
Abstract
Mast cells play important roles in the pathogenesis of allergic diseases. They are generally classified into 2 phenotypically distinct populations: connective tissue-type mast cells (CTMCs) and mucosal-type mast cells (MMCs). The number of mast cells that can be obtained from tissues is limited, making it difficult to study the function of mast cells. Here, we report the generation and characterization of CTMC-like mast cells derived from mouse induced pluripotent stem (iPS) cells. iPS cell-derived mast cells (iPSMCs) were generated by the OP9 coculture method or embryoid body formation method. The number of Safranin O-positive cells, expression levels of CD81 protein and histidine decarboxylase mRNA, and protease activities were elevated in the iPSMCs differentiated by both methods as compared with those in bone marrow-derived mast cells (BMMCs). Electron microscopic analysis revealed that iPSMCs contained more granules than BMMCs. Degranulation was induced in iPSMCs after stimulation with cationic secretagogues or vancomycin. In addition, iPSMCs had the ability to respond to stimulation with the IgE/antigen complex in vitro and in vivo. Moreover, when iPSMCs generated on OP9 cells were cocultured with Swiss 3T3 fibroblasts, protease activities as maturation index were more elevated, demonstrating that mature mast cells were differentiated from iPS cells. iPSMCs can be used as an in vitro model of CTMCs to investigate their functions.
Introduction
Differentiation of both mouse and human embryonic stem (ES) cells into multiple hematopoietic lineages is now well established as a powerful tool for studying hematopoietic differentiation and lineage restriction, and for generating unlimited numbers of hematopoietic stem and progenitor cell populations for transplantation [9 –12]. ES or induced pluripotent stem (iPS) cells into hematopoietic cells have been differentiated by embryoid body (EB) formation or coculture with stromal cells, such as OP9 cells [13 –16]. By using these protocols, several groups have previously established methods to generate mast cells from mouse [17 –19], cynomolgus monkey [20], and human [21] ES cells. ES cell-derived mast cells could respond to stimulation with antigen and substance P by releasing histamine. However, in most cases, these cells do not develop the large granules and high levels of proteolytic enzymes characteristic of tissue mast cells.
In this study, we generated mast cells from mouse iPS cells (iPSMCs), and characterized them from the point of view of morphology, function, and gene expression. Our results showed that the iPSMCs that were differentiated by coculture with OP9 stromal cells or the EB formation method had characteristics similar to CTMCs. When iPSMCs that were generated on OP9 cells were cocultured with Swiss 3T3 fibroblasts, the iPSMCs exhibited a more functional phenotype.
Materials and Methods
Cell cultures
Two mouse iPS cell clones, 38C2 (a kind gift from Dr. S. Yamanaka, Kyoto University, Kyoto, Japan) [22] and 2A-EGFPTg-4F-01 [23], were used in the present study. These mouse iPS cells were routinely cultured in a leukemia inhibitory factor-containing ES cell medium (Specialty Media) on mytomycin C-treated mouse embryonic fibroblasts (MEFs; Specialty Media), and they were passaged every 2 days using 0.25% trypsin-EDTA (Invitrogen). OP9 stromal cells were cultured in an α-minimum essential medium (α-MEM: Sigma) supplemented with 20% fetal bovine serum (FBS), 2 mM
Generation of BMMCs
C57BL/6 mice were purchased from Nippon SLC. Bone marrow cells were prepared from the femurs and tibiae of mice. Cells were cultured in an RPMI 1640 medium containing 10% FBS, 1×NEAA, and 10 ng/mL murine IL-3 (R&D Systems). The culture medium was replaced with a fresh medium every 5 days. After 4 weeks of culture, we confirmed the cellular surface expression of both FcɛRI and c-kit (>95% positive).
Differentiation of iPS cells to mast cells
Before coculturing with OP9 cells or EB formation, mouse iPS cells were suspended in an ES cell medium and cultured on a culture dish at 37°C for 30 min to remove MEF layers. In the OP9 cell-mediated differentiation method, iPS cells were transferred onto OP9 cells in 6-well plates at a density of 1×104 cells per well. The induced cells were trypsinized on day 7, and 1×105 cells were seeded onto fresh OP9 cells with α-MEM supplemented with 20% FBS, 2 mM
In the EB-mediated differentiation method, iPS cell-derived EBs were generated by culturing iPS cells on a round-bottom low-cell-binding 96-well plate at 1×103 cells per well. iPS cell-derived EBs were collected on day 7, and were transferred to a Petri dish with Differentiation Medium I [Dulbecco's modified Eagle's medium containing 15% FBS, 1×NEAA, 2 mM
Transmission electron microscopy
BMMCs or iPSMCs were fixed with 2.5% glutaraldehyde in 0.1 M sodium phosphate buffer (pH 7.4), postfixed with 1% OsO4, dehydrated by a graded ethanol series, passed through QY-1 (Nisshin EM), and then embedded in Epon-812 (TAAB,). Ultrathin sections (0.06-μm thick) were cut with an ultramicrotome (Leica Microsystems), stained with uranyl acetate–lead citrate, and observed using an electron microscope (H-7650, HITACHI) at 80 kV.
Protease assay
BMMCs or iPSMCs were washed with phosphate-buffered saline (PBS), lysed in PBS containing 2 M NaCl/0.5% Triton X-100, and incubated for 30 min on ice. The lysate was centrifuged at 12,000 rpm for 30 min at 4°C. Activities of granule proteases in the resultant supernatants were measured using their specific chromogenic peptide substrates, such as S-2288 for tryptase (Sekisui medical) and M-2245 for carboxypeptidase A (CPA; Bachem) [24].
β-hexosaminidase release assay
β-hexosaminidase activity was measured as a marker of the granular fraction for evaluation of degranulation. Cells were washed with an HEPES buffer (137 mM NaCl, 20 mM HEPES, 5 mM
Coculture of mast cells with Swiss 3T3 fibroblasts
iPSMCs obtained after 28 days of culture with OP9 cells were cocultured with mitomycin C-treated Swiss 3T3 fibroblasts in the presence of 100 ng/mL SCF. BMMCs were also cocultured with Swiss 3T3 fibroblasts under the same conditions. The subculture was performed every 4 days. The cells were trypsinized and replated, and nonadherent cells were collected as mast cells and used for further analysis.
