
Abstract
CD133+ cells purified from hematopoietic tissues are enriched mostly for hematopoietic stem/progenitor cells, but also contain some endothelial progenitor cells and very small embryonic-like stem cells. CD133+ cells, which are akin to CD34+ cells, are a potential source of stem cells in regenerative medicine. However, the lack of convincing donor-derived chimerism in the damaged organs of patients treated with these cells suggests that the improvement in function involves mechanisms other than a direct contribution to the damaged tissues. We hypothesized that CD133+ cells secrete several paracrine factors that play a major role in the positive effects observed after treatment and tested supernatants derived from these cells for the presence of such factors. We observed that CD133+ cells and CD133+ cell-derived microvesicles (MVs) express mRNAs for several antiapoptotic and proangiopoietic factors, including kit ligand, insulin growth factor-1, vascular endothelial growth factor, basic fibroblast growth factor, and interleukin-8. These factors were also detected in a CD133+ cell-derived conditioned medium (CM). More important, the CD133+ cell-derived CM and MVs chemoattracted endothelial cells and display proangiopoietic activity both in vitro and in vivo assays. This observation should be taken into consideration when evaluating clinical outcomes from purified CD133+ cell therapies in regenerative medicine.
Introduction
A
One possibility in explaining these outcomes is the paracrine effect of cells employed for therapy [4]. In support of this possibility, evidence has accumulated that stem cells secrete a variety of growth factors, cytokines, chemokines, and bioactive lipids that interact with the surrounding microenvironment, and if used in therapy, affect cells in damaged organs [5 –11]. These factors are secreted particularly from activated stem cells that have been removed from their physiological niches (eg, aspirated from the BM) or mobilized into the circulation (eg, mPB or UCB) and potentially (i) inhibit apoptosis of cells residing in the damaged organs, (ii) stimulate proliferation of these cells, and (iii) promote vascularization of affected tissues to improve oxygen delivery and metabolic exchange. In addition to soluble growth factors, cytokines, and chemokines, activated stem cells also secrete microvesicles (MVs) [9 –12]. MVs are small, spherical membrane fragments shed from the cell surface or secreted from the endosomal compartment and play an important and under-appreciated role in cell-to-cell communication [9 –12]. Overall, these cell-derived paracrine signals may explain the therapeutic benefits of adult stem cells employed in regeneration of, for example, heart AMI.
By employing reverse transcription–polymerase chain reaction (RT-PCR) in our previous work, we found that highly purified human CD34+ HSPCs express several mRNA transcripts for growth factors, cytokines, and chemokines, and subsequently we confirmed their presence in a conditioned medium (CM) harvested from these cells by employing sensitive ELISA [5,6]. Moreover, in vitro functional studies revealed that a medium conditioned by human CD34+ cells may inhibit apoptosis, stimulate proliferation, and chemoattract several other types of cells, including endothelial cells [5,6]. Our observations demonstrating CD34+ cells as a source of paracrine signals were recently confirmed in an elegant study performed by another group [7].
Since BM-, mPB-, and UCB-derived CD133+ cells are, in addition to CD34+ cells, a potential source of purified stem cells in regenerative medicine for organ repair, we asked whether highly purified human CD133+ cells, which are akin to CD34+ cells, also secrete factors that could play a beneficial paracrine role in regeneration of damaged organs and tissues. We observed that highly purified UCB-derived CD133+ cells express mRNAs and secrete proteins for several soluble factors [eg, vascular endothelial growth factor (VEGF), kit ligand, basic fibroblast growth factor (FGF-2), and insulin growth factor-1 (IGF-1)] and shed MVs from the cell surface and endosomal compartment. These factors possess antiapoptotic properties, increase the in vitro cell survival of endothelial cells, and stimulate their proliferation and tube formation. This important observation suggesting an important role for CD133+ cell-derived paracrine signals has to be considered when evaluating clinical outcomes using purified CD133+ cells in regenerative medicine.
Materials and Methods
Human UCB-derived CD133+ cells
Human UCB cells were obtained after informed consent from 19 donors, and the protocols used were approved by the appropriate institutional ethics review boards. Light-density cells after Ficoll/Paque centrifugation were enriched for CD133+cells by immunoaffinity selection with AutoMACS paramagnetic beads (Miltenyi Biotec). The purity of isolated CD133+ cells was >95% as determined by fluorescence activated cell sorter (FACS) analysis employing an LSRII cytometer.
