
Abstract
Lectins are carbohydrate-binding proteins, which occur ubiquitously in nature and are abundant in all living organisms from bacteria to mammals. They have several biological functions among which cell adhesion is well known and characterized. Based on the characterization of the glycome of human embryonic stem cells (hESCs), we have investigated the properties of glycan-binding lectins as a novel class of culture support matrices supporting hESC culture. We report that an Erythrina cristagalli lectin (agglutinin) (ECA) matrix supported the undifferentiated growth and significantly increased the plating efficiency of both hESC and human induced pluripotent stem cells when used in conjunction with pinacidil, an antihypertensive drug with ROCK inhibition activity. As a matrix, ECA maintained pluripotency, robust proliferation with a normal karyotype, and the ability to differentiate both in vitro and in vivo. Therefore, our findings indicate that lectins are potential candidates for design of culture and differentiation methods, and that ECA is a potent simple defined matrix for human pluripotent stem cells.
Introduction
Lectins are carbohydrate-binding proteins, which occur ubiquitously in nature and are abundant in all living organisms from bacteria to mammals. They have several biological functions among which cell adhesion is well known and characterized [20].
In some cases, they have been reported to enhance also cell growth and proliferation. For example, it was reported that the β-galactoside-binding lectin jacalin induces proliferation of T lymphocytes [21], and other β-galactoside-binding lectins from peanut and mushroom stimulate vascular cell proliferation [22]. The reported beneficial effects on cell proliferation have been specific for both the cell type and the lectin, suggesting that specific interaction between lectins and cell surface glycans are essential for the growth supporting effect.
We have previously characterized the expression profile of the N-glycans present on the surface of undifferentiated human embryonic stem cells (hESCs) [23]. In the current study, we have used this background information to test several lectins, which bind to the epitopes commonly expressed in hPSC in supporting the growth of undifferentiated hPSC.
Best results were obtained with the lectin (agglutinin) from Erythrina cristagalli, ECA (also called ESL), which binds to N-acetyl lactosamine (type 2 chain) glycoconjugates, a common structure in N-glycans of undifferentiated stem cells [23].
Materials and Methods
Cell culture and maintenance
Three hESC lines (FES 29, FES 30, and H9) [24] and 2 human induced pluripotent stem cell (hiPSC) line (FiPS 5-7 [25] and HEL11.4) were included and cultured on Matrigel as previously described [15,24]. HEL11.4 was generated from adult fibroblasts (men, 84 years old) using retrovirus-induced overexpression of 4 genes: Oct-4, Sox2, Klf4, and c-Myc. Cells were infected with equal parts of the hES medium and virus-containing supernatant twice at 24-h intervals. Cells were harvested and reseeded on mitotically inactivated treated mouse embryonic fibroblast (mEF) layer 3 days after infection. Twenty-four days post-transduction, ES-like colonies were picked, expanded, and characterized.
Cells were passaged by using 0.1 mg/mL collagenase IV (Invitrogen) for 5 min at +37°C and harvested onto ECA (Sigma-Aldrich) and Matrigel (Becton Dickinson) plates and cultured either in StemPro® or in mEF-conditioned-medium (CM) 9KnockOut™-DMEM supplemented with 20% KnockOut™ serum replacement, 2 mM Glutamax, 0.1 mM β-mercaptoethanol, and 0.1 mM nonessential amino acids, all from Invitrogen and supplemented with 8 ng/mL recombinant human basic fibroblast growth factor (bFGF; Invitrogen]. Pinacidil (Sigma-Aldrich; 100 μM) was added to the culture medium during passaging. In all experiments, Matrigel™ (BD Biosciences) was used as a control matrix. The Matrigel plates were prepared as recommended by the manufacturer.
Coating of plates with ECA
ECA lectin (Sigma-Aldrich) solution [1 mg/mL in phosphate-buffered saline (PBS)] was let to passively adsorb onto surface of the cell culture plates (5 μg/cm2) (Nunc, Corning Life Sciences) o/n at +4°C, followed by washing twice with PBS. The coated plates were stored at +4°C and used within 4 weeks.
Flow cytometry analysis of surface antigens
Single-cell suspensions were generated by incubation with TrypLE (Gibco) for 5 min at +37°C. Cells were stained with specific cell surface antibodies (SSEA-1, SSEA-3, Tra 1-60, H type 1, and CXCR4) and fluorescein-conjugated secondary antibody before analysis by flow cytometry (FACS Calibur; BD Biosciences). Antibodies are listed in Table 1.