Mast cell reconstitution and induction of passive cutaneous anaphylaxis
BMMCs or iPSMCs (5×105 cells) were injected subcutaneously into the conjunctivae of mast cell-deficient Kit W-sh/W-sh mice. To elicit passive cutaneous anaphylaxis reactions, mice were injected subcutaneously into the conjunctiva with 75 ng anti-DNP IgE or saline. Then, 24 h after IgE injection, 100 μg DNP-HSA containing 2% Evan's blue dye was injected intravenously into mice. Thirty minutes later, the mice were killed, and their conjunctivae were excised. Evan's blue dye was extracted from conjunctivae with formamide, and the absorbance was measured at 610 nm.
Results
Generation of mast cells from mouse iPS cells
iPSMCs were generated by the OP9 coculture method or EB formation method as described in Supplementary Fig. S1 (Supplementary Data are available online at
Next, we performed the staining with May-Grunwald-Giemsa, toluidine blue, Alcian blue, and Safranin O solutions. May-Grunwald-Giemsa staining of the iPSMCs, which were differentiated by coculture with OP9 stromal cells or the EB formation method (Supplementary Fig. S1), revealed that induced mast cells gave rise to a uniform phenotype with rough basophilic granule-containing cells (Fig. 1A, upper). The granules in these cells showed a metachromatic staining pattern when stained with acid toluidine blue (Fig. 1A, middle). We then performed Alcian blue and Safranin O staining, by which mast cells are known to show a specific red color if they are CTMCs and a blue color if they are immature mast cells or MMCs [1]. While BMMCs were Alcian blue positive and Safranin O negative, iPSMCs were positive for both Alcian blue and Safranin O staining (Fig. 1A, [lower], B). Electron microscopic analysis revealed that the iPSMCs differentiated by either method contained more granules than BMMCs (Fig. 1C).
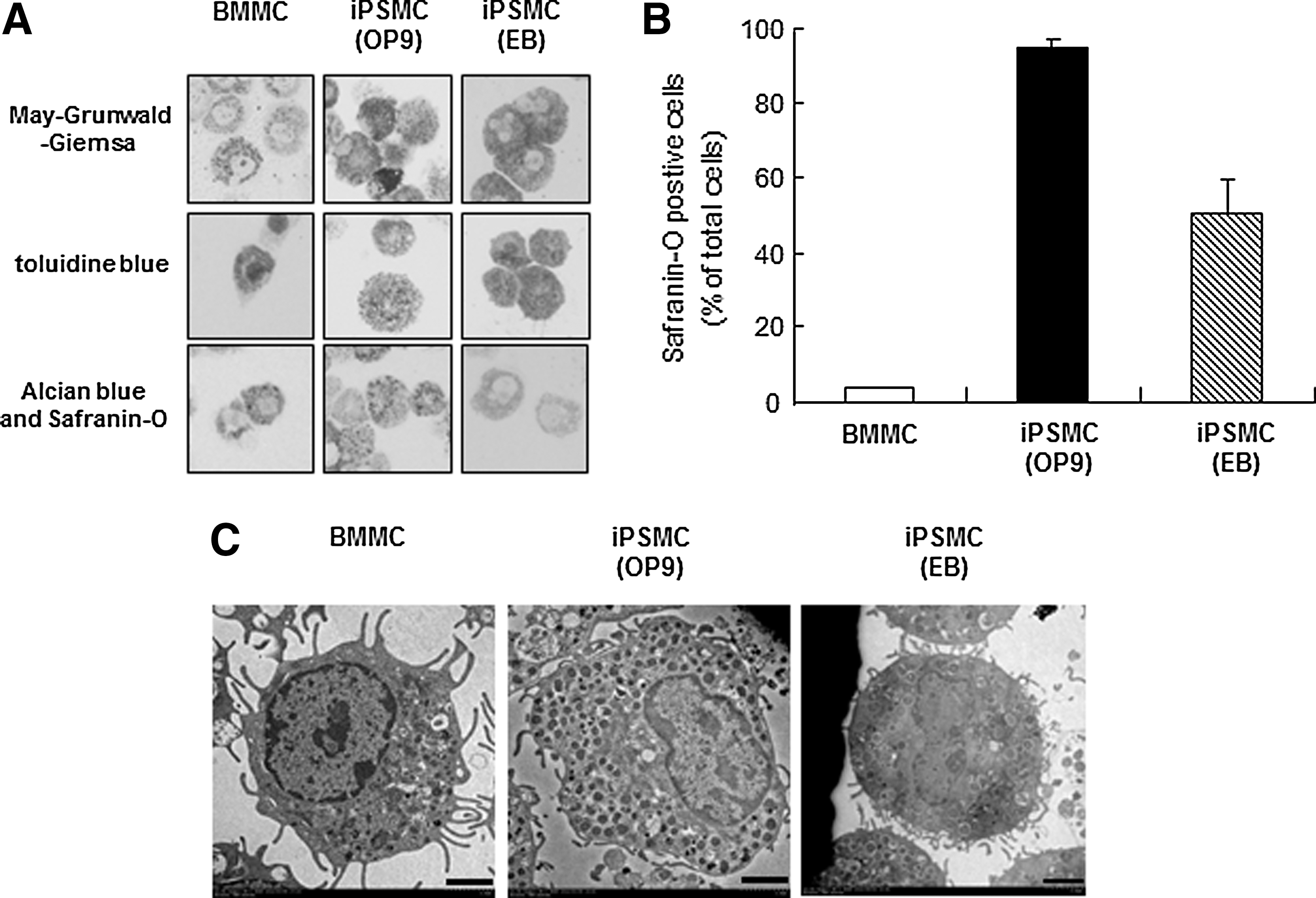
Morphological characterization of induced pluripotent stem cell-derived mast cells (iPSMCs).
