
Abstract
Metalloproteinases (MMPs) and their endogenous inhibitors (TIMPs), produced in the brain by cells of non-neural and neural origin, including neural progenitors (NPs), are emerging as regulators of nervous system development and adult brain functions. In the present study, we explored whether MMP-2, MMP-9, and TIMP-2, abundantly produced in the brain, modulate NP developmental properties. We found that treatment of NPs, isolated from the murine fetal cerebral cortex or adult subventricular zone, with the clinically tested broad-spectrum MMP inhibitor Marimastat profoundly affected the NP differentiation fate. Marimastat treatment allowed for an enrichment of our cultures in neuronal cells, inducing NPs to generate higher percentage of neurons and a lower percentage of astrocytes, possibly affecting NP commitment. Consistently with its proneurogenic effect, Marimastat early downregulated the expression of Notch target genes, such as Hes1 and Hes5. MMP-2 and MMP-9 profiling on proliferating and differentiating NPs revealed that MMP-9 was not expressed under these conditions, whereas MMP-2 increased in the medium as pro-MMP-2 (72 kDa) during differentiation; its active form (62 kDa) was not detectable by gel zymography. MMP-2 silencing or administration of recombinant active MMP-2 demonstrated that MMP-2 does not affect NP neuronal differentiation, nor it is involved in the Marimastat proneurogenic effect. We also found that TIMP-2 is expressed in NPs and increases during late differentiation, mainly as a consequence of astrocyte generation. Endogenous TIMP-2 did not modulate NP neurogenic potential; however, the proneurogenic action of Marimastat was mediated by TIMP-2, as demonstrated by silencing experiments. In conclusion, our data exclude a major involvement of MMP-2 and MMP-9 in the regulation of basal NP differentiation, but highlight the ability of TIMP-2 to act as key effector of the proneurogenic response to an inducing stimulus such as Marimastat.
Introduction
N
MMP role in neurogenesis modulation has just begun to be unraveled. MMPs and their endogenous inhibitors TIMPs are expressed in neural stem progenitor cells [16,20,21], and the expression of MMP-2 and MMP-9 has been found increased in cell- and time-specific manners in the hippocampal neurogenic niche of adult monkeys after ischemia [22]. It has been reported that MMP inhibition significantly decreases neuroblast migration from the subventricular zone (SVZ) to the olfactory bulb under physiological conditions or in the damaged striatum after stroke [11,23,24]. TIMPs have been proposed to exert biological functions, including modulation of cell proliferation, migration, neurite outgrowth, differentiation, and cell death of both neuronal and glial cells, in MMP-dependent and independent manners. For example, TIMP-1 is a soluble protein secreted by activated astrocytes that exerts critical functions in the regulation of oligodendrocyte and astrocyte properties during development and after demyelinating injuries [25]. Cortical neurons from TIMP-3 knockout mice are more resistant against cell death induced by deprivation of glucose and oxygen in vitro, and these mice are more resistant to mild cerebral ischemia [26]. TIMP-2 expression increases during development, and it remains elevated in the adult brain. Recent studies have identified TIMP-2 as a key mediator of neurogenesis [27], since it acts as cell cycle-arresting molecule in vitro and promotes pheochromocytoma (PC12) cell differentiation, and since TIMP-2 knockout mice show delayed neuronal differentiation [28]. These findings strongly suggest that MMPs and TIMPs, produced by NSCs themselves and surrounding cells, can profoundly affect nervous system development and adult brain functions.
In the present study, we demonstrate that treatment of neural progenitors (NPs) with the broad-spectrum MMP inhibitor Marimastat strongly increases the generation of neuronal cells and concomitantly represses astrocyte, but not oligodendrocyte, differentiation by mechanisms that appear as independent on gelatinase inhibition. Specifically, by using specific siRNA and recombinant activated MMP-2, we exclude the involvement of MMP-2 in the regulation of early events of differentiation such as cell fate decision. We provide instead experimental evidence that TIMP-2 is implicated in the generation of neurons from NPs, and that it mediates the effects of Marimastat on these cells.
Materials and Methods
Cell cultures
NP cells used in the present study have been isolated from the E13.5 mouse cerebral cortex (eNPCTX) and from the adult mouse subventricular zone (aNP) [29,30]. NP lines were grown in the DMEM/F12 culture medium containing 1% N2 supplement, 20 ng/mL recombinant human epidermal growth factor (R&D) and 10 ng/mL recombinant human basic fibroblast growth factor (bFGF; R&D), penicillin (100 U/mL) and streptomycin (100 μg/mL; GIBCO), and 1% glutamine (GIBCO); this medium was termed the expansion medium. NP cell lines were plated on poly-ornithine- (10 μg/mL) and laminin- (5 μg/mL) coated flasks and split twice a week, and half medium was replaced every 2 days.
NP differentiation
NPs were plated on poly-ornithine/laminin-coated 6-well plates at 1×104 or 1.5×104 cells/cm2 density, in an expansion medium (defined as earlier). After 24 h, the expansion medium was replaced with the so-called bFGF differentiation medium consisting of a basal medium, supplemented with 10 ng/mL bFGF, 2% B27, and 1% N2 supplement. Cultures were differentiated under these conditions for 3 or 6 days. In the latter case, half of medium (or whole medium when the gamma-secretase inhibitor was included in the differentiation protocol; see below) was replaced with a fresh medium after 3 days. For longer differentiation, on the sixth day, the medium was replaced with a fresh medium containing 1% N2 supplement and 2% B27. To test the effects of the MMP inhibitor, the bFGF differentiation medium was supplemented with 1–2 or 5 μM Marimastat (Tocris) and added to the cells during the first 3 days of differentiation. At the end of treatment, some cultures were processed for the indicated analyses; parallel cultures were washed with phosphate-buffered saline (PBS) to remove residual Marimastat and further differentiated in the appropriate fresh medium for additional 3 or 6 days. In some experiments, Marimastat was added to the bFGF differentiation medium at the third day of differentiation. Marimastat was dissolved in dimethyl sulfoxide (DMSO).
Where indicated, the bFGF differentiation medium was supplemented with 20 ng/mL human recombinant brain-derived neurotrophic factor (BDNF; Peprotech) and 2 μM Marimastat. To test the effect of Notch signaling inhibition, eNPCTX was maintained in the bFGF differentiation medium supplemented with 0.5 μM N-[(3,5-difluorophenyl)acetyl]-
To generate almost pure astrocyte cell cultures, NPs were cultivated for 6 days in a basal medium containing 5% fetal bovine serum (FBS).
