
Abstract
Human embryonic stem cells (hESCs) self-renew indefinitely as highly organized pluripotent colonies. Unlike mouse pluripotent stem cell colonies, human colonies form a uniform, flat, epithelium-like monolayer. Interestingly, it has been reported that colony morphology is closely correlated with the maintenance of pluripotency. However, the molecular mechanisms that underlie human pluripotent colony formation and organization are poorly understood. In this study, we used real-time imaging tools to examine the in vitro colony formation of enzymatically dissociated single hESCs under feeder-free conditions. We demonstrate that colony formation consists of 4 stages: attachment, migration, aggregation, and colony formation, which are facilitated in an intracellular, calcium-dependent manner. Moreover, we found that blocking Gi-coupled G protein-coupled receptor (GPCR) signaling results in enhanced cell–cell interactions and plays an integral role in promoting the survival of hESCs in culture. From the imaging results, we identified the conditions required for colony formation, and we identified the importance of blocking Gi-coupled GPCR by pertussis toxin in modulating hESC colony formation and organization. These results will likely be useful for engineering hESCs to further study the mechanisms involved in their function.
Introduction
H
Interestingly, unlike mouse ESC colonies, hESC colonies form flat epithelium-like monolayers in culture [11]. Because colony morphology is closely correlated with the maintenance of pluripotency, understanding the mechanisms by which colonies form and become organized will likely be critical for controlling hESC growth and differentiation.
hESC colonies are generally vulnerable to apoptosis upon cellular dissociation [1,2]. In particular, they undergo massive cell death after complete dissociation, and the colony formation efficiency observed for dissociated hESCs is generally <1% [12,13]. Therefore, hESCs are difficult to use in dissociated single-cell cultures, which are useful for procedures such as clonal isolation after gene transfer and differentiation induction. In an effort to improve the poor survival rate of hESCs after cell dissociation, researchers have found that the application of the selective Rho-associated kinase (ROCK) inhibitor Y27632 markedly diminishes dissociation-induced apoptosis [14]. The potential mechanism of the effect of Y27632 on apoptosis has also been studied [15 –17]; recent reports have indicated its possible utility during certain modified stem cell culture conditions [14,18 –21]. However, the success of this simple culturing protocol depends on the use and integration of the following 3 critical elements: (i) adapting high-quality undifferentiated hESCs to enzymatic passaging, (ii) seeding the appropriate density of cells, and (iii) using feeder cells [14,22,23]. Unfortunately, the procedures for culturing enzymatically dissociated single hESCs meet none of these requirements. Therefore, the formation of colonies from these cell types has been less than optimal.
The molecular mechanisms involved in hESC colony formation and organization are poorly understood. In an attempt to understand these processes, we focused on the finding that hESC colony formation shares distinct similarities with cell-cell adhesive interactions of mammary epithelial cells [24]. Because G protein-coupled receptor (GPCR) signaling is known to be involved in epithelial cell aggregation, these pathways represent attractive candidates for modulators of hESC colony formation [25 –28]. GPCRs transmit information within cells via 2 major signaling pathways, the regulation of phospholipase C (PLC) and protein kinase C (PKC) activity [29]. These signaling pathways are specifically activated by receptor-associated G proteins; for example, Gs- and Gi-coupled receptors modulate cyclic adenosine monophosphate levels [30]. In particular, Gi-coupled GPCR signaling is a conserved developmental regulator of cell shape and migration [31,32].
In this study, we hypothesized that hESC colony formation and the maintenance of characteristic pluripotent colony morphology and organization are mediated by intracellular calcium and Gi-coupled GPCR signaling. To test this hypothesis, we studied 2 different hESC cell lines, and we treated pluripotent colonies with toxins [pertussis toxin (PTX) or cholera toxin (CTX)] to modulate GPCR signaling. Using live-cell-imaging tools, we demonstrated that single hESCs progress through the 4 following stages of colony formation: attachment, migration, aggregation, and colony formation. We found that cell–cell interactions, which were artificially increased by PTX treatment, play a pivotal role in colony organization and the maintenance of pluripotency by inhibiting PLCβ-independent Gi-coupled GPCR signaling.
Materials and Methods
Maintenance culturing of hESCs
Two undifferentiated hESC lines (CHA15 and H9) were cultured as previously described [33]. Briefly, undifferentiated hESCs were grown on mitotically inactivated MEF cells in knockout DMEM/F-12 (1:1; Invitrogen, Carlsbad, CA) medium supplemented with 20% serum replacement (Invitrogen) and basic hESC medium components, including 1 mM
Feeder-free culture of enzymatically dissociated hESCs
The initial experiments involving feeder-free culturing of hESCs were all performed on a BD Matrigel™ hESC-qualified matrix (BD Biosciences, Bedford, MA). Before the hESCs were cultured, culture dishes were coated with BD Matrigel™ hESC-qualified matrix diluted in DMEM/F-12 (Invitrogen). To dissociate the hESC colonies into single hESCs, the ROCK inhibitor Y27632 (Tocris, Ellisville, MO) was added to the culture medium at a concentration of 10 μM for 18 h before detaching the cells from the feeder layer [14]. hESCs were first detached from the feeder layer using Dispase (Invitrogen), and the detached hESC colonies were washed with phosphate-buffered saline (PBS), which was followed by incubation in TrypLE™-Select (Invitrogen) at 37°C for 5 min. Dissociated single cells were seeded onto 12-well culture plates coated with BD Matrigel™ hESC-qualified matrix and cultured. Dissociated hESCs were cultured in knockout DMEM/F-12 (Invitrogen) supplemented with 1 mM
Toxin treatment
PTX from Bordetella pertussis (Tocris) was prepared in water containing 2.5 mg/mL bovine serum albumin (BSA; Sigma, St. Louis, MO) to create a stock solution with a concentration of 50 mg/mL PTX. CTX from Vibrio cholerae (Sigma) was prepared in water to create a stock solution with a concentration of 10 mg/mL CTX. hESCs were pretreated with PTX (final concentration of 200 ng/mL) or CTX (final concentration of 10 μg/mL) for 18 h in an hESC culture medium. The controls were treated in the same manner, except that water containing 2.5 mg/mL BSA was used instead of the toxin solutions. Fresh medium was added to feeder-free cultures of dissociated hESCs, which were maintained under the same culturing conditions throughout the colony formation assays.