Expression of high-affinity IgE receptor on iPSMCs
Mast cells are known to express c-kit and FcɛRI (high-affinity IgE receptor) [1]. We next performed flow cytometric analysis to examine the surface expression of c-kit and FcɛRI on iPSMCs. There was no significant difference in c-kit expression levels between iPSMCs and BMMCs (Fig. 2). In contrast, the FcɛRIα expression level was significantly lower in the iPSMCs that were generated by coculture with OP9 cells, compared with that in BMMCs. Both c-kit+FcɛRI+ and c-kit+FcɛRI− cells showed a granular phenotype by forward and side scatter (data not shown).
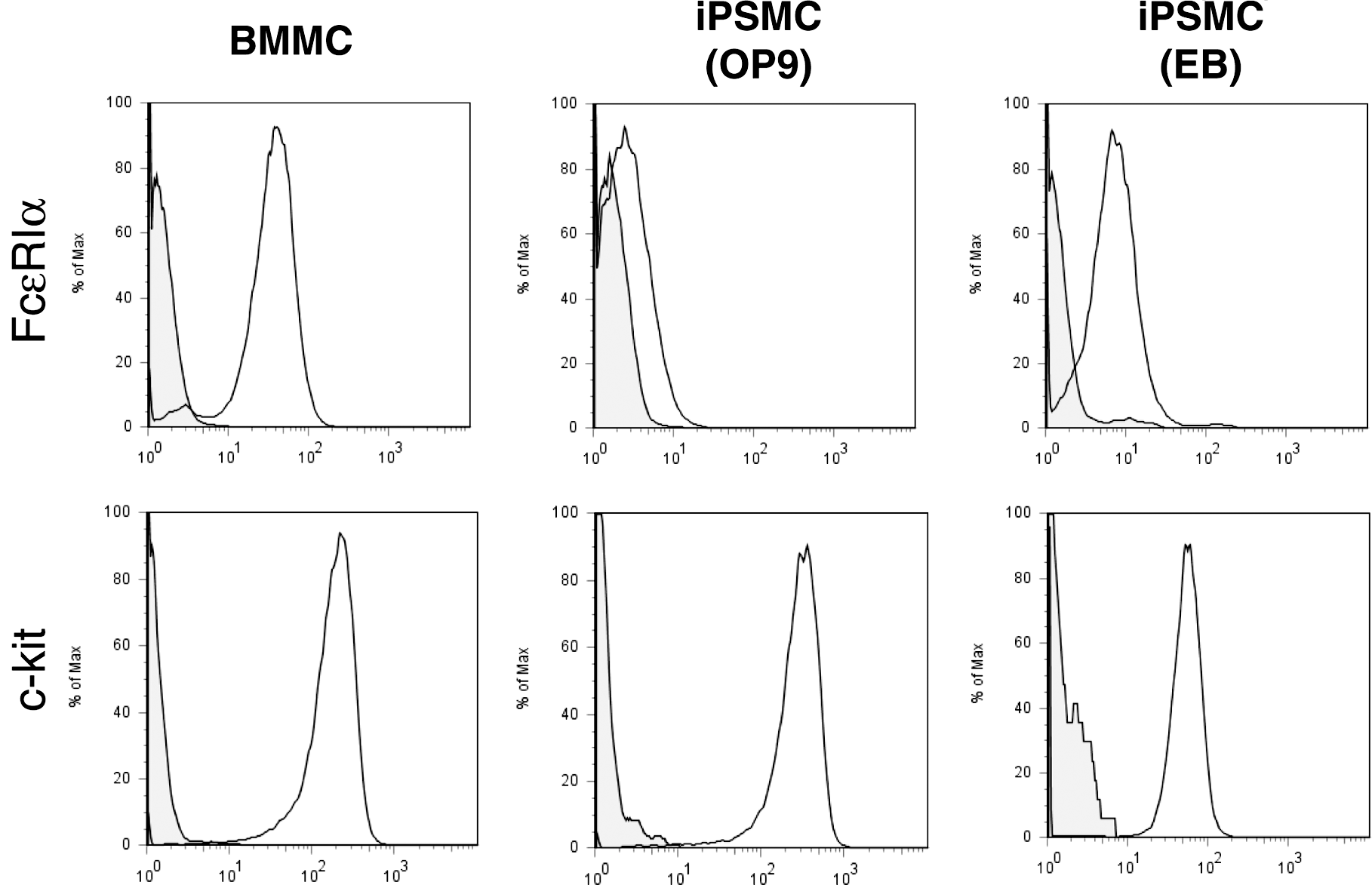
Flow cytometric analysis of FcɛRI and c-kit expression on iPSMCs. BMMCs and iPSMCs were stained with FITC-labeled anti-FcɛRI and PE-labeled anti-c-kit antibodies for 30 min on ice. Stained cells were washed, resuspended in 1% fetal bovine serum–phosphate-buffered saline (FBS-PBS), and analyzed by flow cytometry.
FcɛRI is a heterotrimer composed of one α-chain and 2 γ-chains or a heterotetramer composed of one β-chain and 2 γ-chains. To evaluate the expression of each FcɛRI subunit in iPSMCs, we analyzed mRNA expression levels by reverse transcription and quantitative polymerase chain reaction (RT-PCR). As shown in Supplementary Fig. S2, the expression levels of the mRNAs encoding the FcɛRIα, FcɛRIβ, and FcɛRIγ chains were reduced in the iPSMCs differentiated by either method as compared with the levels in BMMCs.
Phenotypic differences between iPSMCs and BMMCs
To further compare the degree of mast cell differentiation, we measured the tryptase and CPA activities in iPSMCs. The tryptase and CPA activities were elevated in the iPSMCs derived from either method as compared with those in BMMCs (Fig. 3).