Immunocytochemistry
Immunocytochemical analysis was performed on NPs fixed with 4% PFA in PBS for 15 min at room temperature. After fixation, cultures were washed with PBS and then incubated in PBS containing 5% of the appropriate normal serum and 0.025% Triton X-100 (preincubation solution) for 1 h at room temperature. Subsequently, cultures were incubated at 4°C overnight in the preincubation solution containing the primary antibodies listed below in this section. Cells were washed 3 times with PBS and incubated for 1 h at room temperature in the appropriate incubating solution containing 10 μg/mL Hoechst 33342 and the appropriate biotinylated (Vector) and/or Cy3-conjugated (Jackson Immunoresearch) secondary antibodies at dilution 1:200. After rinsing with PBS, cells were incubated for 30 min at room temperature with Alexa-488-conjugated streptavidin (Molecular Probes) at dilution 1:200, rinsed, and coverslipped with a DAKO mounting medium (Dako).
The following primary antibodies were used: mouse monoclonal anti-βIII tubulin (1:500; Promega); rabbit polyclonal anti-GFAP (1:500; DAKO) rabbit polyclonal anti-TAU (1:100; Sigma); mouse monoclonal anti-O4 (1:100; Sigma); and goat polyclonal anti-doublecortin (DCX; 1:200; Santa Cruz).
The percentages of immunopositive cells in the different experimental conditions were determined by counting immunolabeled cells in at least 10–15 microscopy randomly chosen fields of 3 different cell cultures for each condition, in at least 3 independent experiments. An average of 100 cells per field was counted.
Reverse transcriptase–polymerase chain reaction
Total RNA was extracted from cells by TriReagent (Sigma), following the instructions provided by the manufacturer. Purified RNA was treated with RNase-free DNase I (Ambion) and quantified by spectrophotometric analysis. One microgram of RNA was reverse transcribed with M-MLV Reverse Transcriptase using random hexamers (Promega) in a final volume of 25 μL. Real-time polymerase chain reaction (PCR) was performed on the reverse transcription (RT) products with the SsoFast Evagreen Supermix (Biorad) in a Lightcycler apparatus (Biorad), following the manufacturer's instructions. Primer sequences for the examined genes are the following:
Mmp-2: for CAGGGAATGAGTACTGGGTCTATT, rev ACTCCAGTTAAAGGCAGCATCTAC [11]; Timp-1: for GAACGGAAATTTGCACATCA, rev ACCCCACAGCCAGCACTAT; Timp-2: for CGTTGGAGGAAAGAAGGAGTATC, rev GGCACAATGAAGTCACAGAGG [11]; Timp-3 for CAAAGTGGTGGGAAAGAAGC, rev GTGGGGCATCTTACTGAAGC; Timp-4 for CTTGCTATGCAGTGCCATGT, rev GGCTGGATGATGTCAACGTA; Mmp-9: for TGAGTCCGGCAGACAATCCT, rev TCTTGGTCTGCGGATCCTCA [31].
β-actin: for GCGCAAGTACTCTGTGTGGA, rev AAGGGTGTAAAACGCAGCTC; Hes1: for CGGCTTCAGCGAGTGCAT, rev CGGTGTTAACGCCCTCACA [32]. Hes5: for CCGGTGGTGGAGAAGATG, rev CTTTGCTGTGTTTCAGGTAGC; Hprt: for TGTTGTTGGATATGCCCTTG, rev TGGCAACATCAACAGGACTC.
Thermal cycling conditions comprised an initial step at 95°C for 3 s, followed by 40 cycles at 95°C for 5 s and 60°C annealing/extension temperature for 10 s. All samples were run in triplicate, and each well of PCR contained 20 μL as a final volume, including 2.5 μL of cDNA corresponding to 50 ng of total RNA, 0.2 μM forward primer, 0.2 μM reverse primer, and 10 μL SsoFast Evagreen Supermix (Biorad). The mRNA levels for Mmp-2, Mmp-9, and Timp-2 were compared between undifferentiated eNPCTX (proliferating cells) and eNPCTX differentiated with the bFGF differentiation medium or 5% FBS using the 2−ΔΔCT method for qRT-PCR data analysis. The mRNA levels for Mmp-2, Mmp-9, and Timp-2 were also compared between eNPCTX-derived astrocytes cultured in the presence or in the absence of 1 μg/mL lipopolysaccharide (LPS from E. coli 026:B6; Sigma) for 6 h.
Hes1 and Hes5 expression levels were assessed in eNPCTX cultured with the bFGF differentiation medium in the absence or in the presence of 2 μM Marimastat for 2, 4, and 6 h.
β-actin and Hprt were used as internal control genes. Amplification specificity was checked using a melting curve, following the manufacturer's instructions.
Gelatin zymography and Western blot
eNPCTX cell lines were cultured in an expansion medium or in a bFGF differentiation medium, and their conditioned media (CMs) were collected at different time points. Proteins in CMs were concentrated with a salting-out technique, using 85% ammonium sulfate saturation at pH 7.0 to obtain maximum protein precipitation [33]. Samples were thoroughly mixed, incubated at 4°C for 1 h, and then centrifuged at 10,000 g for 20 min. Precipitates were washed twice and resuspended in a nonreducing sample buffer (containing 62.5 mM Tris–HCl, pH 6.8, 35% glycerol, 2.5% Triton X-100, 4% SDS, 0.02% Brij-35, 0.002% NaN3, and 0.01% bromophenol blue). Gelatin zymography was carried out on 7.5% sodium–dodecyl sulfate polyacrylamide gel copolymerized with 0.2% 90 Bloom Type A gelatin from porcine skin [34]. After electrophoresis, gels were washed in 2.5% Triton X-100 and incubated for 24 h at 37°C in 50 mM Tris–HCl, pH 7.5, buffer containing 5 mM CaCl2, 100 mM NaCl, 1 mM ZnCl2, 0.02% Brij-35, 0.25% Triton X-100, and 0.002% NaN3. Gels were incubated in the presence of 5 mM EDTA and 2 mM 1,10-phenanthroline for inhibition studies through divalent ion chelation; selective MMP inhibition was carried out in the presence of 5 μM SB-3CT (4-henoxy-phenylsulfonyl-propylthiirane), a competitive MMP-2/MMP-9 broad inhibitor (Calbiochem); activation of zymogens was achieved with 2 mM p-aminophenyl-mercuric acetate. Staining was performed in 0.5% Coomassie BB R-250, and gels destained appropriately, until a clear band of gelatinolytic activity appeared against a blue background. Gelatinase standard (containing both MMP-2 and MMP-9 forms) was prepared by diluting healthy individual human blood with 15 volumes of nonreducing sample buffer [35].