Reagents and cellular sources
The reagents 2-methylthioadenosine diphosphate trisodium salt (meADP), adenosine 5′-triphosphate disodium salt (ATP), 2,4,6-trimethyl-N-(3-(trifluoromethyl)phenyl)benzene sulfonamide (m-3M3FBS), 1,2-Bis(2-aminophenoxy)ethane-N,N,N′,N′-tetraacetic acid tetrakis (BAPTA-AM), and Go6976 were purchased from Tocris, and 1-oleoyl-2-hydroxy-sn-glycero-3-phosphocholine (LPC) was purchased from Avanti Polar Lipids, Inc. (Alabaster, AL). Bisindolylmaleimide I (Bis) and safingol (Saf) were obtained from Calbiochem (La Jolla, CA). Go6976, Bis, Saf, and the PKCβ inhibitor were dissolved in DMSO. All of the control reactions were performed with the same DMSO volumes that were used in the drug treatments.
An HFF-1 cell line (SCRC-1041™), which originated from normal human skin fibroblast cells, was purchased from ATCC (
Live video imaging
Live video imaging was performed during the culturing of hESCs using the Tokai Hit incubation system for microscopes (Tokai Hit Co., Ltd., Shizuoka-ken, Japan). Movies were recorded using the Nikon ECLIPSE TS 100 microscope (Nikon, Chiyoda-ku, Japan) and analyzed using the i-solution software program (iMT technology, Inc., Seoul, Korea). Live imaging movie files were produced from a time series of images that were captured every minute for 72 h.
Alkaline phosphatase staining
To localize alkaline phosphatase (AP), hESC colonies were fixed with fresh 4% paraformaldehyde for 5 min at room temperature and then washed 3 times with PBS. After washing, AP was detected using an ES Cell Characterization Kit (Millipore, Bedford, MA) according to the manufacturer's instructions. Images were analyzed using an inverted fluorescence microscope (ECLIPSE TE2000; Nikon Instruments, Inc., Melville, NY).
Immunocytochemistry
Cells were fixed with 4% paraformaldehyde for 20 min and then permeabilized with 0.1% Triton X-100 in PBS (Sigma) for 5 min. After a treatment with 5% normal goat serum for 30 min, the cells were incubated with primary antibodies against one of the pluripotency markers [Oct-4; Santa Cruz Biotechnology, Inc., Santa Cruz, CA), E-cadherin (BD Biosciences), or PLCβ (Santa Cruz Biotechnology)] for 18 h at 4°C. The cells were then washed 3 times with PBS and incubated with Alexa Fluor 488- or 594-conjugated secondary antibodies (Invitrogen) for 1 h. After washing 3 times with Dulbecco's modified PBS, DAPI (Invitrogen) staining was performed for 10 min. The samples were immediately imaged using an LSM 510 META confocal microscope (Carl Zeiss, Inc., Oberkochen, Germany). Additional image analyses were performed using the ImageJ software program (
Intracellular calcium measurement
Intracellular calcium concentrations were determined by Grynkiewicz's method using fura-2/AM [35]. Briefly, prepared cells were incubated (under continuous stirring) with 3 μM fura-2/AM at 37°C for 50 min in a fresh serum-free knockout DMEM/F12 medium. The cells for each assay (2×106) were aliquoted into Locke's solution (154 mM NaCl, 5.6 mM KCl, 1.2 mM MgCl2, 5 mM HEPES, pH 7.3, 10 mM glucose, 2.2 mM CaCl2, and 0.2 mM EGTA). Fluorescence was measured at 500 nm using excitation wavelengths of 340 and 380 nm.
Total RNA extraction and reverse transcription–PCR
Total RNA extraction was performed using TRIzol reagent (Invitrogen) following the manufacturer's instructions. Briefly, total RNA was extracted with chloroform and precipitated with 80% (v/v) isopropanol. After the supernatant was removed, the RNA pellet was washed with 75% (v/v) ethanol, air-dried, and dissolved in RNase-free DEPC water (Bioneer, Daejeon, South Korea). After the samples were treated with DNase (Invitrogen), the RNA concentration was determined using a NanoDrop Spectrophotometer (NanoDrop Technologies, Wilmington, DE). The reverse transcription reactions were performed (with 1 μg of pure total RNA as a template) using AccuPower™ RT PreMix (Bioneer) following the manufacturer's instructions. The resulting cDNA was employed as the template in the PCR reactions using AccuPower™ Hotstart PCR premix (Bioneer) according to the manufacturer's instructions. The sequences of the PCR primer sets used in these amplifications can be found in the Supplementary Table S1; (Supplementary Data are available online at
Real-time quantitative PCR
cDNA was synthesized by following the methods described above. The real-time quantitative PCR primers are listed in Supplementary Table S2. The levels of target gene expression were determined by the comparative Ct method, whereby the target is normalized to the endogenous reference GAPDH. The Ct value represents the cycle number at which the fluorescence level reaches the threshold. ΔCt is determined by subtracting the Ct of the GAPDH control from the Ct of the target gene [ΔCt=Ct (target) − Ct (GAPDH)]. This relative value of the target to endogenous reference is described as the fold of GAPDH=2−ΔCt.
Statistical analyses
All of the experiments were performed at least 3 times to assess the reproducibility of the results. The quantitative data are expressed as the mean values±standard error of the mean. Student's paired t-tests or one-way ANOVAs were performed to analyze the statistical significance of each response variable. Prespecified comparisons between groups were performed (when appropriate) by post hoc testing using Tukey's method in the SPSS (Statistical Package for the Social Sciences) software program (version 17; Chicago, IL). P values of<0.05 were considered to be statistically significant. Histograms of the obtained data were generated using the GraphPad Prism program (version 5; SanDiego, CA).