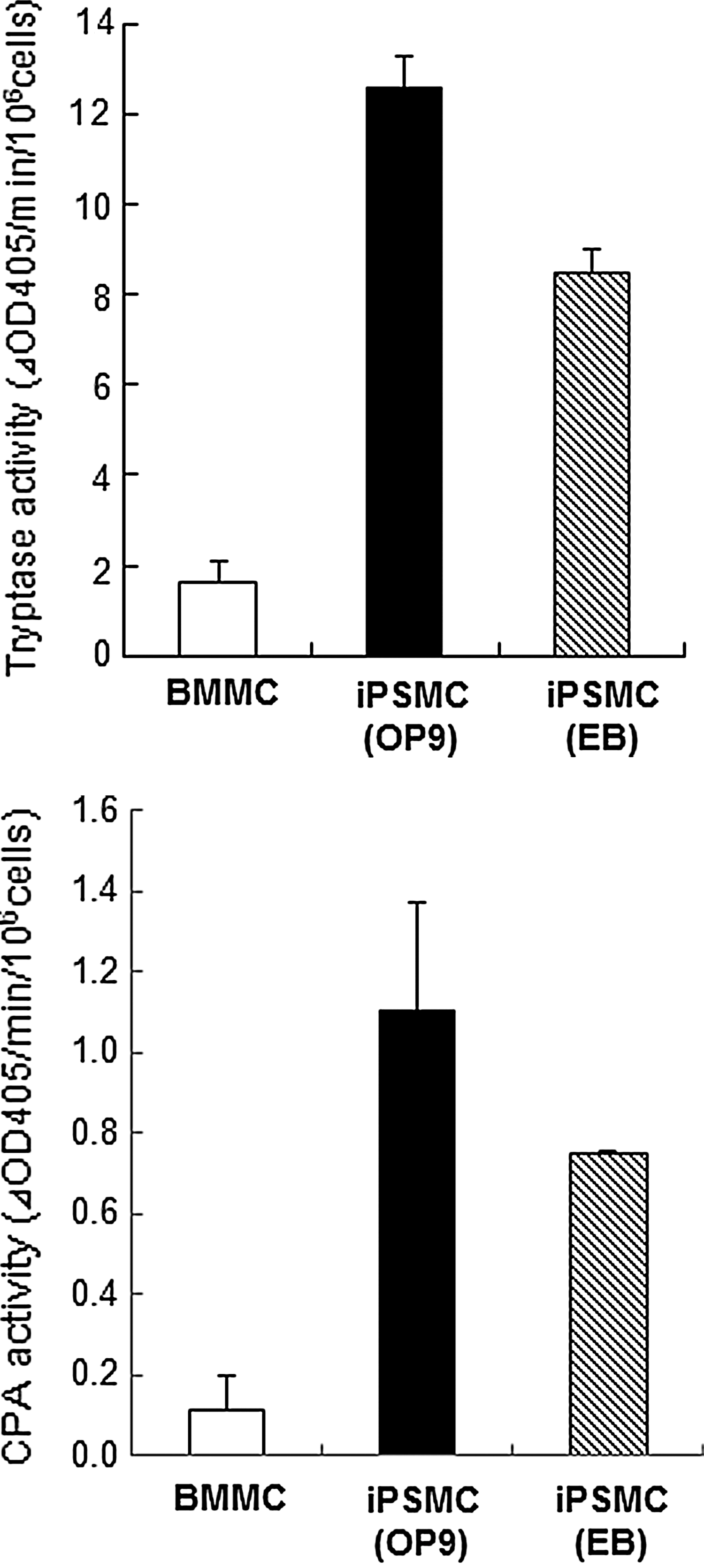
Tryptase and carboxypeptidase A (CPA) activities in iPSMCs. Cell extracts prepared from BMMCs and iPSMCs were assayed for tryptase and CPA activities as described in the Materials and Methods section. All data represent the means±S.D. (n=4).
Histidine decarboxylase (HDC) is a critical enzyme that is involved in the synthesis of endogenous histamine in mammals [25–26], and is considered to be one of the indices of mast cell maturation [26]. Therefore, quantitative RT-PCR analysis was performed to compare the expression of HDC mRNA levels in iPSMCs and BMMCs (Supplementary Fig. S3). The expression level of HDC mRNA was elevated in the iPSMCs that were differentiated by either method as compared with that in BMMCs.
Previously, Takano et al. demonstrated that CD81, a member of the tetraspanin superfamily, is one of the strikingly upregulated genes in BMMCs cocultured with Swiss 3T3 fibroblasts [8]. CD81 is also considered to be a marker of CTMCs. FACS analysis showed that expression of CD81 was elevated in the iPSMCs differentiated by either method as compared with that in BMMCs (Supplementary Fig. S4). In particular, our results revealed that the iPSMCs that were differentiated by coculture with OP9 cells were almost all CD81 positive and showed a homogeneous population.
Degranulation of iPSMCs
We compared FcɛRI-mediated degranulation of BMMCs and iPSMCs by measuring the extracellular activity of β-hexosaminidase, a marker enzyme for histamine-containing granules. As shown in Fig. 4A, the iPSMCs that were differentiated by coculture with OP9 cells displayed significantly less release of β-hexosaminidase than the BMMCs in response to IgE-mediated antigen stimulation. Similar results were obtained in iPSMCs that were derived from EB formation methods (Supplementary Fig. S5A). On the other hand, when the iPSMCs that were pretreated with anti-DNP monoclonal IgE were incubated with DNP-HSA in the presence of Lyso-PS, which is known to enhance IgE-mediated degranulation in rat peritoneal mast cells [27], the amount of β-hexosaminidase release was increased.