To detect the amount of MMP-2 protein, samples (30 μg) were diluted in a sample buffer, boiled, subjected to 10% sodium dodecylsulfate–polyacrylamide gel electrophoresis, and electrotransferred to a 0.2-μm nitrocellulose membrane. The membrane was then blocked with 5% nonfat dry milk in Tris-buffered saline with 0.1% Tween-20. The MMP-2 antibody (2C1-1D12) used was a mouse monoclonal antibody from Zymed®, and for loading control, a rabbit polyclonal anti-actin antibody (Sigma) was utilized. Secondary anti-mouse horseradish peroxidase-conjugated antibody was from Bio-Rad, and ECL reagents were from Amersham. Densitometric analysis was performed with the ChemiDoc System (Bio-Rad).
To detect TIMP-2 protein, CMs were collected from proliferating NPs or NPs differentiated for 6 days. The whole medium was replaced with either a fresh expansion medium or a bFGF expansion medium (5 mL for each flask) 48 h before it was collected and concentrated (100×) in Vivaspin concentrators with 10,000-Da cut-off membrane (Sartorious), according to manufacturer's instructions. The corresponding cell cultures from which CMs were collected were lysated by a lysis buffer containing 20 mM Tris, pH 7.5, 150 mM NaCl, 1% Triton X-100, 0.1% Tween-20, 1% NP-40, and a 100×protease inhibitor cocktail (Sigma), and their protein content was determined by Bradford assay. Cell lysates and concentrated media were loaded onto 12% sodium dodecylsulfate–polyacrylamide gels to resolve proteins and then electrotransferred to a 0.45-μm nitrocellulose membrane. The mouse monoclonal anti-TIMP-2 antibody (Immunological Sciences) and the anti-GAPDH polyclonal antibodies (ABCAM) were used at the concentration of 1 μg/mL and 0.4 μg/mL, respectively. Anti-mouse and anti-rabbit IgG alkaline phosphatase-conjugated secondary antibodies (Sigma) were used at 1:10,000 dilution. Bands were stained with nitro blue tetrazolium in the presence of 5-bromo-4-chloro-3-indolyl-phosphate.
Electroporation of NPs and RNA interference
Mmp-2 silencing was obtained by using a specific pool of 4 different siRNAs validated and commercially available (ON-TARGET plus siRNA; Dharmacon). siRNAs were delivered into eNPCTX by electroporation according to the manufacturer's instruction (Nucleofector; Lonza). Briefly, 2–3×106 cells were resuspended in 100 μL of Mouse NSC Nucleofector Solution (Mouse Nucleofector Electroporation Kit; Lonza; VPG-104) containing 2 μg of a plasmid carrying the reporter gene GFP and subjected to an electric pulse. Under these experimental conditions, around 80% of the cells received the reporter plasmid. The same procedure was used to silence Mmp-2. eNPCTX were resuspended in 100 μL of electroporation mix containing either 2 μg of siMMP-2 or 2 μg of the ON-TARGET plus non-targeting siRNA (NT-siRNA; negative control). Immediately after electroporation, 4,000 cells/cm2 were plated on poly-ornithine-/laminin-coated plates in a prewarmed and carbon-dioxide-equilibrated complete culture medium and left in the incubator for 24 h. Cells were then rinsed with a basal medium and differentiated for 72 h in a neuronal differentiation medium supplemented with 2 μM Marimastat or DMSO. Mmp-2 gene expression and MMP-2 protein level and activity were determined 96 h after electroporation by either RT-PCR on cell lysates or by gel zymography on CMs.
Parallel cultures were also differentiated for additional 3 days, and their capacity to generate neurons after silencing was evaluated by immunocytochemistry.
Statistical analysis
Statistical analysis was performed using one-way ANOVA for multiple comparisons, and Bonferroni post hoc test or Student's t-test where indicated. P values≤0.05 were considered as significant.
Results
Effects of the MMP inhibitor Marimastat on NP survival and differentiation