Results
The colony-forming process of enzymatically dissociated single hESCs in a feeder-free system
To evaluate the process of colony formation of completely dissociated single hESCs, we set up a Matrigel™-based feeder-free culture system for 2 hESC lines (CHA15 and H9) and observed the process using live-cell imaging for 72 h. Briefly, hESCs were pretreated for 18 h with or without 10 μM Y27632 [14]. The hESCs were dissociated into single cells using the enzyme TrypLE™-Select. The dissociated hESCs were then spread on Matrigel-coated plates in the presence or absence of Y27632 in a maintenance medium after a pretreatment step (Fig. 1 and Supplementary Movies S1 and S2). Although cell attachment was observed for dissociated hESCs without Y27632 treatment 18 h after plating, the cells failed to form colonies within 72 h (Fig. 1A, B and Supplementary Movie S1). In contrast, the Y27632-treated dissociated hESCs attached and subsequently formed many large colonies (Fig. 1D, E, and Supplementary Movie S2). These colonies were primarily positive for alkaline phosphatase (AP) staining (Fig. 1F), unlike the colonies that were not treated with Y27632 (Fig. 1C).

The Rho-associated kinase (ROCK) inhibitor Y27632 markedly increases the efficiency of colony formation by enzymatically dissociated human embryonic stem cells (hESCs).
As demonstrated by live image analysis, colony formation of Y27632-treated single hESCs was composed of 4 stages (Fig. 1G; attachment, migration, aggregation, and colony formation), and most of the attached cells migrated and aggregated with neighboring cells (Fig. 1G, white circle indicates aggregation area). Interestingly, the aggregation of ∼10 single hESCs formed colonies and subsequently initiated cell division in unison, while nonaggregated single hESCs did not exhibit this behavior (Fig. 1G, Supplementary Fig. S1 and Supplementary Movie S3). From these observations, aggregation is required to commence colony formation of dissociated single hESCs. These results show that enzymatically dissociated single hESCs require contact with neighboring cells to form pluripotent colonies, and Y27632 guarantees the survival of dissociated single hESCs, which allows them to initiate colony formation.
To further support the importance of aggregation for colony formation, we plated single hESCs at 3 different cell densities. At 2.5×104 cells/well (0.63×104/cm2), no significant colony formation was observed (from 23.5±5.18 to 40.17±7.12 colonies for H9 and from 0.33±0.89 to 21.58±7.86 colonies for CHA15). However, cells that were plated at 5.0×104 cells/well (1.25×104/cm2) produced 3-fold more colonies when compared to those that were plated at 2.5×104 cells/well (from 29.42±7.23 to 133.83±16.88 colonies for H9 and from 2.75±2.30 to 151±23.50 colonies for CHA15). The highest amount of colony formation occurred at 1.0×105 cells/well (2.5×104/cm2) (from 54.58±6.67 to 175.42±16.78 colonies for H9 and from 8.91±3.26 to 197.25±12.86 colonies for CHA15), indicating that high cell density increased colony formation (Fig. 1H, I).
Inhibition of Gi signaling with PTX causes dissociated hESCs to form more condensed colonies
Single hESCs induced by Y27632 were found to form colonies 72 h after plating; they adopted a widespread, thin-layered morphology with bridge-like interconnections between neighboring colonies (Fig. 1E). The ROCK inhibitor Y27632 promotes the survival of dissociated single hESCs and allows them to initiate colony formation. However, the colonies formed by Y27632 have different morphologies from naïve hESC colonies cultured on feeder cells.
In an attempt to identify the major factors affecting colony formation, we focused on the role of Gi-coupled GPCRs with respect to cell physiology [26,29], because they represent a key signaling component involved in cell migration/aggregation. To study the role of GPCR signaling in this system, we pretreated hESCs with PTX or CTX, which irreversibly and specifically modulate Gi or Gs signaling, respectively. We then assessed colony formation using live video imaging and confocal microscopy. Pluripotent hESC colonies were pretreated with Y27632 and then treated with either PTX or CTX for 18 h, which was followed by enzymatic digestion. The fully dissociated hESCs were plated with the same cell numbers (5.0×104 cells/well) on Matrigel™-coated plates in a maintenance medium under the same conditions that were used in the pretreatment step (Fig. 2). After 72 h, the PTX-treated group (Fig. 2A and Supplementary Movie S4) adopted a densely aggregated, organized morphology; the outward projections of the cells disappeared, and they formed smaller colonies compared to the other groups (Fig. 2B, C). The CTX-treated hESC colonies exhibited constitutively activated Gs signaling, and their morphology was not affected by the treatment (Fig. 2B); they appeared similar to the control Y27632-treated group (Fig. 2C).