Immunohistochemistry of the cells on ECA
Cells were fixed with 4% paraformaldehyde and permeabilized with 0.1% Triton X-100 (Sigma-Aldrich) if needed. The antibodies used are listed in Table 1. The cells were probed with secondary antibodies for 30 min in the dark at room temperature. Cells were mounted using a Vectashield mounting medium with 4,6-diamidino-2-phenylindole (Vector Laboratories).
Validation of ECA binding specificity
Cells were passaged and plated as described earlier. The 2 compounds expected to act as specific binding inhibitors for ECA were lactose monohydrate (Sigma-Aldrich; 100 mM) and lacto-N-neotetraose (Kyowa Hakko Kogyo; 100 mM). Saccharose (Sigma-Aldrich; 100 mM) was used as a control. The inhibitors were added to the culture medium at the time of passaging, and the attached cells were counted after 20 h.
In vitro differentiation
Hepatic differentiation was done on FES 29, H9, and HEL11.4 cells, which had been cultured either on ECA or Matrigel for at least 10 passages. The differentiation protocol is described in Table 2.
DE, definitive endoderm; Act A, activin A; NaB, natrium butyrate; KO-DMEM, KnockOut™-DMEM; KO-SR, KnockOut™ serum replacement; NEAA, nonessential amino acids; DMSO, dimethyl sulfoxide; FBS, fetal bovine serum; HGF, hepatocyte growth factor; Onc M, oncostatin M.
Teratoma formation
Cells were harvested with collagenase IV from ECA and Matrigel plates, and ca. 100,000 cells from each matrix were injected into nude mouse testis. After 7–8 weeks, tumors were dissected, fixed with 4% paraformaldehyde, and hematoxylin and eosin-stained sections histologically examined. The animal experiments were approved by the experimental animal welfare committee of the District Government of Southern Finland.
RNA isolation and quantitative polymerase chain reaction
Total RNA was isolated using NucleoSpin® RNA II (Macheray-Nagel GmbH & Co. KG) according to the manufacturer's instruction. Complementary DNA was synthesized from 50 μg of total RNA using iScript™ cDNA Synthesis Kit (Biohit) according to manufacturer's instruction.
Real-time SYBR Green quantitative polymerase chain reaction (qPCR) analyses were performed with Corbett Rotor-Gene 6000 (Corbett Life Science) using the following conditions: 95°C 7 min and 40 cycles of 95°C, 20 s; 56°C, 20 s; 72°C, 20 s. The data were analyzed according to the comparative Ct method (Applied Biosystems; User Bulletin No. 2). Cyclophilin gene expression was an internal reference for normalization. All samples and controls were analyzed in duplicates. Primers used for qPCR are shown in Table 3.
PCR arrays
FES 29 cells were cultured for 9 passages on ECA or Matrigel in CM. Total RNA was isolated from 3 separate plates using RNeasy Mini kit (Qiagen), and complementary DNA was synthesized from 1 μg of total RNA using RT2 First-Strand Kit and RT2 qPCR Master Mixes (SABiosciences) according to manufacturer's instruction. The RT2 qPCR primer assays (SABiosciences) were used to study the gene expression profile of genes related to the identification, growth, and differentiation of stem cells (array PAHS-081).
Karyotype analysis
Karyotype was detected by G-banding technique in the cytogenetics laboratory of the Yhtyneet Medix Laboratories, Inc. Twenty metaphases were examined from each sample.
Clonogenicity assay
Cells were dissociated with TrypLE for 5 min and passed through an 80-μm cell strainer (Becton Dickinson). Dissociated single cells from either ECA or Matrigel were seeded onto both ECA and Matrigel (35 cells/cm2) and cultured in an mEF-CM supplemented with 8 ng/mL bFGF. Pinacidil (100 μM) was used during passaging. To evaluate clonogenic capacity, cells were alkaline phosphatase stained, and colony numbers were counted 10 days after plating.
Cell viability analysis
Cells were plated and cultured on ECA and Matrigel 6 days. Cell viability was analyzed in the beginning, on day 3 and on day 6 using Trypan Blue staining of dissociated cells. The results represent 8 separate experiments, each performed in duplicate. Cell viability was tested also on plate without dissociation using the Live/Dead Viability/Cytotoxity Kit (Invitrogen) according to manufacturer's instructions.