To investigate the contribution of gelatinases in the regulation of NP properties, we used Marimastat, a broad-spectrum pharmacological inhibitor of MMPs clinically tested [7,36,37]. NP cell lines (eNPCTX) were isolated from the fetal brain cortex and expanded as adherent cultures according to the protocol described by Conti and colleagues [38]. We have previously demonstrated that under these culture conditions, virtually all eNPCTX express NSC markers (eg, nestin) and lack of spontaneous differentiation, indicating that these cultures are highly homogeneous and devoid of differentiated cells [30]. To evaluate the effects of the inhibitor on neuron generation, eNPCTX were differentiated in a bFGF differentiation medium (for medium composition see the Material and Methods section) supplemented with the solvent DMSO or with Marimastat (1, 2, and 5 μM, as chosen on the basis of previous studies) [39] and fixed after 3 days, or switched to a Marimastat-free differentiation medium for additional 3 days before fixing. Neuronal cells were identified according to morphological criteria and by immunodetection of the immature neuronal marker βIII tubulin. At 6 days, Marimastat treatment increased, in a dose-dependent manner, the percentage of βIII tubulin-positive cells with respect to the cells differentiated in the absence of the inhibitor (Fig. 1A, B). Similar results were obtained by assessing the expression of the neuronal marker microtubule-associated protein Tau (Fig. 1C). Despite the higher percentage of neurons, the average length of their main neuritic process, stained by the anti-Tau antibody, was decreased in Marimastat-treated cultures (Supplementary Fig. S1; Supplementary Data are available online at
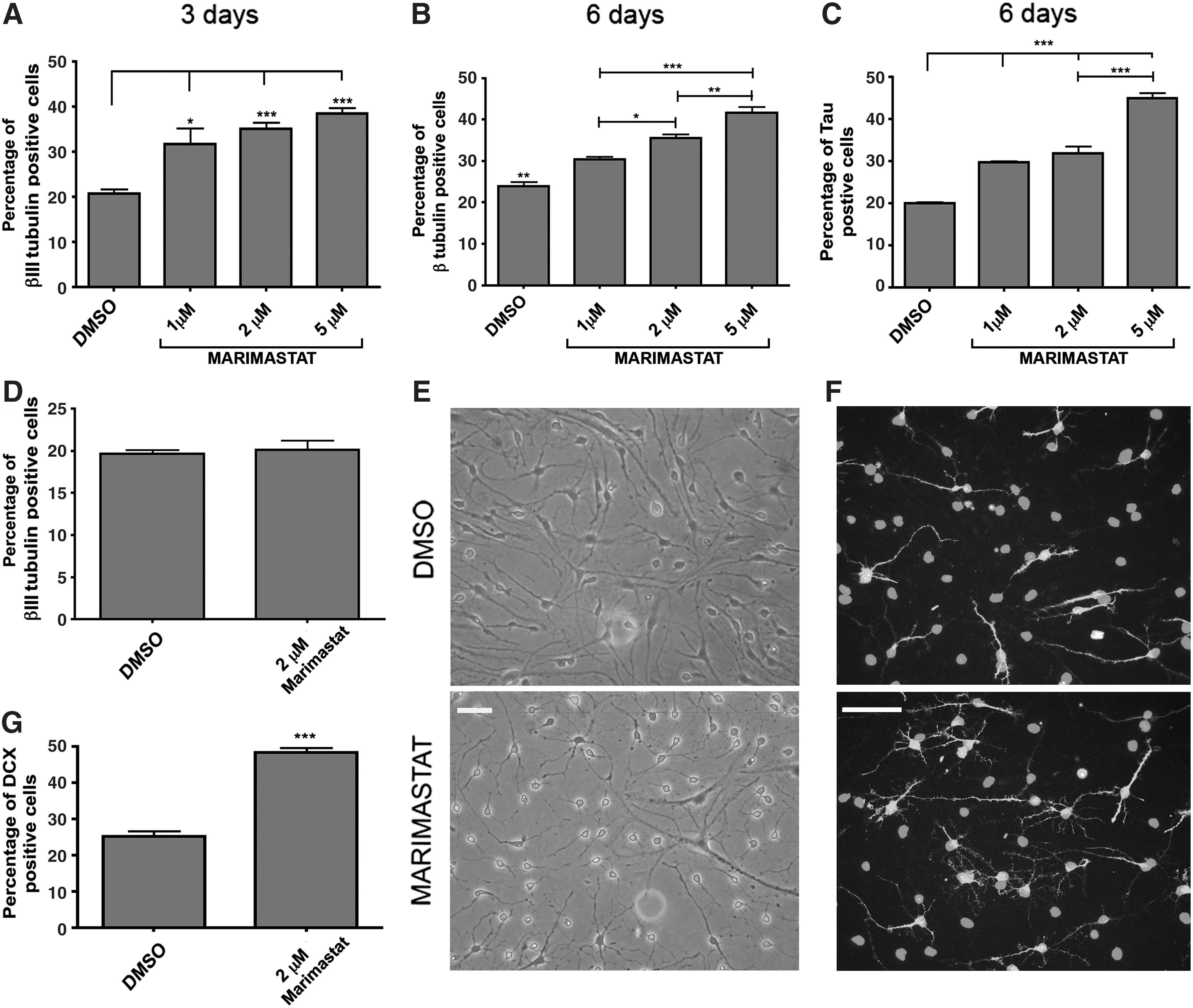
Effect of Marimastat on eNPCTX differentiation into neurons.
Establishing simple in vitro conditions to obtain highly enriched neuronal cultures is a critical requirement to make NPs as a valuable source for the generation of neurons and for the investigation on neuronal functions at the biochemical and molecular level. To increase the efficacy of Marimastat in differentiating eNPCTX into neurons, we adopted a slightly modified differentiation protocol. Fresh Marimastat-containing medium was replaced every day, and BDNF was added during the entire course of the differentiation protocol to increase viability of newly formed neurons. Under these conditions, the majority of cells acquired a neuronal morphology, and the percentage of doublecortin (DCX)-immunopositive cells doubled compared with control cultures after 6 days of differentiation (48%±0.63% vs. 25%±0.68% neuronal cells), indicating that these experimental conditions allowed for a relevant enrichment in neuronal cells (Fig. 1E–G).
The higher percentage of neurons found after Marimastat or Marimastat plus BDNF treatment could indicate either that Marimastat affects eNPCTX commitment or it rather accelerates the differentiation process. To discriminate between these hypotheses, the generation of neuronal and glial cells was evaluated at later time points. Because of the increased complexity in data interpretation due to BDNF addition, we performed all the subsequent experiments, aimed at dissecting Marimastat molecular mechanisms of action, in the absence of BDNF. After 9 days of differentiation, with Marimastat treatment limited to the first 3 days, we found that the percentage of βIII tubulin-positive cells increased from 19.3%±2.6% in control cultures to 37.6%±2.1% (Fig. 2A, B), whereas the percentage of GFAP-positive cells was reduced with respect to control cultures from 27.7%±2.3% to 17.7%±1%, respectively (Fig. 2A–C). Cells were also immunostained with an anti-O4 antibody, a marker for oligodendrocytes. As expected from the literature, the number of O4-positive cells was extremely low under control differentiation conditions (<1%; Fig. 2D); in the presence of Marimastat, the number of O4-positive cells remained negligible with respect to the others neural cell types. Altogether, these data suggest that Marimastat affects the early phases of the differentiation process, driving eNPCTX to differentiate into neurons and concomitantly reducing the generation of glial cells.
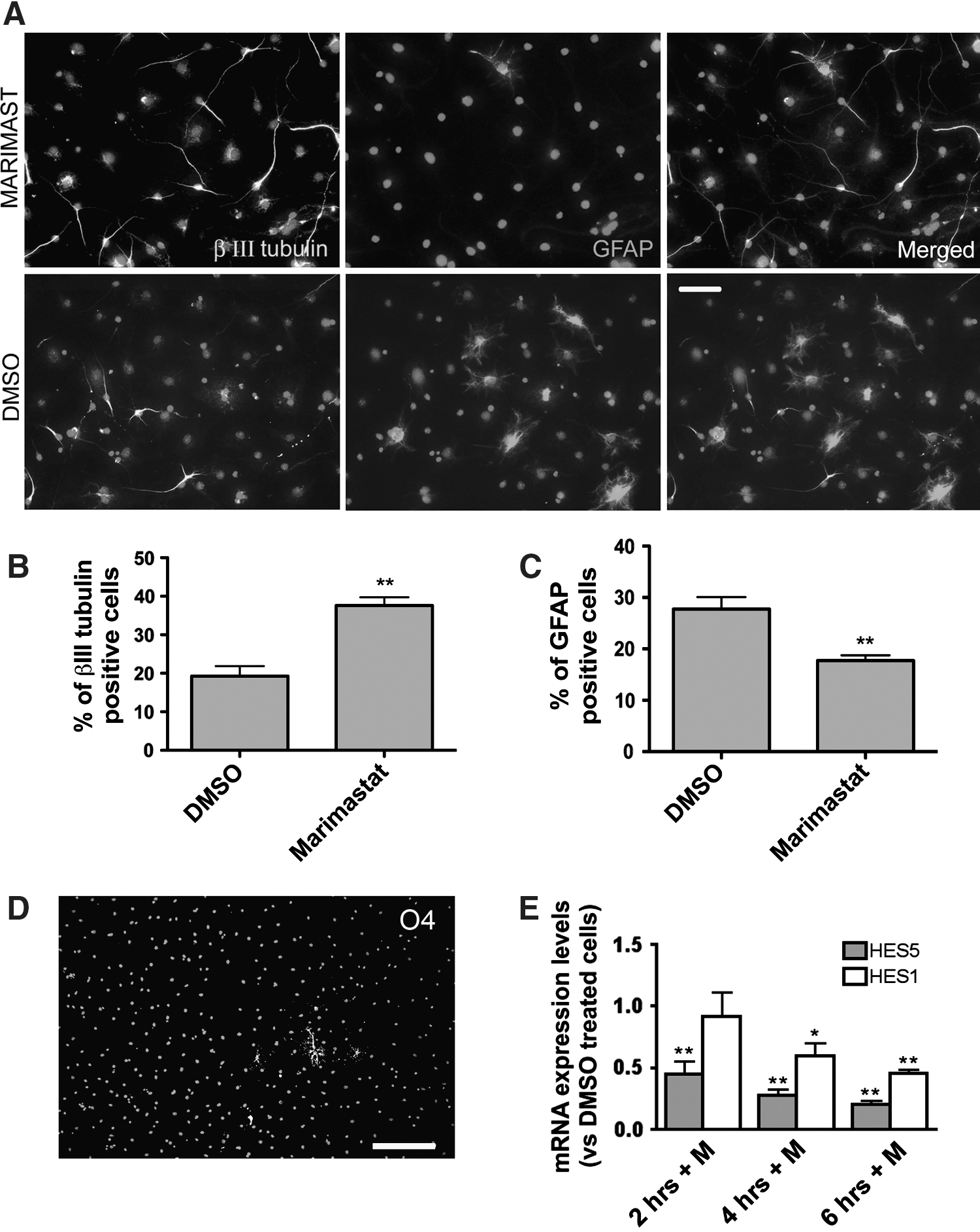
Evaluation of Marimastat effects on the neurogenic and gliogenic potential of eNPCTX differentiated for 9 days. eNPCTX were treated for 3 days with 2 μM Marimastat (or DMSO) and differentiated for additional 6 days.