Human umbilical endothelial cells
Human umbilical endothelial cells (HUVECs) were isolated from umbilical cord as described previously [13], and aliquots of cells were kept frozen until used for experiments.
Collection of CM and MVs from CD133+ cells
CD133+ cells were incubated overnight at the concentration 5×106 cells/1 mL RPMI [with 0.5% bovine serum albumin (BSA)]. On the next day, cells were spun down and pelleted for RNA isolation, and the CM was collected. CM was subsequently centrifuged at 2,000 g for 15 min at 4°C, and the MV-enriched supernatants were collected. These supernatants were again centrifuged at 28,000g for 1 h at 4°C. The pellets were than washed and suspended in a HEPES buffer, pH 7.4. The protein content in CD133+ cell-derived MVs was evaluated using Bradford assay as described by us [13].
Collection of MVs from human PB platelets
Human PB platelets were isolated from healthy volunteer donors who had given informed consent; the protocols used were approved by the Institutional Review Board of the University of Louisville. The platelets were activated by thrombin (0.1 U/mL) and collagen (4 μg/mL) for 30 min at 37°C with stirring, and centrifuged twice at 3,000 rpm (2,000 g) for 15 min at 4°C. The platelet-derived microvesicle (PMV)-enriched supernatants were collected and centrifuged at 100,000g for 60 min at 4°C. These PMV enriched in exosomes were washed once and resuspended in HEPES buffer, pH 7.4. The concentrations of the PMV were estimated by Bradford assay as described [13].
Transmission electron microscopy analysis of CD133+ cell-derived MVs
For transmission electron microscopy (TEM), the UCB CD133+ cell-derived MVs were fixed in 3% glutaraldehyde in 0.1 M cacodylate buffer, pH 7.4, for 10 h at 4°C, postfixed in osmium tetride, and dehydrated. Fixed cells were subsequently embedded in LX112 and sectioned at 800 Å, stained with uranyl acetate and lead citrate, and viewed on a Philips CM10 electron microscope operating at 80 kV.
Real-time quantitative reverse transcription–PCR
Total RNA was isolated from the purified CD133+ cells, CD133+ cell-derived MVs, and UCB mononuclear cells (MNCs) with the RNeasy Kit (Qiagen). The RNA was reverse transcribed with MultiScribe reverse transcriptase and oligo-dT primers (Applied Biosystems). Quantitative assessment of mRNA levels was performed by real-time RT-PCR on an ABI 7500 instrument with Power SYBR Green PCR Master Mix reagent. Real-time conditions were as follows: 95°C (15 s), 40 cycles at 95°C (15 s), and 60°C (1 min). According to melting point analysis, only one PCR product was amplified under these conditions. The relative quantity of a target, normalized to the endogenous control β-2 microglobulin gene and relative to a calibrator, is expressed as 2-ΔΔCt (the fold change), where Ct is the threshold cycle, ΔCt=(Ct of target genes)−(Ct of endogenous control gene, β-2 microglobulin), and ΔΔCt=(ΔCt of samples for target gene)−(ΔCt of calibrator for the target gene). The sequences of primers employed for real-time quantitative reverse transcription–PCR (RQ-PCR) are shown in Table 1.
IGF-1, insulin growth factor-1; VEGF, vascular endothelial growth factor; FGF-2, basic fibroblast growth factor; HGF, hepatocyte growth factor; IL, interleukin; TNF, tumor necrosis factor; TGF, transforming growth factor; MIF, migration inhibitory factor; SDF-1, stromal-derived factor-1; MIP-1α, macrophage inflammatory protein-1α; SCF, stem cell factor; IP, inflammatory protein; IFN-γ, interferon-γ.
ELISA measurements
The secretion of growth factors by CD133+ cells was evaluated by Quantikine human immunoassays (R&D Systems) according to the manufacturer's protocol. Equal numbers (1×106 cells/mL) of CD133+ cells were cultured for 24 h in serum-free conditions before harvesting of the CM. The collected media were analyzed by a quantitative ELISA technique. The sensitivity of the ELISA assays (pg/mL) was as follows: hepatocyte growth factor (HGF), >40; VEGF, >5; FGF-2, >3; IGF-I, >12; FLT3 ligand, >7; and stem cell factor (SCF), >3. The data are shown as a fold difference compared to the medium alone.