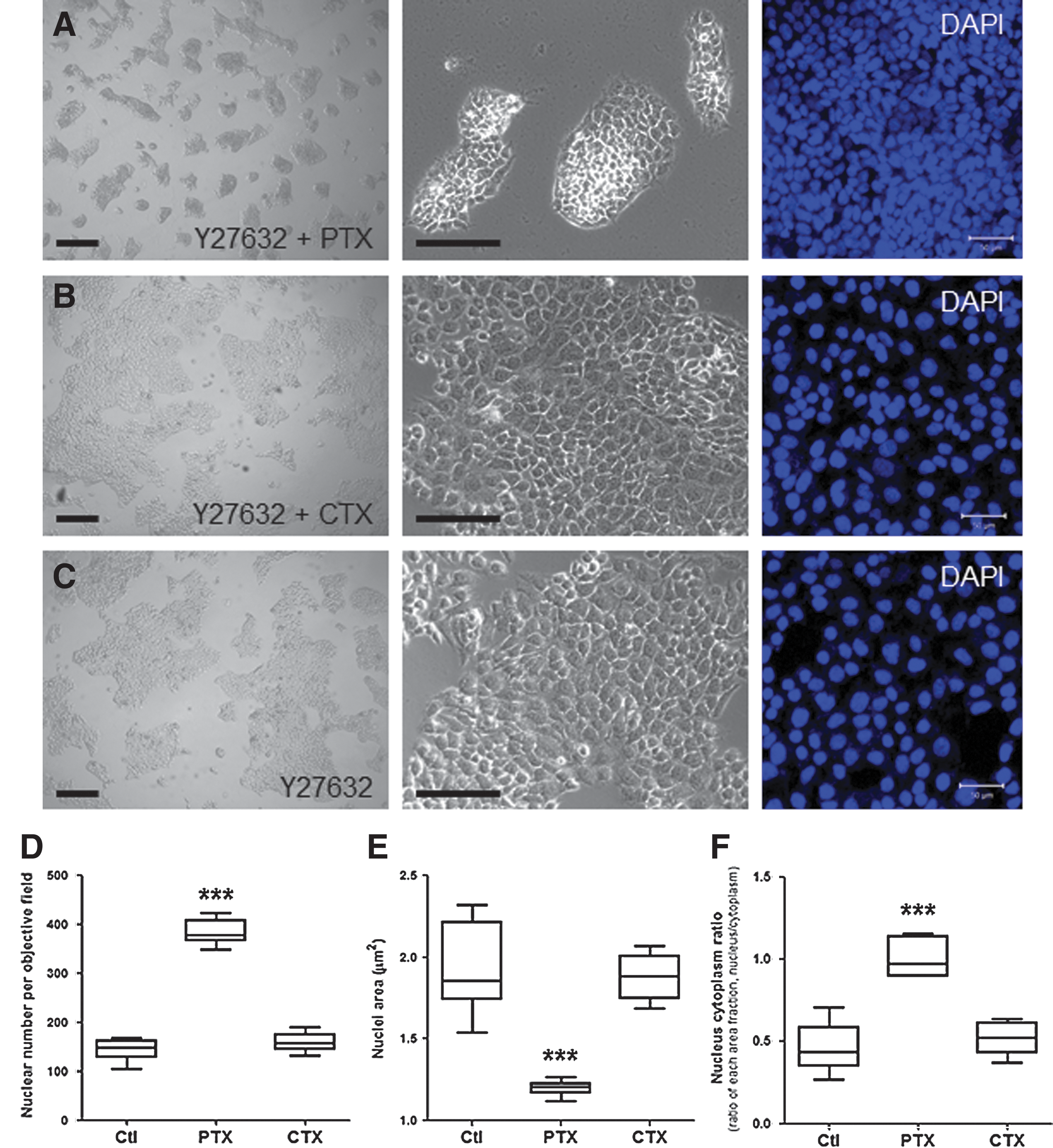
Inhibition of Gi signaling with pertussis toxin (PTX) enhances the organization of colonies formed by dissociated hESCs.
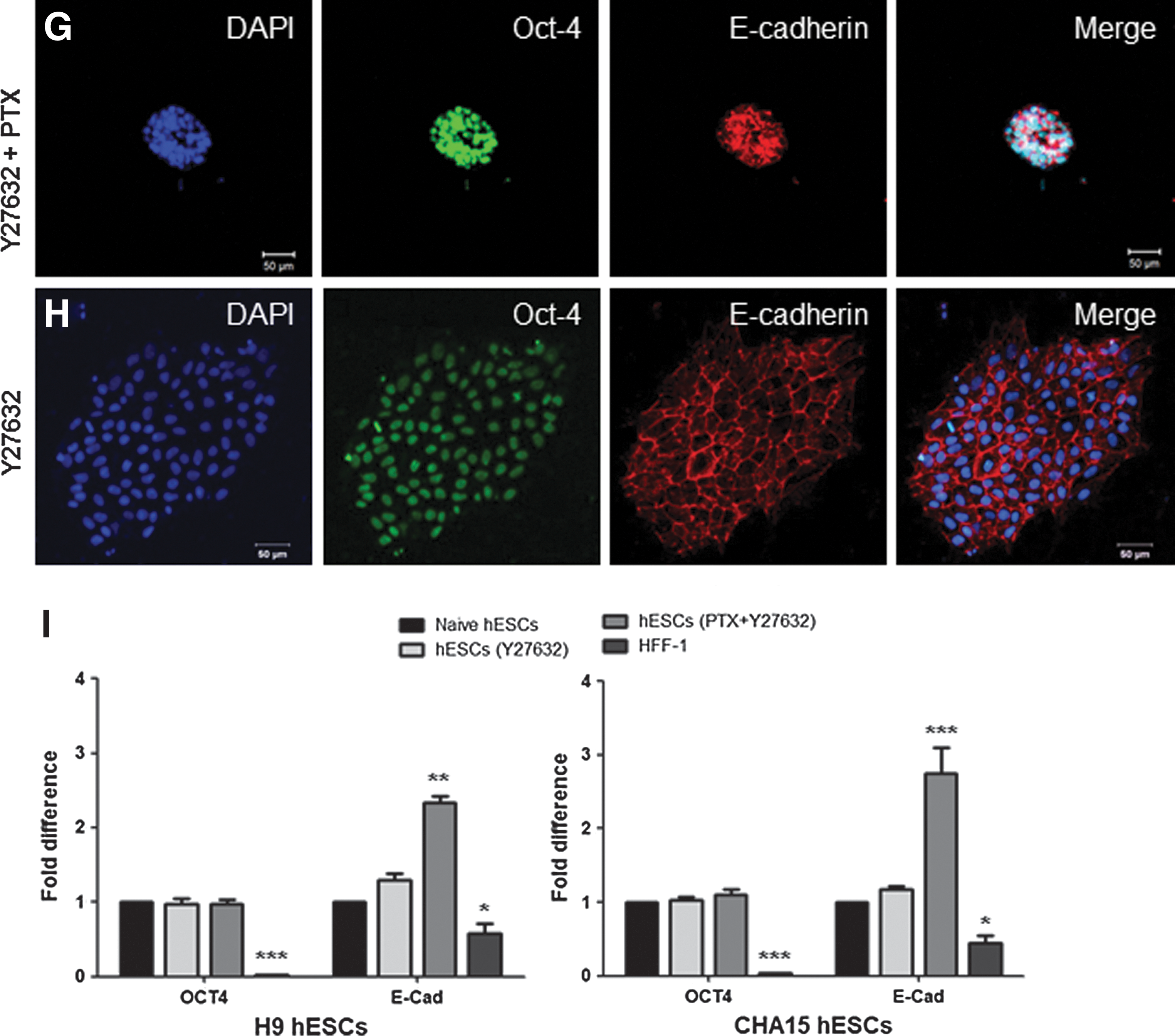
To investigate the effects of PTX on colony formation, we next assessed the cellular organization of the PTX-treated group by analyzing DAPI-stained nuclei using confocal microscopy and image analysis. The PTX-treated colonies (Fig. 2A, right panel) underwent reorganization and were significantly thicker and denser than the CTX-treated groups (Fig. 2B, right panel) and the control (Fig. 2C, right panel). Additionally, the colonies exhibited the largest number of nuclei per visual field (Fig. 2D), the smallest nuclear area (Fig. 2E), and the highest nuclear:cytoplasmic ratio (Fig. 2F) of the 3 groups. This result suggests that the inhibition of Gi signaling via PTX pretreatment disrupts endogenous mechanisms that maintain hESCs as characteristic flat monolayers. Therefore, the inhibition of Gi signaling with PTX consistently altered the morphology and organization of the hESC colonies, resulting in smaller, more compact colonies. Furthermore, we confirmed that the PTX treatment did not affect the pluripotency of hESCs (Fig. 2G–I and Supplementary Fig. S2) when immunocytochemistry was used to detect the expression of the pluripotency markers Oct-4 and E-cadherin (Fig. 2G) compared to the Y27632-treated control group (Fig. 2H). Although no difference in Oct-4 expression was observed, E-cadherin expression in the PTX-treated group was more than 2 times more than that of the Y27632-treated group (Fig. 2I). In addition, no differences were detected in other pluripotency markers (Supplementary Fig. S2A), and the PTX-treated hESCs retained the competence to differentiate into all 3 germ layers in vitro (Supplementary Fig. S2B).
Abolishing Gi signaling with PTX enables dissociated human pluripotent stem cells to maintain pluripotent colonies in a continuous and stable manner
We next examined whether PTX influences the continuous, stable maintenance of pluripotent colonies formed by enzymatically dissociated hESCs. Over the course of 5 passages, PTX-treated cells demonstrated positive expression of pluripotency markers (Oct-4 and E-cadherin) and displayed densely organized nuclei (Fig. 3A). Furthermore, the PTX-treated hESC colonies retained the ability to form colonies after 5 passages (Fig. 3A). Despite continuous enzymatic dissociation, the PTX-treated group generated stable AP-positive colonies with