To investigate the underlying molecular mechanism, we studied whether Marimastat affected the expression of HES genes, downstream targets of the Notch-signaling pathway, known to be involved in the maintenance and differentiation of NPs [40 –43]. Among the other functions, Notch signals control NP fate, inhibiting the neuronal and promoting the glial lineage. Interestingly, it has been recently demonstrated that the knockout of a specific disintegrin/MMP (namely ADAM10) affects Notch1 processing, increasing NP neuronal differentiation, and concomitantly repressing astrocyte formation [20]. We found that Hes-1 and Hes-5 expression was downregulated in eNPCTX cultured for 4 and 6 h in a differentiation medium supplemented with 2 μM Marimastat (Fig. 2E, F). To investigate whether Notch signaling inhibition in eNPCTX could indeed affect their differentiation, we used the gamma-secretase inhibitor DAPT. We first verified that in our cellular system, DAPT inhibited the canonical Notch signaling: in fact, the expression of Hes1 and Hes5 was significantly reduced after 4 and 6 h of treatment with DAPT (Supplementary Fig. S2A). eNPCTX maintained for 3 days in the bFGF differentiation medium supplemented with 0.5 μM DAPT generated a percentage of cells immunopositive for βIII tubulin comparable to that obtained with Marimastat alone. When Marimastat and DAPT were added simultaneously to the culture medium, the generation of neuronal cells was only slightly enhanced compared to the 2 drugs alone, further suggesting that similarly to DAPT Marimastat acts, at least in part, through the Notch pathway (Supplementary Fig. S2B).
To evaluate whether the enhanced neurogenic response to Marimastat was a unique characteristic of NPs derived from the fetal cerebral cortex or it was a mechanism shared with other NP types, we analyzed the effects of Marimastat on adult NPs derived from the mouse SVZ (aNP) as previously described [29,30]. All the experiments were conducted by using Marimastat at the dose of 2 μM. As found for eNPCTX, Marimastat increased the differentiation of immature aNPs into neurons (53%±7% and 31%±4% of neuronal cells in Marimastat-treated cultures and in control-differentiated cultures, respectively; Supplementary Fig. S2A, B). These data were reproduced also when performed on a clonally derived aNP line (referred as aNP1). Marimastat increased neuronal differentiation while decreasing astrocyte generation (Supplementary Fig. S2C). Same results were obtained on a different clone (aNP2; data not shown). Our data suggest that Marimastat acts on preserved cell-signaling pathways implicated in the regulation of neurogenesis in fetal and adult NPs.
Expression of endogenous gelatinases MMP-2 and MMP-9 and tissue inhibitor TIMP-2
To clarify the role played by MMP-2 and/or MMP-9 in eNPCTX differentiation, we investigated the expression profile of these MMPs at different time points. Qualitative RT-PCR analysis demonstrated that Mmp-2 is expressed in either proliferating eNPCTX or differentiating cells, whereas Mmp-9 was not expressed or expressed at very low levels compared with brain extracts from 2 months old mice used as a positive control (data not shown). To address whether Mmp-2 expression is modulated during differentiation, we performed a relative quantification of its specific mRNA by real-time RT-PCR analysis. Mmp-2 expression did not significantly change at any of the time points examined although a tendency to increase could be observed at 6 and 9 days of differentiation (Fig. 3A). As shown in Supplementary Fig. S2D, also in aNPs, Mmp-2 expression was not modified at these time points. Mmp-9 was barely detectable, and its expression was not modulated during differentiation of both eNPCTX (Fig. 3B) and aNPs (not shown), in accordance with previous data [31]. Since MMP-2 activation requires TIMP-2 [9], we investigated TIMP-2 expression profile, and we found that Timp-2 was progressively increased during differentiation of both eNPCTX and aNPs (Fig. 3C and Supplementary Fig. S3E). TIMP-2 increase after 6 days of differentiation was also confirmed by western blot analysis (Fig. 3D).

Expression of metalloproteinase (Mmp)-2, Mmp-9, and tissue inhibitors of metalloproteinase (Timp)-2 during eNPCTX differentiation by real-time reverse transcription–polymerase chain reaction (RT-PCR) or western blot analyses.