Determination of cell growth rate
FES 29 and HEL11.4 cells were passaged by collagenase IV to small clumps from ECA and Matrigel and plated on 12-well plates, ∼6,000 cells/well on both matrices. Cells were counted at 2 time points, day 3 and 6.
Live cell imaging was used as an alternative method. For this purpose, FES 29 and HEL11.4 cells were dissociated by collagenase IV to small aggregates of 10–20 cells from ECA and Matrigel and plated on 12-well plates, ∼1,000 cells/well on both matrices. Cells were let to adhere in the cell culture incubator for 24 h, and the plates were then transferred into Cell IQ culture platform (CM-Technologies). All wells were imaged every second hour for 5 days. The images were analyzed using a Cell IQ Analyzer.
Statistics
Statistical significance between 2 groups was tested using Students t test. One-way analysis of variance (ANOVA) was used for comparison of multiple groups, followed by Tukey's test.
Results
Testing of lectins
Based on our previous analysis of the N-glycans expressed on the surface of hESCs [23], we selected specific lectins as candidates for the cell culture matrix. ECA (binding specificity in type 2 N-acetyl-lactosamine structures), Maackia amurensis agglutinin (MAA, specific for α-2,3-linked sialic acid), Wisteria floribunda agglutinin (binding preferentially to N-acetylgalactosamine in α- or β-linkage), and Phytolacca americana agglutinin (with N-acetylglucosamine specificity, binding also to polylactosamine structures) were tested for their ability to act as a growth supporting matrix for ESC lines FES 29 and FES 30 in mEF-CM. In cell culture conditions, stem cells attached onto ECA and MAA, but continuous growth, was acquired only on an ECA matrix (Supplementary Fig. S1; Supplementary Data are available online at
Basic characteristics of hPSCs cultured on ECA
Long-term culturing on ECA-coated plates was evaluated with hPSC lines (FES 29 and HEL11.4), and the results were compared to the same cell lines cultured on Matrigel. For most of the experiments, the cells were cultured in the mEF-CM and treated with pinacidil during passaging. Without pinacidil the cells did not attach as effectively, and they also partly changed morphology forming a lot of feeder-like cells. In long-term cultures, the analysis of these cell lines by immunocytochemical stainings (Oct4, Nanog, Sox2, and E cadherin) and flow cytometry (Tra 1-60, SSEA-3, H-type1, and SSEA-1) demonstrated a profile characteristic for undifferentiated hESCs (Fig. 1A, B). The expression levels of major pluripotency-associated genes remained essentially similar throughout 20 passages on both matrices. Minor upregulation of primitive streak/early differentiation markers Brachyuru and Goosecoid occurred at later passages of FES 29 cells on ECA (Fig. 1C). With the iPSC line HEL11.4, the pluripotency genes tended to remain higher and the differentiation genes lower on ECA throughout the culture period (Fig. 1D). In general, FES 29 showed a constant gene expression pattern independent of the matrix as tested by the PAHS-081 qPCR array, including 84 genes controlling growth and differentiation of stem cells (Fig. 1E). The ECA-cultured cells also retained their full in vivo differentiation capacity as indicated by highly complex teratoma containing all 3 germ layer derivatives (Fig. 1F). Both cell lines were karyotypically normal after 18 passages on ECA (not shown).

We next tested the ability of ECA to support the growth of undifferentiated hPSC in a defined cell culture medium StemPro™. Cells were first adapted to StemPro for 1 passage using 1:1 mix of StemPro and CM media, and then only StemPro was used. The results indicated that also defined media supported self-renewal, and cells maintained stem cell markers and normal karyotype detected after 9 passages on ECA in StemPro (data not shown).
Clonogenicity and cell growth
The ability of the ECA matrix to support clonogenicity of hPSC cells was studied by plating dispersed cells first adapted to ECA or Matrigel for at least 2 passages on either of the 2 matrices at the density of 35 cells/cm2. In the presence of pinacidil, the colony-forming efficacy was clearly highest (10.3%,) when ECA-adapted cells were plated on ECA, as compared with all other conditions where the efficacy was ∼6% (P<0.05, one-way ANOVA, Tukey's post hoc test) (Fig. 2A). Pinacidil was found to be essential for the development and survival of the single-cell-derived clones in these experiments.