little loss of colony numbers compared to the Y27632-treated control group (Fig. 3C). In contrast, the expression levels of Oct-4 and E-cadherin decreased in the Y27632-treated cells (Fig. 3B), and we were unable to maintain colonies after 3 passages (Fig.3C). The ability of dissociated hESCs to form colonies sharply declined in the Y27632-treated group (Fig. 3C). The colonies of dissociated single hESCs treated with PTX maintained a normal karyotype over the course of 5 passages (Fig. 3D). These results indicate that PTX causes the reorganization of dissociated hESC colonies and facilitates the continuous, stable maintenance of pluripotent colonies in an enzymatically dissociated feeder-free culture system.

PTX directly enhances passaging continuity in feeder-free cultures of dissociated hESCs.
Gi-mediated phospholipase Cβ signaling is absent in hESCs
Previous studies have demonstrated that the activation of ligand-specific Gi-coupled GPCRs initiates several intracellular signaling pathways, including increased intracellular calcium concentrations mediated by phospholipase Cβ (PLCβ) and PKC [26,29,30]. In general, both ATP and ADP can increase intracellular calcium concentrations by stimulating the purinergic Gi-coupled GPCRs, such as P2Y1, P2Y12, and P2Y13, in fibroblasts (such as HFF-1 human follicle fibroblasts) (Fig. 4A) [26,29]. In certain exceptional cases [(eg, in human peripheral blood-derived natural killer cells (hPB-NKs)], both ATP and ADP fail to increase intracellular calcium concentrations despite the expression of these purinergic receptors (Fig. 4B) [34]. In both cell types (HFF-1s and hPB-NKs), intracellular calcium concentrations are greatly increased by the direct PLC activator m-3M3FBS. However, both ligands (ATP and ADP) and the PLC activator failed to increase intracellular calcium concentrations in hESCs (Fig. 4C), even though the P2Y1, P2Y12, and P2Y13 purinergic Gi-coupled GPCRs were expressed (Fig. 4D). Additionally, we were unable to identify any additional ligands of Gi-coupled GPCRs that could increase intracellular calcium levels in hESCs (data not shown). We next determined whether PLCβ was expressed in hESCs and found that the PLC activator m-eM3FBS was unresponsive. Interestingly, although no response to m-3M3FBS was observed in the hESCs, PLCβ was expressed in the hESCs in an expression pattern similar to that of the HFF-1 cells (Fig. 4E, F). However, PLCβ expression in the HFF-1 cells was more than 3 times greater than in the hESCs (Supplementary Fig. S3).