Since eNPCTX cultures differentiated under these conditions (bFGF differentiation medium) originate mixed neuronal and astrocyte cultures, with very few O4-positive oligodendrocytes, we aimed at identifying the cell population (neurons vs. astrocytes) mainly expressing Mmp-2 and Timp-2. To this purpose, eNPCTX differentiated for 6 days as above with the bFGF differentiation medium were compared to NPs differentiated with a medium containing 5% FBS, a culture condition inhibiting neuron generation and promoting astrocyte differentiation (almost 100% of the cells were positive for the astrocytic marker GFAP) (Fig. 4A). Real-time PCR analysis showed that Mmp-2 did not significantly increase under differentiating conditions, independently from the culture protocol used (Fig. 4B), and that Mmp-9 expression was barely detectable and did not increase upon astrocyte differentiation (Fig. 4C). Instead, Timp-2 levels in cultures enriched in astrocyte-like cells were significantly higher than in neuronal-/glial-mixed cultures (Fig. 4D), suggesting that astrocytes derived from eNPCTX are the major producers of TIMP-2 among the different neural cell types generated under these conditions. We also studied and compared the expression of Timp-1, Timp-3, and Timp-4 between proliferating undifferentiated NPs and astrocytes derived from eNPCTX after 6 days of differentiation with 5% serum. We found that Timp-1 was significantly downregulated, whereas Timp-3 expression was increased. Timp-4 expression remained unchanged (Fig. 4E). These data demonstrate that TIMPs are modulated during NP differentiation into astrocytes, and this could reflect their critical role in astrocyte differentiation and functions.

Characterization of Mmp-2, Mmp-9, and Timp-2 expression in different neural cell types by real-time RT-PCR.
To assess the functional properties of eNPCTX-derived astrocytes, the cultures were treated with 1 μg/mL LPS for 6 h, and their responses in terms of Mmp-2 and Timp-2 expression were analyzed. Stimulation with LPS did not change Mmp-2 and Timp-2 levels (Supplementary Fig. S4A, B), although eNPCTX-derived astrocytes were able to respond to LPS with increased expression of proinflammatory cytokines, such as Il-6 (347±18-fold increase vs. unstimulated eNPCTX-derived astrocytes). Our data suggest that eNPCTX-derived astrocytes acquire molecular and functional properties comparable to those displayed by primary astrocyte cultures [12,44 –46] and confirm NPs as a suitable cellular model to investigate on the role of MMPs and TIMPs in differentiation, maturation, and maintenance of both neuronal and astroglial cells.
MMP gelatinase activity and protein expression in eNPCTX and functional properties of eNPCTX treated with specific siRNA for Mmp-2 or with purified recombinant activated MMP-2 protein
MMP-2 is mainly regulated at the protein expression and activity levels. Therefore, the gelatinolytic activities of CMs collected from undifferentiated eNPCTX or collected over the course of their differentiation (at 3, 5, 7, and 10 days of differentiation) were analyzed by gelatin zymography. A band of increasing intensity, corresponding to the proMMP-2 (72 kDa), was observed throughout differentiation (Fig. 5A). These data were confirmed by western blotting analysis on cellular extracts demonstrating that MMP-2 was not expressed in proliferating cells and was upregulated upon differentiation (Fig. 5B). In agreement with the RT-PCR analysis, MMP-9 protein was not detectable in the conditioned medium.
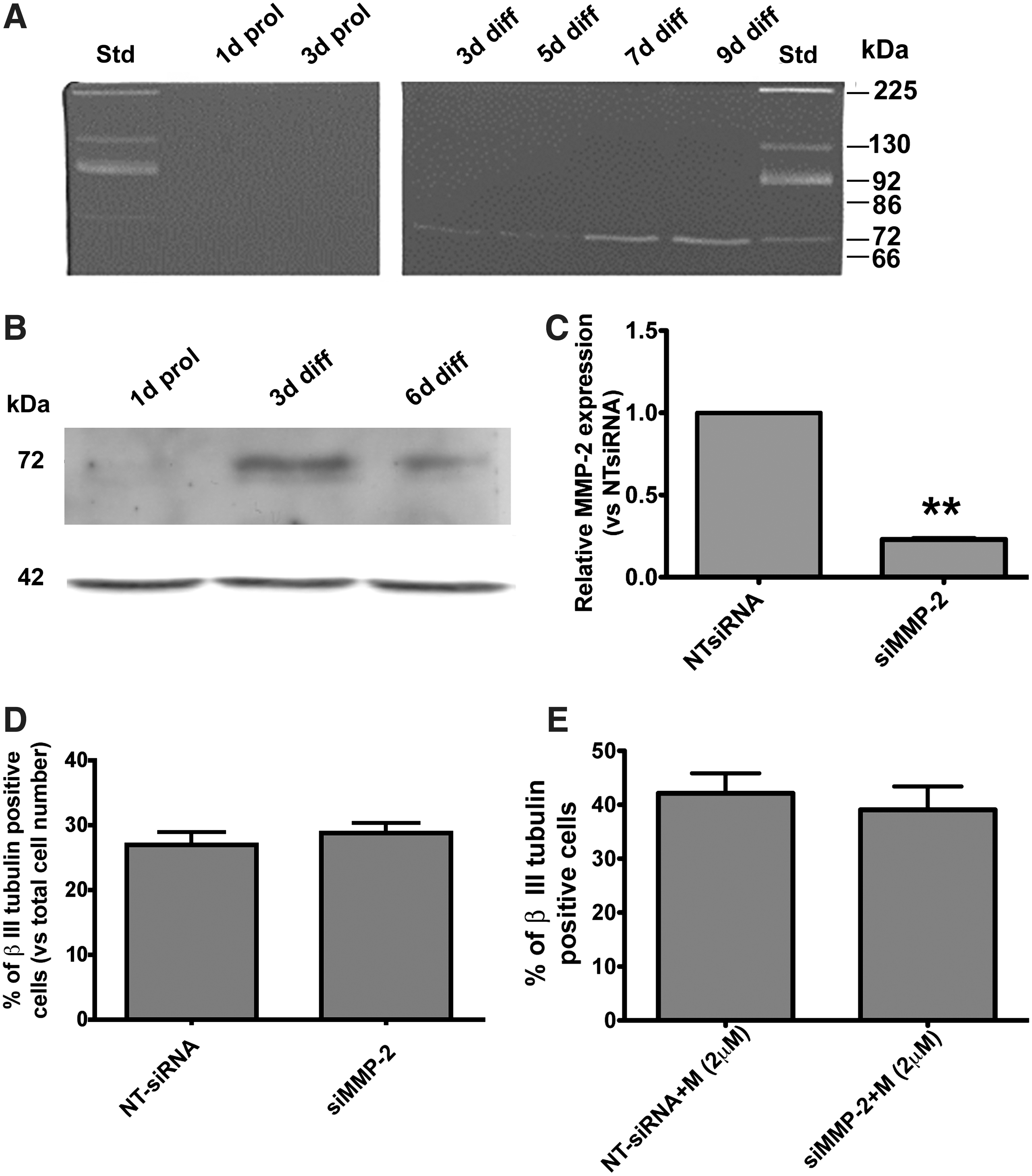
Gelatinases activity and Mmp-2 silencing.