Chemotaxis assay
The 8-μm polycarbonate membranes were covered with 50 μL of 0.5% gelatin. HUVECs expanded in vitro were detached from culture dishes with 0.5 mmol/L ethylenediaminetetraacetic acid, washed in RPMI 1640, resuspended in RPMI 1640 with 0.5% BSA, and seeded at a density of 3×104 in 120 μL into the upper chambers of Transwell inserts (Costar Transwell; Corning Costar). The lower chambers were filled with the CM harvested from CD133+ cells, with a suspension of CD133+ cell-derived MVs (30 μg/mL), or with the CM harvested from CD133+ cells from which MVs had been removed (w/o MV). As positive controls, we employed human recombinant FGF-2 (50 ng/mL), or as a negative control, 0.5% BSA RPMI 1640. After 24 h, the inserts were removed from the Transwell plates. Cells remaining in the upper chambers were scraped-off with cotton wool, and cells that had transmigrated were stained with HEMA 3 according to the manufacturer's instructions (Fisher Scientific) and counted either on the underside of the membranes or in the bottom chamber of the Transwell plates.
Phosphorylation of intracellular pathway proteins
Western blotting was performed on extracts prepared from HUVECs that had been kept in an RPMI medium containing low levels of BSA (0.5%) to render them quiescent. The cells were divided into aliquots of 2×106 cells and stimulated by the CM from CD133+ cells (1 mL), CD133+ cell-derived MVs (30 μg/mL), CM from CD133+ cells w/o MV (1 mL), FGF-2 (50 ng/mL), or medium alone for 5 min at 37°C, and then incubated (for 10 min) on ice in an M-PER lysing buffer (Pierce) containing protease and phosphatase inhibitors (Sigma). The extracted proteins were then separated by 12% dodecyl sulfate–polyacrylamide gel electrophoresis and the fractionated proteins transferred to a nitrocellulose membrane (Schleicher & Schuell). Phosphorylation of the intracellular kinases, p44/42 mitogen-activated protein kinase (MAPK, Thr 202/Tyr 204), and AKT was detected using commercial mouse phosphospecific p44/42 MAPK mAb or rabbit phosphospecific polyclonal antibodies (all from New England Biolabs) with horseradish peroxidase-conjugated goat anti-mouse immunoglobulin G (IgG) or goat anti-rabbit IgG as secondary antibody (Santa Cruz Biotech). Equal loading in the lanes was evaluated by stripping the blots and reprobing with anti-p44/42 MAPK mAb (clone# 9102; Santa Cruz Biotech). The membranes were developed with an enhanced chemiluminescence reagent (Amersham Life Sciences), dried, and exposed to film (HyperFilm; Amersham LifeSciences).
Human endothelial tube formation assay
Endothelial tube formation was assessed on a synthetic basement membrane according to the manufacturer's protocol (Matrigel; BD Biosciences). Briefly, the matrix was thawed overnight at 4°C and polymerized at 37°C for 30 min before use. HUVECs were resuspended in a reduced-serum medium with 0.5% BSA (negative control), a medium supplemented with FGF-2 (50 ng/mL, as positive control), CM from CD133+ cells (1 mL), CM from CD133+ cells w/o MV (1 mL), or with CD133+ cell-derived MVs (30 μg/mL). The cells were subsequently seeded (3×103/cm2) on the Matrigel and the plates placed in a humidified atmosphere of 5% CO2 at 37°C. Four hours later, identical fields in each well were inspected, and the number of formed tubes was enumerated, and the field was photographed. All conditions were tested in duplicate wells in 3 separate experiments. In some of the tube formation assays, we compared the proangiopoietic activity of CD133+ cell-derived MVs (30 μg/mL) to human PMVs (30 μg/mL).
Matrigel implants in SCID mice
Matrigels were implanted in mice UCB CD133+ cell-derived MVs. The angiogenesis assays were carried out by injecting SCID mice with 0.5 mL Matrigel (Collaborative Biomedical Products) mixed on ice with 30 μg/Matrigel of CD133+ cell MVs or PMVs or not (controls). Matrigel was injected subcutaneously in the abdominal midline on day 0. The Matrigel implants were isolated after 1–2 weeks and photographed; the extent of vascularization was later studied by determination of hemoglobin content using the Drabkin method as described by us [14]. Briefly, the Matrigel implants were harvested, and all surrounding tissues dissected away. Cell pellets were homogenized at 4°C and assayed for hemoglobin content according to the manufacturer's protocol (Drabkin's reagent kit; Sigma Diagnostics).