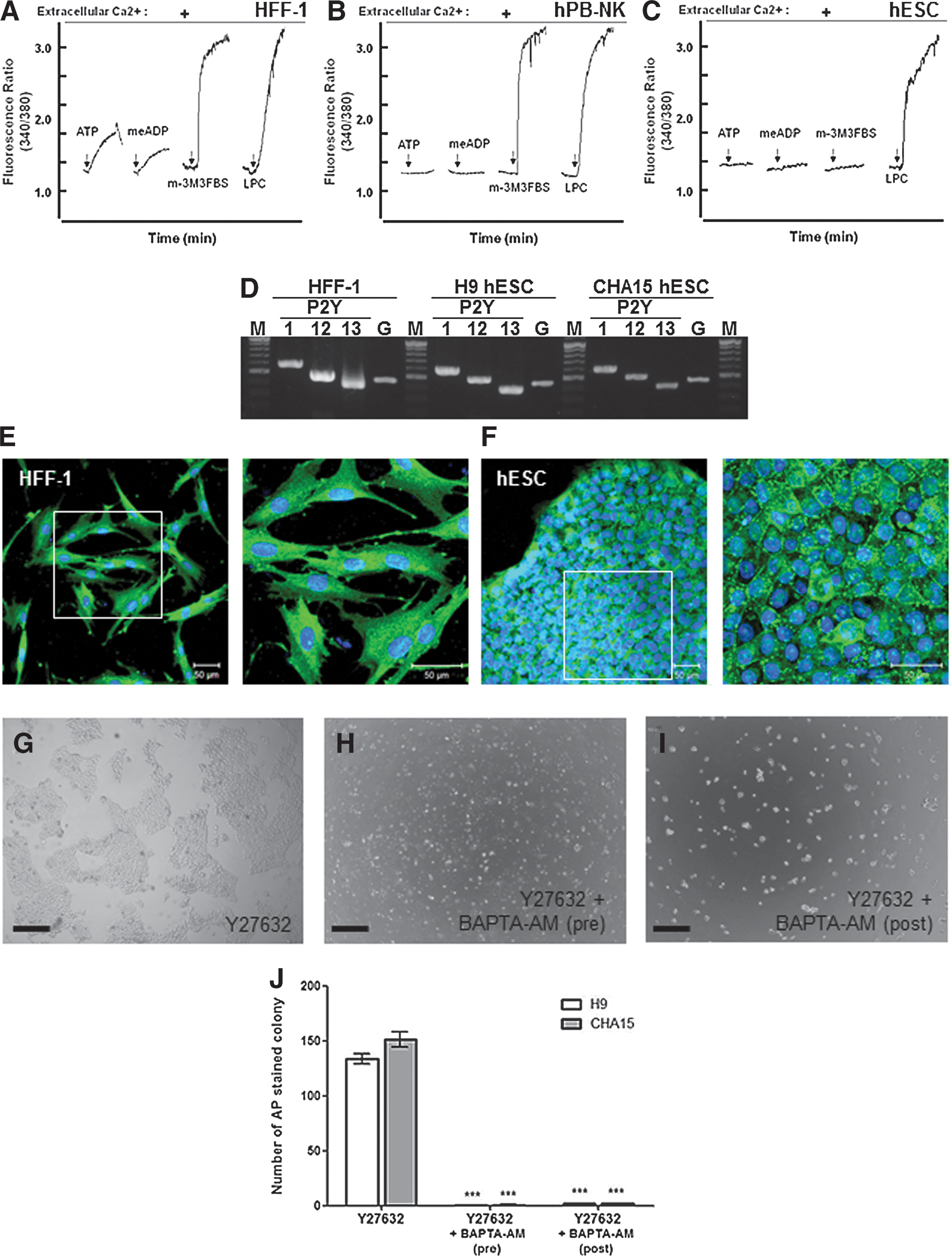
Gi-mediated phospholipase Cβ (PLCβ) signaling is absent in hESCs.
We also examined whether intracellular calcium is required for colony formation in dissociated human pluripotent stem cells. After pretreatment with Y27632 in the presence or absence of BAPTA-AM for 2 h, completely dissociated single hESCs were plated for 72 h (Fig. 4G, H). This study indicated that only the Y27632-treated cells produced multiple large colonies, which is consistent with our previous results (Fig. 4G). In contrast, BAPTA-AM-pretreated hESCs failed to attach to Matrigel-coated plates (Fig. 4H). Additionally, we tested the effects of the application of BAPTA-AM 18 h after Y27632-treated single cells were seeded; we found that the attached hESCs did not initiate cell migration or colony formation (Fig. 4I). The effects of the BAPTA-AM treatment were similar for all of the hESC types (Fig. 4J). These results indicate that intracellular calcium is a crucial factor for initiating colony formation in dissociated hESCs, but Gi-coupled GPCR signaling in hESCs does not utilize PLCβ-mediated intracellular calcium mobilization.
PTX enhances hESC colony organization through differential expression of PKC isotypes
We could not find evidence that PTX treatment causes intracellular calcium mobilization via PLCβ, which suggests that the primary driving forces in hESC colony formation are innate pluripotent properties, such as the differential expression of PKC isotypes. The PKC family consists of ∼11 isozymes [36], which are divided into 3 subfamilies, conventional, novel, and atypical, based on their second-messenger requirements [37]. To investigate how PTX affects the endogenous mechanism of colony formation in dissociated hESCs, we analyzed the expression levels of 11 PKC isotypes by reverse transcription–PCR. PKC is a downstream signaling mediator of Gi-coupled GPCR signaling (Fig. 5A). In hESCs, PKCγ, which is a member of the conventional PKC (cPKC) subfamily, PKCδ, PKCɛ, PKCη, and PKCθ, which are novel PKCs (nPKC), and PKCζ, PKC$$$$, and PKCμ, which are members of the atypical PKC (aPKC) subfamily, were expressed. However, PKCα and PKCβ1 in the cPKC subfamily, PKCθ in the nPKC subfamily, and PKCζ and PKC$$$$ of the aPKC subfamily were expressed in HFF-1 cells, indicating that the second-messenger requirements of hESCs differ from those of HFF-1 (Fig. 5A). The expression profiles of the PKC isotypes were not affected by treatment with either Y27632 or PTX (data not shown). To examine the effects of PTX in PKC inhibition as a potent colony organizer of dissociated single hESCs, we treated Y27632-treated single hESC cultures with PKC subtype inhibitors (Fig. 5B). After PKCα/β inhibition, the Go6976-treated group partially enhanced colony organization (Fig. 5B, upper right panel) compared to the Y27632-treated control group (Fig. 5B, upper left panel). However, PKCα inhibition led to a failure in colony formation for the Safingol-treated group (Saf) (Fig. 5B, lower left panel). The PKCβ inhibitor (Fig. 5B, lower center panel) and the broad-spectrum PKC inhibitor Bis (Fig. 5B, lower right panel) did not obviously affect the colony formation of dissociated single hESCs compared to the Y27632-treated control group (Fig. 5B, upper left panel). To analyze the effectiveness of the PKC inhibitors, we quantified the colony area ratio in an objective field for the PKC inhibitor-treated groups and compared it to the Y27632-treated and PTX-treated groups (Fig. 5C). The colony formation of the Go6976-treated group partially enhanced colony formation compared to the Y27632-only treated group (P<0.001, Fig. 5C) and the PTX-Y27632-treated group (P>0.05, Fig. 5C). However, the PKCβ inhibitor- and Bis-treated groups did not show the same pattern as the PTX-treated group (Fig. 5C, P>0.05). These results may suggest that the mechanism of PTX-induced hESC colony formation is related to the downregulation of PKC α/β activity.