Although activated MMP-2 (62 kDa) was never observed under our culture conditions, it is still possible that gel zymography is not sensitive enough to detect the presence of low levels of activated MMP-2. To further investigate on the role played by MMP-2, we used an approach based on specific siRNAs. eNPCTX electroporated with siRNAs against Mmp-2 (or appropriate non-targeting NT-siRNAs) were collected after 24 h, and Mmp-2 was analyzed by RT-PCR. Mmp-2 was significantly knocked down 24 h after silencing (60%±2.9%). Other eNPCTX cultures were shifted in a bFGF differentiation medium 24 h after electroporation and cultured for additional 2 days. These cultures showed 77%±1% reduction of Mmp-2 mRNA levels at 72 h from electroporation (Fig. 5C). Altogether, these data indicate that Mmp-2 was efficiently downregulated by siRNAs during the early phase of eNPCTX differentiation. eNPCTX cultures in which Mmp-2 had been silenced were differentiated for 6 days, fixed, and immunostained with anti-βIII tubulin antibody. Neuron generation was not affected upon Mmp-2 silencing (Fig. 5D).
Marimastat is a well-documented gelatinase inhibitor (MMP-2 and MMP-9 wide inhibitor), even though previous studies have demonstrated that it can, at least under certain experimental conditions, promote the activation of MMP-2 [39]. The observation that Mmp-2 knockdown did not significantly change the proportion of neuronal cells generated after treatment with Marimastat compared with control (NT-siRNA) cultures (Fig. 5E) suggests that Marimastat-mediated proMMP-2 activation, if occurring, is not responsible for the proneurogenic effect of the inhibitor. These data reinforce the hypothesis that MMP-2 activity is not implicated in regulating eNPCTX fate choice neither in the presence of the bFGF differentiation medium alone nor in combination with Marimastat. To further support our data, recombinant activated MMP-2 (rhMMP-2) was exogenously added into the bFGF differentiation medium at the final concentrations of 25 ng/mL (this concentration was chosen on the base of previous studies [47,48]); the effect of higher doses of rhMMP-2 (200 and 300 ng/mL) was also tested. Immunocytochemistry, performed after 6 days of differentiation, did not show any substantial modification in the neurogenic potential of eNPCTX (data not shown), suggesting that MMP-2 is not a major regulator of eNPCTX fate.
The proneurogenic effects of Marimastat require the expression of Timp-2
To investigate the possible link between the proneurogenic effect of Marimastat and TIMP-2, we knocked down endogenous Timp-2 by RNA interference. Based on a set of test experiments, we selected 125 nM as the most effective siRNA concentration to knockdown Timp-2 (not shown). Timp-2 was strongly downregulated 24 h after electroporation (82%±0.7% reduction compared with the control cultures). Furthermore, 3 days after silencing with the specific siRNA, Timp-2 reduction was >90%. Timp-2 mRNA in NT-siRNA-treated cells remained unchanged with respect to the mock-electroporated cells (Fig. 6A). eNPCTX in which TIMP-2 had been silenced did not display any significant change in the capacity of generating neurons when differentiated in the bFGF differentiation medium without Marimastat for 3 days (Fig. 6B). However, Timp-2 silencing reverted Marimastat proneurogenic activity, as eNPCTX treated with Marimastat generated a lower percentage of neurons after Timp-2 silencing as compared to NT-siRNA-electroporated cells (Fig. 6C).

Role of endogenous TIMP-2 in eNPCTX neuronal differentiation in the presence or in the absence of Marimastat.
To test the hypothesis that the proneurogenic effect of Marimastat could be mediated by the upregulation of Timp-2 above the endogenous levels, we performed real-time RT-PCR on eNPCTX differentiated for 4, 6, or 24 h. No differences in Timp-2 expression were detected at any of the analyzed time points (Fig. 6D).
Discussion
Neural stem/progenitor cell differentiation is regulated by means of both cell autonomous and external signals, which restrict temporarily and spatially the capacity of such cells to generate the different neural cell types. NP cell cultures derived from the developmental nervous system and adult brain have been successfully employed in the past years to unveil the contribution of soluble signals in the modulation of neurogenesis and gliogenesis. NPs are multipotent cells able to generate neurons, astrocytes, and oligodendrocytes, and they are a powerful cellular model to dissect neural differentiation and maturation processes at biochemical and molecular levels. In this regard, establishing simple in vitro conditions to obtain highly enriched neuronal (or glial) cultures represents an important goal [38,49 –51]. In the present article, we demonstrate that the broad-spectrum MMP inhibitor Marimastat is a potent regulator of the differentiation potential of NPs derived from the fetal and adult mouse nervous system. In particular, we show that Marimastat promotes neuronal differentiation allowing for substantial culture enrichment in neuronal cells. Moreover, Marimastat reduces astrocyte differentiation and does not substantially affect oligodendrocyte differentiation, at least in the absence of appropriate inducing signals for oligodendrocyte differentiation. The proneurogenic effect of Marimastat is independent on the modulation of MMP-2 and MMP-9 activities and requires the expression of the tissue inhibitor TIMP-2. We also observed that Marimastat can inhibit neurite outgrowth, possibly through an MMP-dependent mechanism. This observation, together with data from other groups, suggests that the use of MMP inhibitors could interfere with regenerative processes and synaptic plasticity. Although several data demonstrate that MMP inhibition stimulates neurogenesis from NPs and protects the nervous system from MMP-mediated detrimental effects, MMPs are also reported to exert beneficial effects in brain repair, promoting axonal growth and regeneration. The opportunity to use MMP inhibitors in brain phatologies needs therefore to be carefully evaluated and further investigated [52].
The use of MMP inhibitors has been adopted for the treatment of several pathologies, including brain diseases such as stroke. In the past years, the MMP inhibitor Marimastat has been largely used in in-vitro and in vivo studies and in clinical trials on patients affected by different types of tumors [7]. In the present study, Marimastat increased, in a dose-dependent manner, the percentage of neuronal cells in NPs derived from the fetal CTX and from adult SVZ. By using an optimized protocol, that is, by addition of BDNF plus Marimastat, we obtained around 50% of neuronal cells expressing specific neuronal markers and displaying a clear neuronal morphology. The interaction between MMP functions and BDNF signaling has been shown to be important for the regulation of synaptic plasticity and survival pathways mediated by BDNF, as the activation of MMPs can mediate proneurotrophin processing and, hence, Trk receptor activation [53,54]; in addition, increasing examples demonstrate that proteolytic processing of Trk receptors and coreceptor molecules by proteases, including metalloproteases, can be relevant for Trk downstream signaling, acting either as a self-limiting regulatory mechanism or as generator of fragments with signaling capabilities [55]. One possibility is that the enhancement of the proneurogenic effect of Marimastat in the presence of BDNF in NPs is mediated by the inhibition of TrkB cleavage, if any, by Marimastat; however, other mechanisms, independent of cleavage inhibition, remain to be explored.