Statistical analysis
All data were analyzed using one-factor analysis of variance with Bonferroni's multiple comparison test. We used the Instat1.14 program (GraphPad Software), and statistical significance was defined as P<0.05 or P<0.01.
Results
Human CD133+ cells express and secrete several antiapoptotic and proangiopoietic factors
To address whether purified human CD133+ cells, such as CD34+ cells [5,7], express factors that affect tissue/organ regeneration, we evaluated mRNA expression for several growth factors, including SCF, IGF-1, VEGF, FGF-2, and HGF in mRNA extracts isolated from these cells. Furthermore, based on our previous observation that stem cells secrete MVs enriched in mRNA [15], in parallel we isolated and evaluated the expression of these mRNA species in CD133+ cell-derived MVs. The representative purity of UCB CD133+ cells is shown at Fig. 1A, and FACS as well as transmission electron microscopy (TEM) characteristics of UCB CD133+ cells-derived MVs are demonstrated at Fig. 1B. We observed MVs of different sizes mostly >100 nm corresponding to cell surface shed MVs, but also some MVs <100 nm derived from an endosomal compartment called exosomes. The FACS-based analysis (Fig. 1C) reveled expression of CD34, CXCR4, CD117 (c-kit), and CD135 (FLT3) antigens on their surface, what supports our observation that UCB CD133+ cell-derived MVs may express on the surface receptors characteristic for cells of their origin [5].
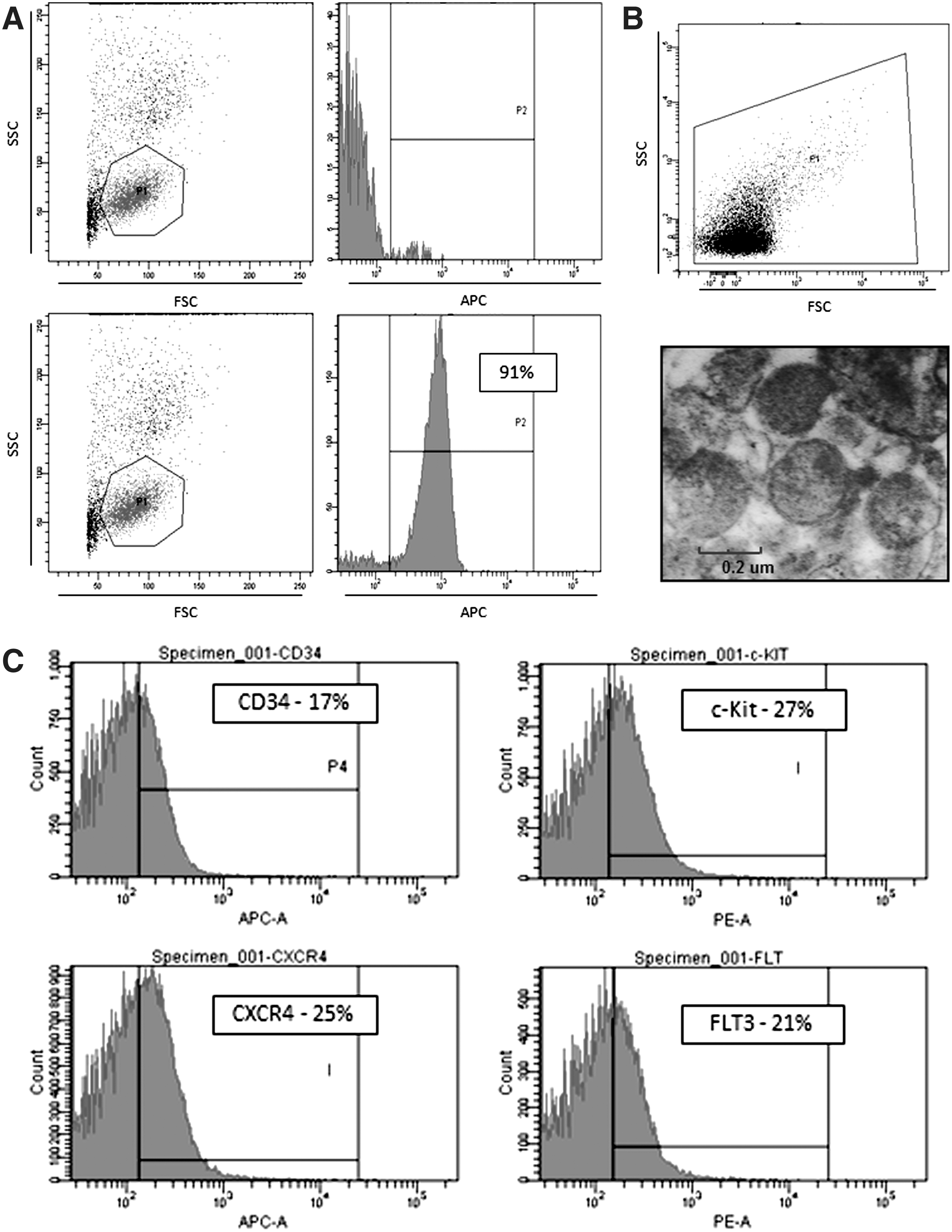
Purity of isolated from umbilical cord blood (UCB) CD133+ cells and fluorescence activated cell sorter (FACS) and transmission electron microscopy (TEM) analysis of UCB CD133+ cell-derived microvesicles (MVs).