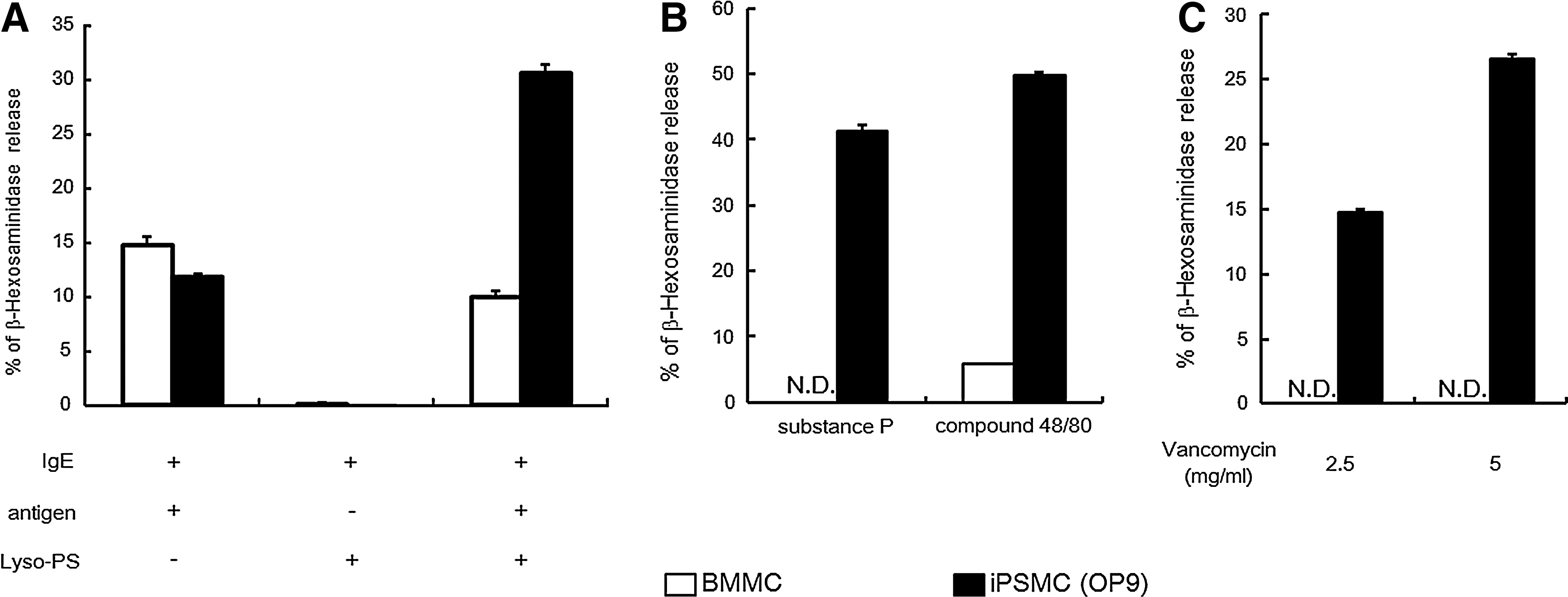
β-hexosaminidase release from iPSMCs after stimulation with IgE/antigen, compound 48/80, substance P, and vancomycin.
Responses to cationic secretagogues such as compound 48/80 and substance P are functional characteristics of CTMCs [4]. We next compared the IgE-independent responses between BMMCs and iPSMCs. Stimulation of iPSMCs with compound 48/80 resulted in marked exocytosis of β-hexosaminidase, whereas no or little release of this enzyme was detected from stimulated BMMCs (Fig. 4B and Supplementary Fig. S5B). Similarly, β-hexosaminidase release from iPSMCs was much more markedly elevated by substance P treatment than β-hexosaminidase release from BMMCs (Fig. 4B and Supplementary Fig. S5B). In addition, stimulation of iPSMCs with vancomycin resulted in marked exocytosis of β-hexosaminidase, whereas no or little release of β-hexosaminidase was detected from vancomycin-stimulated BMMCs (Fig. 4C and Supplementary Fig. S5C). These results indicate that the iPSMCs display a CTMC-like phenotype.
Comparison between iPSMCs differentiated by the OP9 coculture and EB formation protocols
We next compared the degree of differentiation between the iPSMCs that were differentiated by the OP9 coculture method and those differentiated by the EB formation method. The expression level of FcɛRI was significantly lower in the iPSMCs that were differentiated by coculture with OP9 cells as compared with the iPSMCs that were differentiated by EB formation (Fig. 2). However, the number of Safranin O-positive cells was significantly greater in the iPSMCs that were cocultured with OP9 cells than in the iPSMCs that were derived from the EB formation method (Fig. 1B). In addition, the expression levels of HDC mRNA and CD81 protein were significantly higher in the iPSMCs that were cocultured with OP9 cells than in those that were derived from the EB formation method (Supplementary Figs. S3 and S4). These results showed that the iPSMCs that were cocultured with OP9 cells were more mature than the iPSMCs that were derived from the EB formation method. The iPSMCs that were derived from EB formation were more mature than BMMCs (Figs. 1 –3). During the differentiation step, the iPSMCs that were derived from the EB formation method were designated as mast cells I (Fig. 5F). The iPSMCs that were differentiated by coculture with OP9 cells were also designated as mast cells II.