It has been reported that Marimastat exerts, among others, antiproliferative effects on tumor cells [56,57], and it can promote eNPCTX differentiation favoring their cell cycle exit. In the present study, we show that 3 days of treatment with this drug induced an increase in the percentage of neurons compared to control cultures, and the effect persisted at 6 and 9 days of differentiation, as assessed by using different neuronal markers. At 9 days, we also found that the percentage of astrocytes was reduced by Marimastat treatment. Interestingly, adding Marimastat when NPs had started differentiating did not affect the ratio between neurons and astrocytes. All together, these observations indicate that Marimastat modifies the balance between neuronal and glial cells and suggest that Marimastat can modulate the fate choice of eNPCTX and aNPs. Although we cannot exclude that Marimastat differentially modulates the proliferation and/or survival of already committed neuronal and/or glial cells, the expression of Hes1 and Hes5, 2 genes supporting both NSC self-renewal and cell fate decision [40 –43], was downregulated few hours after Marimastat treatment.
To address whether the effect of Marimastat was due to the inhibition of its recognized targets, the gelatinases MMP-2 and MMP-9, the most abundant MMPs in the central nervous system, we investigated their expression and activity in eNPCTX and aNPs. Our data demonstrate that MMP-9 is not expressed neither in proliferating nor in differentiating conditions, whereas pro-MMP-2 protein was found in the medium of differentiating, but not in that of proliferating cells. Proteolytic pro-MMP-2 activation was never observed in the presence of the bFGF differentiation medium, strongly suggesting that MMP-2 is not directly responsible for NP differentiation, at least under our culture conditions. This view is further supported by the findings that neither the use of specific siRNAs to knockdown Mmp-2 expression nor the treatment of NPs with the recombinant purified MMP-2 affected NP differentiation. We conclude that MMP-2 does not modulate the early phases of the differentiation process in eNPCTX, and that the proneurogenic effect of Marimastat is not dependent on gelatinases, and other MMPs are likely to be implicated. In a recent study, Barkho and colleagues demonstrated that MMP-9 and MMP-3 are induced in NPs from adult SVZ differentiated in the presence of VEGF and SDF1 chemokines, and positively modulate neuroblast migration and neuronal differentiation [31]. The ability of Marimastat to inhibit other MMPs together with differences in cell cultures and conditions adopted could reconcile this discrepancy. In addition, our study shows that Marimastat can drive NP differentiation through an alternative mechanism involving TIMP-2. Tissue-specific inhibitors regulate MMP activity, and in particular, TIMP-2 is an important modulator of MMP-2 activation. Besides its classical MMP-2 inhibitory function, TIMP-2 can exert its effects in an MMP-independent manner [58], and it has been shown that TIMP-2 inhibits cell cycle progression and promotes neuronal differentiation [27,28]. In accordance with its antiproliferative role, we found that Timp-2 mRNA levels increased during differentiation, doubling after 3 days; moreover, Timp-2 levels further raised at 6 and 9 days of differentiation (up to 7-fold and 12-fold increase compared with proliferating NPs, respectively). Since cortical NPs sequentially generate neurons and then astrocytes both in vivo and in vitro [30,59 –61], we hypothesize that the progressive increment of Timp-2 is due to the maturation of neuronal cells and/or late appearance of glial cells. Consistently, the differentiation of NPs in the presence of 5% FBS for 6 days, a condition known to induce NPs to differentiate into astrocytes, almost abolished neuronal differentiation and increased Timp-2 expression (around 15 times compared with proliferating NPs). Hence, under serum culture conditions, the levels of Timp-2 were significantly higher than in neuronal-/glial-mixed cultures and undifferentiated NPs. We can reasonably assume that astrocyte-like cells generated from NPs in vitro are the major source of TIMP-2 among the different neural cell types studied, at least under our culture conditions.
TIMP-2 expression has been reported to regulate neuronal differentiation by affecting cell-cycle progression and neurite outgrowth, but it is not clear if TIMP-2 can affect NP cell fate too. To address this issue, we silenced Timp-2 and induced eNPCTX to differentiate by removing growth factors; we did not observe any change in the percentage of neuronal cells after 6 days. These data seem to exclude that TIMP-2 modulate early phases of differentiation process (ie, NP cell commitment to neurons). However, if TIMP-2 was knocked down and eNPCTX were induced to differentiate in the presence of Marimastat, the proneurogenic effect of this drug was lost. The underlying mechanism could involve the regulation of the matrix metalloproteinase MT1-MMP, whose activation by TIMP-2 elicits signaling pathways controlling cell proliferation and migration, independently by extracellular matrix proteolysis [62 –65]. Marimastat has been shown to reduce MT1-MMP autocatalytic turnover and to promote its accumulation [39,66]. In light of these observations and of our results demonstrating the dependence of the Marimastat proneurogenic function on TIMP-2 expression, it is possible to speculate the involvement of MT1-MMP in the regulation of eNPCTX fate. As far, whether and how MT1-MMP regulates NP properties and neurogenesis has not been explored. It remains to be established whether Marimastat exerts its action by modulating other unidentified MMPs. Transcriptome analysis on NPs from adult brain identified a small number of specific MMPs regulated during differentiation. In particular, MMP-24 is strongly regulated during differentiation and is also a direct target of the RE1-silencing transcription factor, a master regulatory gene of the neuronal differentiation (N. Buckley, personal communication). MMP-24 is a candidate for regulating NP differentiation, possibly by cooperating with TIMP-2 in mediating the proneurogenic effects of Marimastat. Interestingly, Marimastat can regulate MMP-24 activity [67].
In conclusion, we demonstrate that the broad-spectrum MMP inhibitor Marimastat is a potent modulator of NP cell fate decision, although the molecular mechanisms, which appear as gelatinases independent and TIMP-2 dependent, need further investigations. Unveiling these mechanisms could deepen our knowledge about the capability of MMPs and TIMPs in tuning NP properties both in vitro and in vivo. The emerging concept from the present and other studies is that different MMPs and TIMPs can specifically modulate NP functions (eg, inducing cell cycle exit, regulating neurite outgrowth, supporting cell migration, and promoting or repressing NP cell commitment and differentiation), thus making fundamental the development of appropriate tools to selectively target them to finely modulate neurogenesis and gliogenesis in a perspective of therapeutic intervention.
Footnotes
Acknowledgments
This work was supported by Fondazione Cenci Bolognetti and by Sapienza University of Rome to E.C. and by MIUR (PRIN 20088JEHW3_002) to S.B.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.