Figure 2A shows real-time PCR analysis of mRNA expression for these factors in purified CD133+ cells and CD133+ cell-derived MVs. We observed mRNA enrichment for all these factors, and in particular, high expression was observed for FGF-2 and IGF-1 mRNAs. The expression of these factors was subsequently confirmed at the protein level in CM derived from highly purified CD133+ cells by sensitive ELISA (Fig. 2B). As shown, we observed a high level of VEGF protein in the supernatants derived from these cells. Thus, human CD133+ cells, such as CD34+ cells, are a potential source of important paracrine factors that may play a role in tissue/organ regeneration by inhibiting apoptosis (eg, IGF-1 or SCF) and stimulating angiogenesis (eg, VEGF).
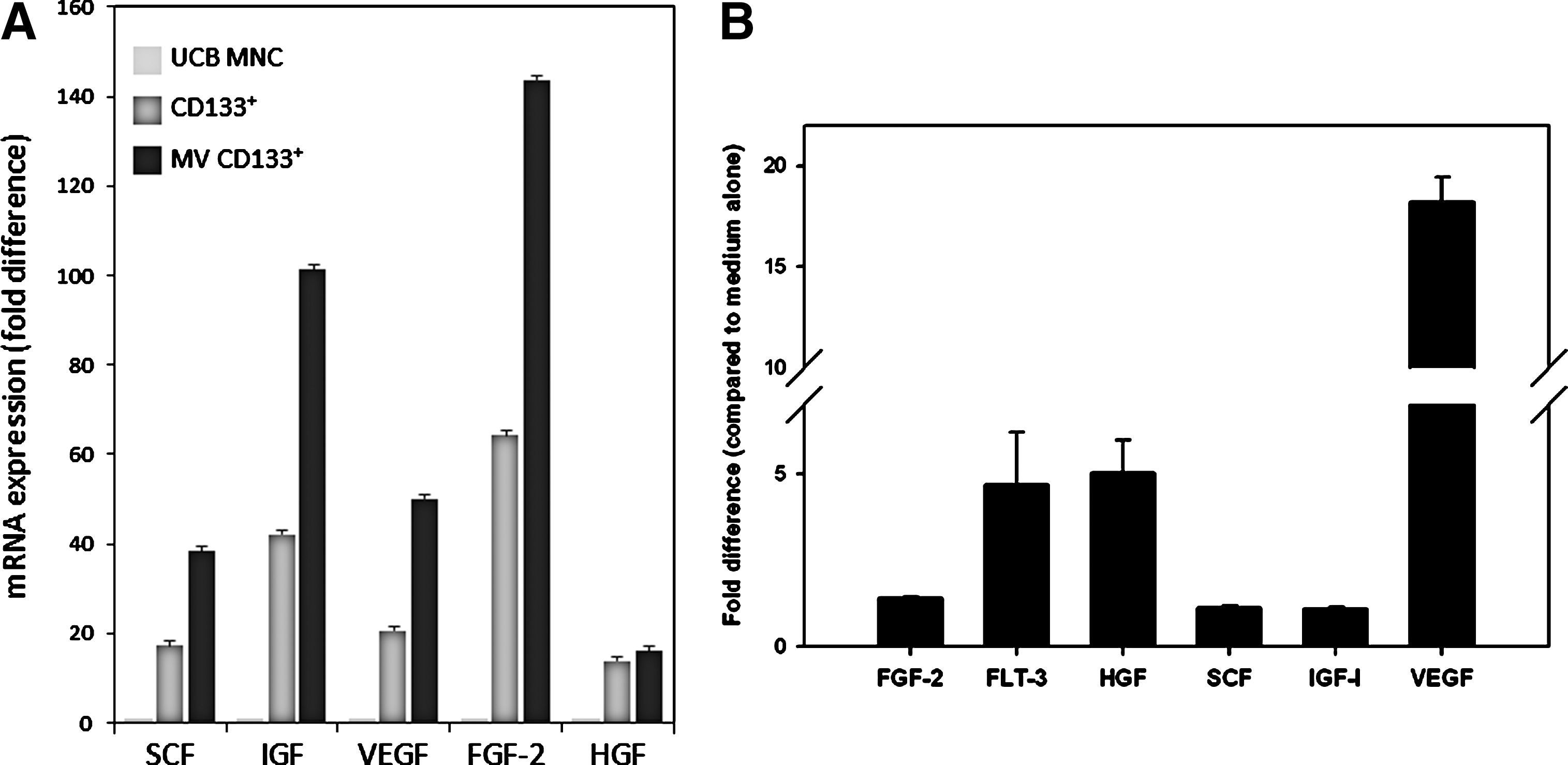
Expression of selected proangiopoietic factors by human CD133+ cells, CD133+ cell-derived MVs, and CD133+ cell-derived conditioned media (CM).
We also evaluated mRNA expression for selected cytokines and chemokines (Table 2) and observed that both CD133+ cells as well as CD133+ cell-derived MVs are highly enriched for interleukin-8 (IL-8), IL-16, RANTES, macrophage migration inhibitory factor, and IP-10 mRNAs. However, at the same time, we did not detect mRNA expression for IL-1α, stromal-derived factor-1, macrophage inflammatory protein-1α (MIP-1α), and interferon-γ. Interestingly, we also observed that CD133+ cells and CD133+ cell-derived MVs express tumor necrosis factor-alpha (TNF-α), TNF-β, and transforming growth factor-β mRNAs.
−, < 1-fold increase; +, < 10-fold increase; ++, < 100-fold increase; +++, < 1,000-fold increase; ++++, > 1,000-fold increase compared to umbilical cord blood mononuclear cells; MV, microvesicle; MIP-1α, macrophage inflammatory protein-1α.
CD133+ cell-derived CM stimulate HUVECs