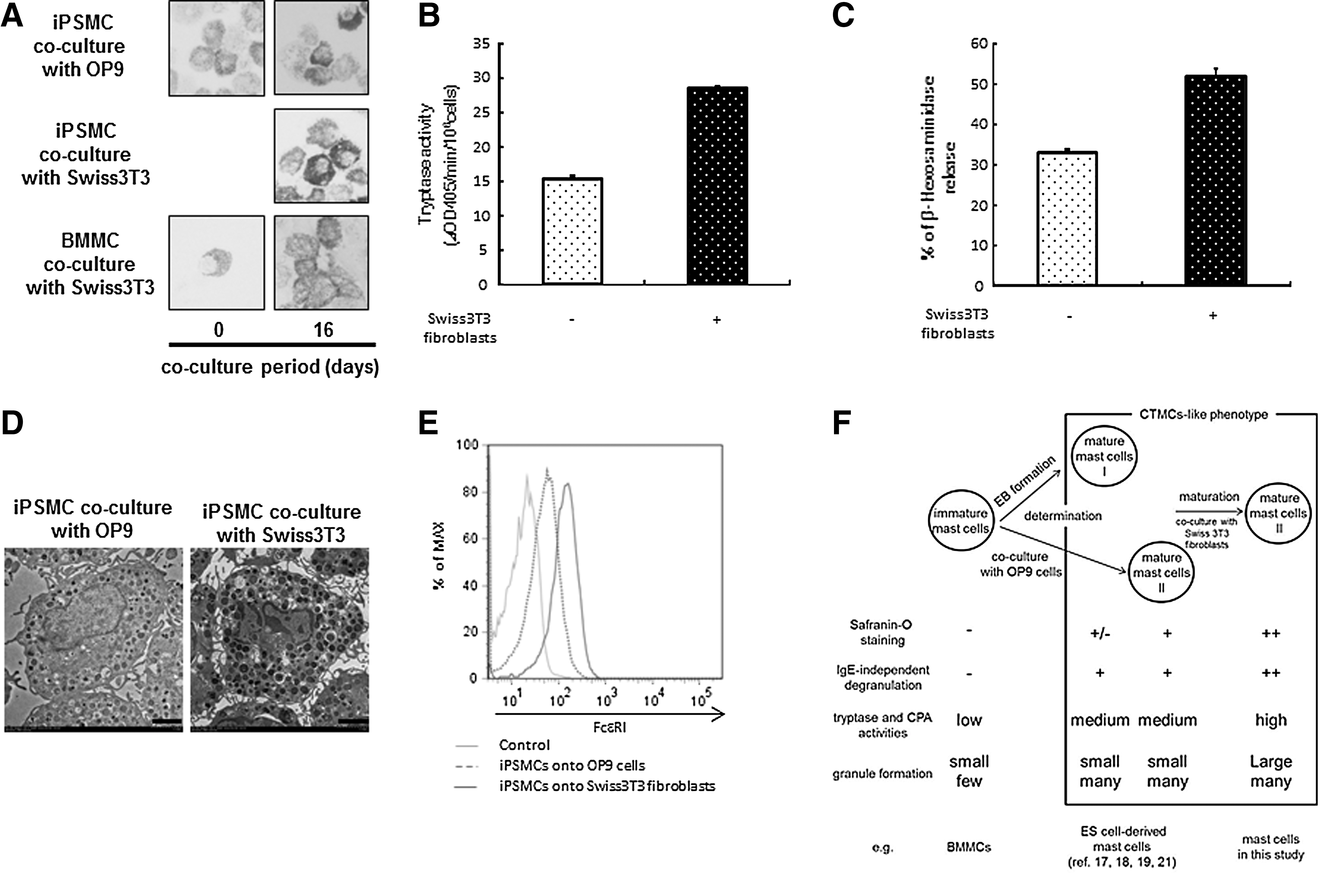
Maturation of iPSMCs cocultured with Swiss 3T3 fibroblasts in the presence of stem cell factor.
Enhancement of maturation in iPSMCs by Swiss 3T3 fibroblasts
A previous study reported that coculture of BMMCs with Swiss 3T3 fibroblasts in the presence of SCF facilitated morphological and functional maturation toward a CTMC-like phenotype [8]. It is possible that Swiss 3T3 fibroblasts promote the maturation of the iPSMCs (mast cells II) that are generated on OP9 cells. Therefore, we expected to generate more mature iPSMCs by coculturing with Swiss 3T3 fibroblasts. We compared the degree of mast cell maturation of the iPSMCs that were cocultured with OP9 cells or Swiss 3T3 fibroblasts. Although the majority of BMMCs were Alcian blue positive and Safranin O negative, the percentage of Safranin O-positive mast cells was gradually increased up to ∼80% on day 16 after coculturing with Swiss 3T3 fibroblasts (Fig. 5A). The staining intensity for Safranin O on the iPSMCs that were cocultured with Swiss 3T3 fibroblasts was stronger than the iPSMCs (mast cells II) (Fig. 5A). Therefore, the iPSMCs that were differentiated by coculture with Swiss 3T3 fibroblasts were found to be more mature than mast cells II and designated as mast cells III (Fig. 5F). We measured the tryptase activity in the iPSMCs (mast cells II) or iPSMCs (mast cells III), and found the elevated tryptase activity in the iPSMCs (mast cells III) relative to the iPSMCs (mast cells II) (Fig. 5B). Similarly, β-hexosaminidase release by compound 48/80 in the iPSMCs (mast cells III) was markedly elevated in comparison with that in the iPSMCs (mast cells II) (Fig. 5C). Electron microscopic analysis of mast cells revealed that the iPSMCs (mast cells III) contained more large granules (Fig. 5D). We performed flow cytometric analysis to examine the surface expression of c-kit and FcɛRI on the iPSMCs (mast cells III). After coculturing, iPSMCs (mast cells III) and BMMCs still expressed similar levels of c-kit (data not shown). Remarkably, the expression level of FcɛRI was elevated in the iPSMCs (mast cells III) (Fig. 5E). These results showed that there were 2 steps in iPSMC maturation process.
Mast cell reconstitution and induction of passive cutaneous anaphylaxis
We assessed whether the iPSMCs (mast cells I) with a C57BL/6 background could exhibit passive cutaneous anaphylaxis 6 weeks after injection of iPSMCs or BMMCs into the conjunctiva of mast-cell-deficient (Kit W-sh/W-sh ) mice [28]. Kit W-sh/W-sh mice reconstituted with BMMCs or iPSMCs exhibited passive cutaneous anaphylaxis reactions in the conjunctivae as measured by extravasation of Evan's Blue dye (Fig. 6). On the other hand, no passive cutaneous anaphylaxis reactions in the conjunctivae were observed for Kit W-sh/W-sh mice that were not reconstituted with mast cells. These results indicate that iPSMCs had the ability to respond to stimulation with IgE/antigen in vivo.