The mechanism of hESC colony formation induced by PTX is related to the differential expression of protein kinase C (PKC) isotypes.
Discussion
The unique properties of hESCs suggest that various degenerative diseases may be successfully treated [5,38]. However, such prospects are hindered by the lack of reliable tools that are required to monitor many important aspects of in vitro culturing and differentiation. This study demonstrates that the colony formation capacity of enzymatically dissociated single hESCs is mediated by PTX-sensitive Gi-coupled GPCRs. Undifferentiated hESC colonies are discrete and present a radial morphology with a flat monolayer organization [1,39]. The absence of such characteristics in hESC cultures usually indicates the lack or loss of pluripotency [40]. Enzymatic dissociation of hESCs into single cells leads to significant decreases in hESC propagation due to the low efficiency of cellular attachment; this phenomenon is not typically associated with mouse embryonic stem cells (mESCs) [1,2]. In addition, dissociated hESCs undergo massive cell death, particularly after complete dissociation, and the efficiency of cloning dissociated hESCs is generally very low [12,13]. Thus, it has been more technically challenging to culture hESCs than mESCs with a dissociation method. The ability to perform this particular type of culturing is important for procedures such as clonal isolation after gene transfer and the induction of differentiation for the purpose of obtaining an improved understanding of somatic cell reprogramming.
hESC colonies are organized as flat monolayers of cells that adhere tightly to one another. However, in-depth studies of this type of cellular organization are currently unavailable [41]. One possible solution would be to use ROCK inhibitors, which render hESC colonies more resistant to enzymatic disassociation and increase survival in single-cell suspensions [14,19,20]. We first observed the colony-forming procedure of Y27632-mediated dissociated single hESCs. Our live imaging observations demonstrated that dissociated single hESCs progress through after 4 stages of colony formation: attachment, migration, aggregation, and colony formation. The ROCK inhibitor Y27632 guarantees the survival of dissociated, single, attached hESCs. The colony formation stages are mediated by the migration of unspecified individual cells (Fig. 1G and Supplementary Movie S2). However, colonies formed from single hESCs induced by Y27632 have a widespread thin-layered morphology unlike naïve hESC colonies. To study the mechanism of hESC colony formation, we initially seeded 5.0×104 cells/well (1.25×104/cm2) of dissociated single hESCs to observe colony formation. In this culture system, enzymatically dissociated single hESCs were cultured under feeder-free conditions without feeder-independent maintenance media, such as mTeSR1 (STEMCELL Technologies, Vancouver, BC, Canada).
We next investigated the improved aggregation activity of dissociated single hESCs for complete pluripotent colony formation. Using our new experimental culture system, we focused on Gi-coupled GPCRs, which play major roles in cell migration and morphogenesis and, in contrast to the action of Gq, G11, G12, and G13-coupled GPCRs, do not activate the Rho-ROCK axis [42,43]. The effector of the Gi-GPCR pathway is PLCβ, which catalyzes the cleavage of membrane-bound phosphatidylinositol 4,5-biphosphate (PIP2) into the second messengers inositol-(1,4,5)-trisphosphate (IP3) and diacylglycerol (DAG). IP3 activates IP3 receptors found in the membrane of the endoplasmic reticulum (ER) and elicits the release of calcium from the ER; DAG diffuses through the plasma membrane, where it may activate any membrane-localized form of the second serine/threonine kinase PKC [29]. In addition, many PKC isoforms are activated by increases in intracellular calcium levels. Both of these pathways can converge on each other to signal through the same secondary effectors [37].
Interestingly, Fig. 2 and Supplementary Movie S4 show that PTX, which blocks Gi-coupled GPCR signaling, enhances colony organization from dissociated single hESCs. This result was not observed in the group treated with Y27632 alone. The inhibition of Gi-coupled GPCR signaling using PTX generated smaller, denser colonies compared to those observed in the untreated groups. The colonies generated by the PTX-treated cells exhibited an increased number of nuclei per visual field, the smallest nuclear areas, and the highest nuclear:cytoplasmic ratio in comparison to the Y27632- or CTX-treated groups; these characteristics are hallmark characteristics of healthy hESC monolayers. In addition, the PTX treatment enabled the hESCs to maintain a continuous, stable pluripotent state (Fig. 2 and Supplementary Fig. S2), which was apparent because they consistently expressed 2 pluripotency markers (Oct-4 and E-cadherin) and maintained a normal karyotype up to passage 5 (Fig. 3). This result is in accordance with a previous report that PTX and Y27632 do not apparently affect the pluripotency of hESCs [44].
By analyzing intracellular calcium influx using specific ligands of Gi-coupled GPCRs and a PLC activator, we found that Gi-mediated PLCβ signaling is absent in hESCs even though PLCβ is expressed in these cells (Fig. 4). Apart from PLCβ signaling, intracellular calcium was still required to initiate colony formation by dissociated hESCs. These results suggest that PTX-sensitive colony organization is mediated by PKC signaling, another key downstream mediator of Gi-coupled GPCR signaling. We also found that PKCγ, novel PKCs, and atypical PKCs were expressed in hESCs, whereas PKCα, PKCβ1, and atypical PKCs were expressed in HFF-1 cells (Fig. 5A). We extended our analyses to the effects of some PKC isotype inhibitors on Y27632-induced single-hESC-colony formation. Go6976, a PKCα/β inhibitor, partially affected the colony organization of single hESCs. Unfortunately, a full characterization of the differential expression of the PKC isotypes during colony formation from dissociated hESCs is currently unavailable. According to a previous report, the inhibition of the PKCα and β isoforms of PKC with Go6976 induced rapid clustering of cultured carcinoma cells and the formation of an increased number of cell–cell junctions [45]. In addition, lysophosphatidic acid has been shown to stimulate PKCα, β, ɛ, and ζ in a PTX-sensitive pathway in vascular smooth muscle cells [46].