In our previous work, we demonstrated that the CM harvested from CD34+ cells stimulated migration of HUVECs [5]. We therefore became interested in whether the CM harvested from CD133+ cells also stimulates migration of these cells.
To address this question, we first studied activation of intracellular signaling (phosphorylation of p44/42 MAPK and AKT) in human umbilical cord vein endothelial cells (HUVEC) stimulated by CD133+ cell-derived CM, CD133+ cell-derived CM from which MVs have been removed by high-speed centrifugation, and CD133+ cell-derived MVs (Fig. 3A). In parallel, we evaluated the chemotactic responsiveness of HUVECs to the same CD133+ cell-derived CM and MVs (Fig. 3B). In both experiments, FGF-2 and the medium were used as positive and negative controls, respectively.

CD133+ cell-derived CM and MVs stimulate signaling pathways and chemoattract human umbilical endothelial cells (HUVECs).
Figure 3A shows phosphorylation of AKT and p44/42 MAPK in HUVEC signaling pathways, which are activated during HUVEC migration, as previously demonstrated [5]. We observed that all CD133+ cell-derived CM and MVs strongly activated phosphorylation of AKT and p44/42 MAPK in these cells. Furthermore, Figure 3B demonstrates that the CM and MVs derived from CD133+ cells strongly chemoattract HUVECs in the Transwell migration assay. The chemotaxis of HUVECs was inhibited after they were preincubated with the AKT and p44/42 MAPK small-molecule inhibitors, LY 294002 and U0126, respectively (not shown). Based on this finding, we concluded that human CD133+ cells secrete paracrine factors (growth factors and MVs) that chemoattract human endothelial cells in a p44/42 MAPK- and AKT-dependent manner. Of note, the chemotactic activity of our CD133+ cell-derived supernatants was even stronger than for FGF-2, which had been employed at its optimal dose for stimulating maximal HUVEC migration.