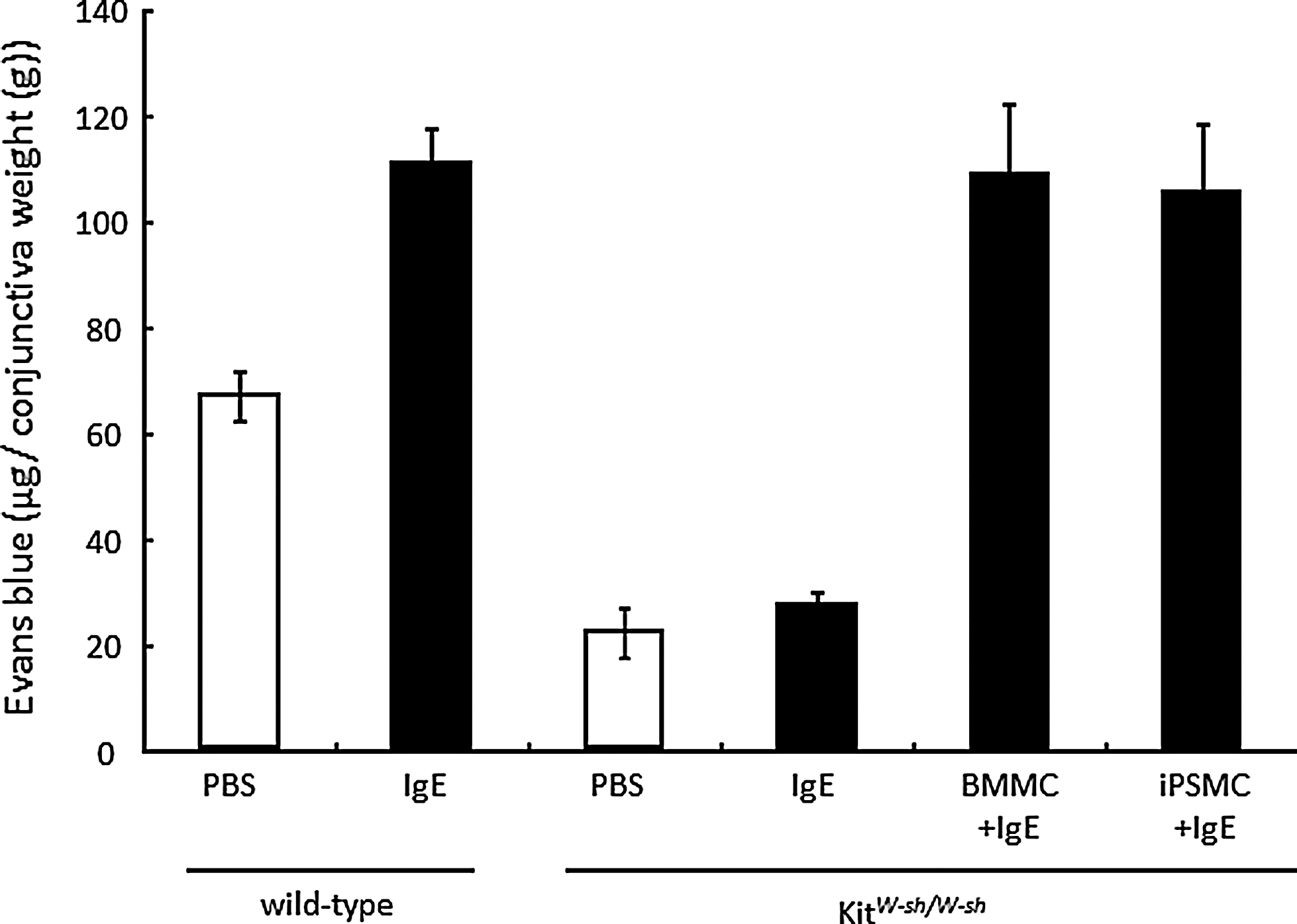
IgE-dependent passive cutaneous anaphylaxis in mast cell-deficient Kit W-sh/W-sh mice reconstituted with iPSMCs. BMMCs or iPSMCs were injected subcutaneously into the conjunctivae of mast cell-deficient Kit W-sh/W-sh mice. After 6 weeks, mice were subcutaneously sensitized with anti-DNP IgE or saline, followed by induction of passive cutaneous anaphylaxis. After 24 h, mice were intravenously injected with DNP-HSA along with 2% Evan's blue dye. Thirty minutes later, conjunctivae were excised, and Evan's blue dye was extracted. Extravasation of Evan's blue dye was quantified as described in the Materials and Methods section. Results are normalized to average conjunctivae weight and are expressed as mean±S.D. (n=3 mice per group).
Discussion
In this study, we developed a protocol consisting of mesoderm induction (stage 1), mast cell specification (stage 2), determination of mast cells (stage 3), and maturation of mast cells (stage 4) for mast cell differentiation from iPS cells (Supplementary Figs. S1 and S6). BMMCs have been used extensively as a mast cell model. We compared the degree of maturation in BMMCs and iPSMCs. Expression level of CD81 was higher in the iPSMCs that were differentiated by both methods than in BMMCs (Supplementary Fig. S4). We also showed that IgE-mediated degranulation of iPSMCs was elevated in comparison with that in BMMCs in the presence of Lyso-PS (Fig. 4A and Supplementary Fig. S5A), demonstrating that iPSMCs were more mature than BMMCs. Therefore, both OP9 cells and EB-derived feeder cells might moderately promote the maturation of mast cells. The other possibility is that the maturation of iPSMCs might be accelerated by SCF. SCF is one of the most important cytokines for mast cell maturation [29]. BMMCs are in general generated without SCF. These are reasons why iPSMCs were more mature than BMMCs with respect to their phenotypes and functions.