Taking all of these findings together, we suggest a new model for inducing colony formation by dissociated hESCs as summarized in Fig. 6. First, enzymatically dissociated single hESCs should be treated with the ROCK inhibitor Y27632 to ensure their survival. Next, because single hESCs initiate colony formation in an intracellular calcium-dependent manner and because single hESCs are highly mobile and form large, loose cell aggregates, PTX should be used to inhibit Gi signaling and ultimately restrict the mobility of these single hESCs. This inhibition induced consistent changes in aggregation and colony formation. The PTX treatment resulted in the development of denser colonies, which closely resembled those formed from feeder-based undifferentiated hESCs, and these colonies could be stably cultured, even during enzymatic dissociation. In summary, treatment with PTX enables hESC growth and differentiation under unfavorable culture conditions, such as after dissociation and in suspension. The improved cloning efficiency conferred by PTX treatment will likely be useful for revealing unknown characteristics of hESCs and other important cell lines. Future studies of the mode of action of PTX may shed new light on colony formation and organization of hESCs.
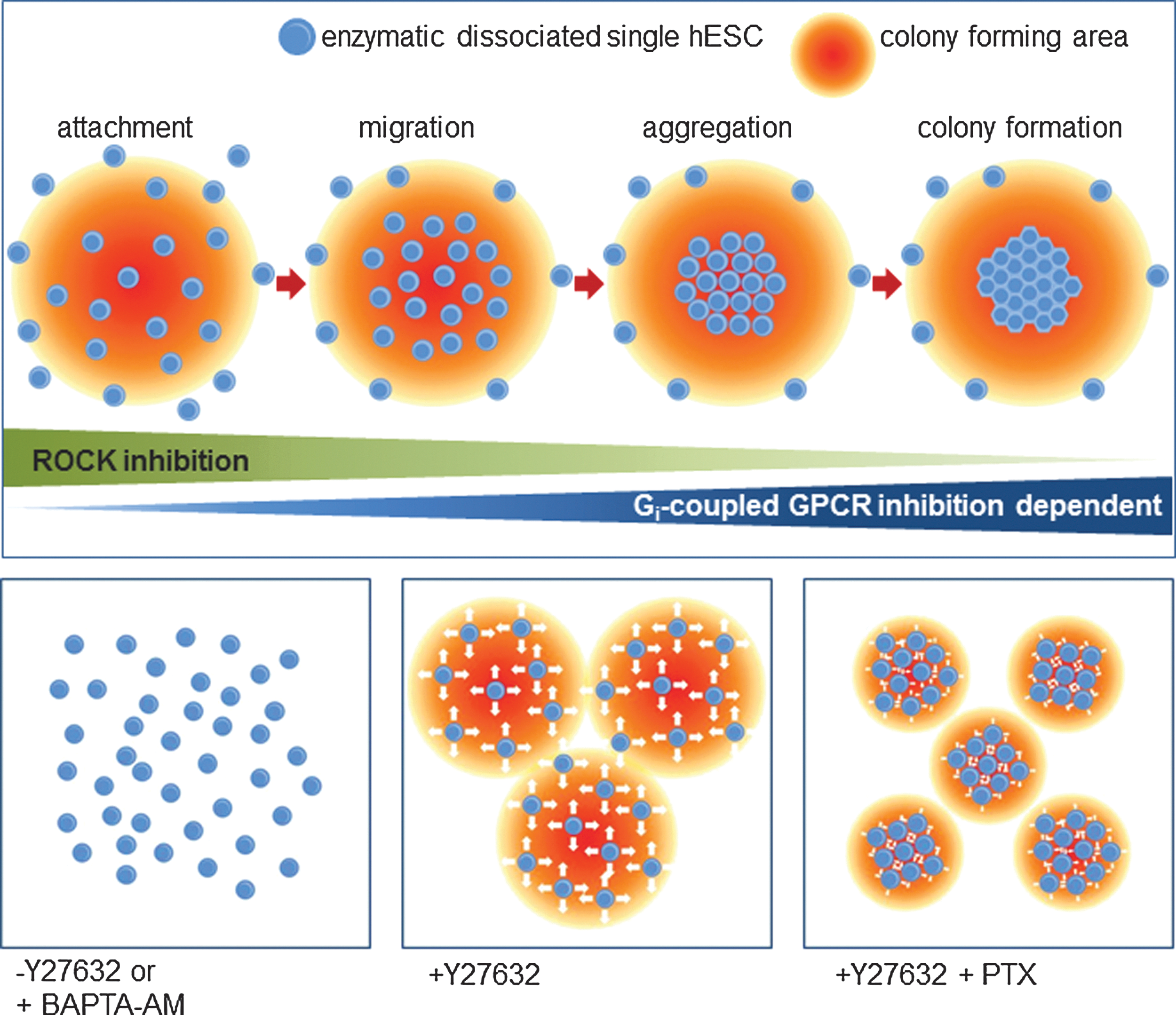
A schematic representation of colony formation by and organization of dissociated hESCs. The survival of enzymatically dissociated single hESCs is ensured using the Y27632 treatment. Single dissociated hESCs also initiate colony formation in an intracellular calcium-dependent manner. However, single hESCs are highly mobile, which results in the formation of large, loose cell aggregates. Inhibition of Gi signaling with PTX minimizes mobility, which alters the formation and organization of hESC colonies. Color images available online at
Footnotes
Acknowledgments
This research was supported by the Bio & Medical Technology Development Program of the National Research Foundation (NRF), which is funded by the Korean government (MEST) (no. 2012-0006107), and by the Industrial Core Technology Development Program, which is funded by the Korean government (MKE) (no. 10041913).
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.