CD133+ cell-derived MVs stimulate angiogenesis in vitro
Finally, to evaluate whether the CD133+ cell-derived CM and MVs stimulate angiogenesis, we employed an in vitro functional tube formation assay. Figure 4A and B show that soluble factors present in the CM that are secreted from these cells, and in particular, CD133+ cell-derived MVs strongly stimulate HUVECs to form tubes in Matrigel cultures. This proangiopoietic effect of CD133+ cell-derived MVs was comparable to MVs isolated from activated human platelets (PMVs).

Tube formation by HUVECs in the presence of CD133+ cell-derived CM and MVs.
Finally, we tested the angiogenic potential of UCB CD133+ cell-derived MVs and human PMVs in vivo by examining the development of vascularization-dependent hemoglobinization of Matrigel plugs implanted subcutaneously into SCID mice as described by us in the past [14]. We noticed in 2 independent experiments (3 mice/group) that Matrigels containing UCB-CD133+ cell MVs become hemoaglobinized to a similar degree as Matrigels containing PMVs [5.3±2.1, 4.8±2.7 and 0.3±0.1 of Hb mg/g of Matrigel for UCB CD133+ cell-derived MVs, PMVs, and empty Matrigel (negative control); respectively]. These functional assays in vitro and in vivo further support the notion that CD133+ cells stimulate angiogenesis in a paracrine-dependent manner.
Discussion
The field of regenerative medicine is searching for stem cells that can be safely and efficiently employed for regeneration of damaged organs [16 –18]. The first cells that have been widely employed in animal models and patients to regenerate damaged organs were MNCs isolated from BM, enriched for HSPCs. However, despite initial exciting and promising reports postulating plasticity of these cells, their role in the repair of damaged organs has become controversial. Further experiments with highly purified populations of HSPCs showed them not to be able to replace cells in damaged tissues as seen, for example, in animal models of a regenerating heart [19] or brain [20].
In the meantime, various types of stem cells isolated from adult tissues are being employed in several clinical trials (eg, to treat myocardial infarction, liver damage, ischemic kidney failure, or stroke) [1 –4]. These cells are isolated most frequently from BM, mPB, or UCB and are enriched mostly for HSPCs, but also contain some rare endothelial progenitor cells (EPCs), mesenchymal stem cells, and very small embryonic-like stem cells (VSELs). However, the ideal stem cell for regenerative medicine would be pluripotent stem cells, which according to their definition have a broad potential to differentiate into cells from all 3 germ layers (mesoderm, ectoderm, and endoderm).
CD133 antigen (prominin) is a 4-transmembrane domain-spanning protein expressed on the surface of HSPCs, EPCs, and VSELs [21]. Development of specific antibodies against this antigen conjugated with paramagnetic beads enabled a fast and convenient strategy to purify these cells from hematopoietic tissues. However, while some CD133+ cells simultaneously express CD34+ antigen, there are still 40%–80% of CD133+ cells that are CD34 negative [21 –23], which may explain some differences in in-vivo biological effects between cell preparations. Since compared to the population of CD34+ cells, the population of CD133+ cells is more enriched for the most primitive stem cells that seem to be somewhat more suitable for application in regenerative medicine [23].
We previously demonstrated that purified normal human BM- and mPB-derived CD34+ HSPCs express mRNAs for various growth factors, cytokines, and chemokines, including VEGF, HGF, SCF, IGF-1, FGF-2, IL-8, and RANTES; more importantly, these factors were subsequently detected by ELISA in a medium conditioned by these cells [5]. Moreover, in vitro functional studies revealed that media conditioned by CD34+ cells inhibit apoptosis, stimulate proliferation, and chemoattract several types of cells, including endothelial cells [5,6]. These observations demonstrating CD34+ cells to be the source of several paracrine factors were recently confirmed in an elegant study by another group [7].
In our current study we observed that UCB-derived CD133+ cells express a similar repertoire of growth factors, cytokines, and chemokines as CD34+ cells. There were, however, some small differences; for example, CD133+ cells, in contrast to CD34+ cells, do not express MIP-1α mRNA. What is additionally important, in our current work we performed RQ-PCR analysis of mRNA expression and observed that, compared to UCB MNCs, not only CD133+ cells but also CD133+ cell-derived MVs are highly enriched in mRNAs for several factors.