Cytokines and feeder cells were required to induce the mast cell development from mouse iPS cells. As previously described, IL-3 is known to play an important role in mast cell specification. In contrast, BMMCs are generated without OP9 cells, suggesting that OP9 cells are not necessary for mast cell specification. OP9 cells might promote the maturation of mast cells, possibly by OP9 cell-derived factors, such as IL-4 [30–31], IL-6 [32 –34], and nerve growth factor [34]. Therefore, cytokines and OP9 cells are all-essential and have distinct roles in the differentiation of mast cells from iPS cells.
We found a difference in FcɛRI expression levels between iPSMCs (mast cell I) and iPSMCs (mast cell II) (Fig. 2). Surface expression level of FcɛRI was lower in the iPSMCs (mast cells II), compared with that in the iPSMCs (mast cells I). More recently, Kovarova et al. reported that expression of FcɛRIα mRNA was lower in human ES cell-derived mast cells that were cocultured with OP9 cells than in human ES cell-derived mast cells that were derived from the EB formation method [21]. These findings were fully consistent with our results. However, our results showed that iPSMCs (mast cells II) were more mature than iPSMCs (mast cells I). These results indicate that the expression levels of FcɛRI are not completely correlated with the degree of mast cell differentiation, although the iPSMCs (mast cells III) showed a high level of FcɛRI expression (Fig. 5E).
In the present study, we demonstrated that, as in the case of BMMCs, Swiss 3T3 fibroblasts could promote the maturation of iPSMCs (Fig. 5). A recent study has reported that cynomolgus monkey ES cells that are cocultured with the murine aorta-gonad-mesonephros-derived stromal cell line AGM-S1 cells are differentiated into CTMCs [20]. These results suggest that feeder cells, including AGM-S1 and Swiss3T3 fibroblasts, would promote mast cell maturation by similar mechanisms. These feeder cells might contribute to the identification of factors that play a role in mast cell maturation.
While iPSMCs (mast cells II) were almost all Safranin O positive, iPSMCs (mast cells I) included both Safranin O-positive and negative populations, suggesting that immature cells were contained in iPSMCs (mast cells I). Expression levels of CD81 protein and HDC mRNA, and protease activities, were slightly elevated in the iPSMCs (mast cells II) as compared with the iPSMCs (mast cells I). Our results suggest that the iPSMCs (mast cells II) were more mature than iPSMCs (mast cell I). The iPSMCs (mast cells III) exhibited more mature phenotypes, such as large granules and high activity of protease. Taken together, the rank order of maturity in mast cells was the following: iPSMCs (mast cells III) > iPSMCs (mast cells II) > iPSMCs (mast cells I). The precise mechanisms of mast cell maturation process remain to be clarified. Classification of mast cells by using cell surface or internal marker can contribute to clarify the maturation mechanism of mast cells. Further studies are needed to find cell surface or internal marker that can clearly distinguish iPSMCs (mast cells I and II) and iPSMCs (mast cells III).
Our data demonstrate that iPSMCs could functionally respond to IgE stimulation in vivo (Fig. 6). There was no significant difference in Evan's blue extravasation in the Kit W-sh/W-sh mice reconstituted with iPSMCs or BMMCs. Previously, Fukuda et al. demonstrated that conjunctiva reconstituted with BMMCs display a CTMC-like phenotype [28]. Therefore, passive cutaneous anaphylaxis reactions were comparable in Kit W-sh/W-sh mice reconstituted with iPSMCs or BMMCs.
Galli's group first reported the identification of mast cell-committed progenitors (MCPs) in adult murine bone marrow [35]. They indicated that MCPs may be directly developed from multipotential progenitors independent of the myeloid pathway. In contrast, Arinobu et al. demonstrated that granulocyte/monocyte progenitors gave rise to MCPs [36]. The models of the developmental process in mast cells differed between these 2 reports. To analyze cells at each differentiation step, our differentiation protocol will be useful for clarifying the developmental process of mast cells.
Because of their pluripotency and self-renewal, ES cells and iPS cells are potential cell sources for regenerative medicine and other clinical applications, such as cell therapies, drug screening, toxicology, and investigation of disease mechanisms. Notably, iPS cell-based screening approaches might support the development of personalized medicine and tailor-made treatment plans. Vancomycin, an antibiotic to which methicillin-resistant Staphylococcus aureus (MRSA) is sensitive, frequently induces allergic reaction [37]. In this study, the stimulation of the iPSMCs with vancomycin resulted in marked exocytosis of β-hexosaminidase, whereas no or little release of this enzyme was detected from BMMCs (Fig. 4C and Supplementary Fig. S5C). Therefore, iPSMCs would be potential cell sources for drug-allergy-screening system.
We developed a 2-step differentiation protocol of mast cells from iPS cells. In the conventional method, CTMC-like mast cells are produced from bone marrow cells after 45 days of culture. In contrast, the iPSMCs generated on OP9 cells in the present study were produced after 28 days of culture. Thus, homogeneous CTMC-like mast cells can be easily generated from iPS cells by the OP9 coculture method. On the other hand, Swiss 3T3 coculture methods have different advantage from OP9 coculture systems. The iPSMCs that were cocultured with Swiss 3T3 fibroblasts were more mature than the iPSMCs that were generated on OP9 cells. Because each of these methods has its advantages, the protocol should be chosen in accordance with the intended use.
We successfully developed a 2-step differentiation protocol for generating more mature mast cells from mouse iPS cells. The iPSMCs generated in this study exhibit many characteristics distinct from BMMCs. The iPSMCs possessed the characteristics of mature mast cells, including the heparin contents and degranulation, in response to cationic secretagogues and vancomycin. The iPSMCs serve as an excellent model for in vitro studies of CTMCs. Our results could facilitate clarification of the mechanisms that control the development of mast cells.
Footnotes
Acknowledgments
We thank Dr. S. Yamanaka for kindly providing the mouse iPS cell lines 38C2. We would like to thank Misae Nishijima and Mary S. Saldon (National Institute of Biomedical Innovation) for technical assistance. We thank Dr. Keizo Nishikawa (National Institute of Biomedical Innovation) for helpful discussion.
Author Disclosure Statement
The authors have no financial conflict of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.