tissue promptly available for surgical reconstruction at or before birth. In a similar fashion, the present study suggests that autologous NSCs could be obtained by amniocentesis in a fetus with spina bifida and expanded ex vivo for therapeutic applications, either prenatally or postnatally. This is also, to our knowledge, the first demonstration of disease-specific stem cells in the amniotic fluid.
The inherent limitations of an introductory study such as this notwithstanding, our findings, taken together with the large body of data on the use of NSCs in other forms of spinal cord injury, support further development of this multifaceted, perinatal therapeutic approach, combining local NSC delivery with mechanical/surgical strategies aimed at inducing protective and/or regenerative processes within the spinal cord in spina bifida.
Footnotes
Acknowledgment
D.O.F. was supported by a Translational Research Program Award from Children's Hospital Boston. Y.D.T. and his group were supported by DoD-CIMIT, VARR&D, and HeadNorth Project funding.
Authors Disclosure Statement
No competing financial interests exist.