In addition to soluble factors, MVs also play an important role in organ tissue regeneration [4,9] as signaling devices in cell–cell communication—a mechanism that for many years has been largely overlooked. In several experimental tissue injury models, MVs can even replace intact cells employed in cellular therapies to improve the function of damaged organs [9 –11]. MVs may be shed from the cell surface or derived from the intracellular membrane compartment of normal healthy cells in a membrane-blebbing mechanism and hijack both membrane components and the engulfed cytoplasmic contents [9 –11].
Like apoptotic bodies, MVs have for many years been regarded as mere cellular fragments or debris. However, it turns out that MVs not only contain numerous proteins and lipids similar to those present in the membranes of the cells from which they originate, but since MV membranes engulf some cytoplasm during membrane blebbing, they may also contain intracellular proteins, mRNAs, regulatory miRNAs, and even intact organelles such as mitochondria that could be transferred to target cells [9 –11]. In this molecular transfer process, MVs act as naturally engineered liposomes. Interestingly, it has been demonstrated that mRNA and miRNA molecules are somehow preferentially enriched within MVs by a mechanism that involves docking proteins. To explain this phenomenon, cells under steady-state conditions tend to store mRNA and miRNA for later utilization under stress conditions, which explains why they can release these molecules into the extracellular space encapsulated within MVs. This stem cell-derived MV horizontal transfer of proteins, mRNA, and miRNA [15,24,25] was first demonstrated in a model of expansion of murine HSPCs in the presence of murine embryonic stem cell (ESC)-derived MVs [15] and was subsequently confirmed in several other elegant studies showing transfer of hematopoietic cell- or mesenchymal cell-derived MVs into a damaged lung [24] or liver [25].
However, while some cell-of-origin-specific expression of biologically active components in MVs has been observed, others may be commonly shared between cells. For example, bioactive lipid components of cell membranes, including sphingosine-1-phosphate (S1P), ceramide-1-phosphate (C1P), and lysophosphatidylic acid, are highly enriched within all types of MVs. In particular, since S1P and C1P inhibit cell apoptosis and stimulate angiogenesis [26,27], they can be envisioned as important paracrine factors delivered by all types of MVs.
We demonstrate here that CD133+ cell-derived MVs, as reported previously for MVs isolated from CD34+ cells, blood platelets, or even more differentiated BM-derived hematopoietic cells, directly chemoattract endothelial cells [7,12,28]. However, in contrast to our data [12], a recent report has proposed that the proangiopoietic effect of MVs is related to the stem cell phenotype, and that more differentiated cells lack this potential [7].
To address this question, it should be remembered that membrane lipid components such as S1P are endowed with strong proangiopoietic potential and are ubiquitously present in MVs secreted by all types of cells. Therefore, we propose instead that MVs, regardless of the cells from which they originate, are very strong chemoattractants for endothelial cells. Furthermore, since CD133+ cell-derived MVs are enriched for mRNAs encoding several proangiopoietic factors, additional studies are needed to see whether MV-delivered RNA species are translated into appropriate peptides after transfer into target cells as we have previously shown for ESC-derived MVs [15]. In addition, since MVs may transfer miRNAs between cells, additional studies are also needed to evaluate the composition of regulatory miRNAs in CD133+ cell-derived MVs. Further studies should also dissect a contribution of soluble factors versus MVs to observed effects as well as similarities and differences in biological activity of cell surface-shed MVs to smaller MVs called exosomes that are derived from the endosomal compartment.
In conclusion, we have demonstrated that purified human CD133+ cells, which are akin to purified CD34+ cells [5,7], are a source of several soluble factors and MVs that promote angiogenesis. Based on these findings, the rescue of damaged organs or tissues after an infusion of stem cells could be explained by the paracrine and antiapoptotic factors contained within MVs secreted from these cells after their delivery to the damaged organs. This important phenomenon awaits further exploitation, for example, by optimizing the secretion of proangiopoietic factors and MVs by cells employed in therapeutic approaches in regenerative medicine [27].
Footnotes
Acknowledgments
This work was supported by Maestro 2011/02/A/NZ4/00035 grant to MZR.
Author Disclosure Statement
No competing financial interests exist.