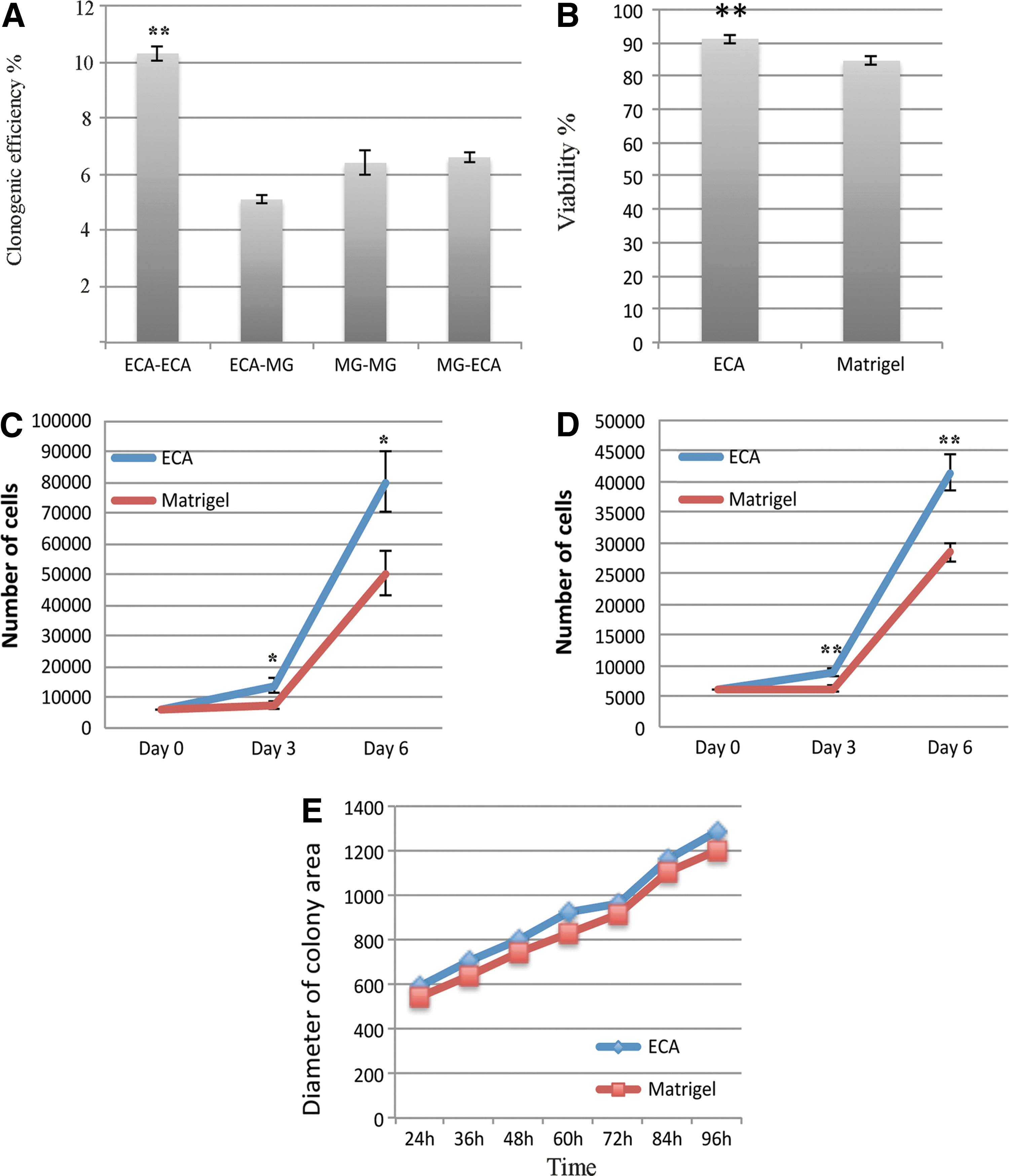
Long-term cell imaging was used to study colony area and cell growth. Colonies were imaged every second hour during 4 days after plating to record colony areas and the number of cells in the colonies. In accordance with the clonogenicity assay, the initial number of colonies was higher on ECA than on Matrigel. An explanation to this was provided by cell viability analysis, which showed higher viability for cells grown on ECA than on Matrigel (90.1% vs. 82.9%, P<0.01, Fig. 2B). The cells were counted 3 and 6 days after plating. The number of cells was significantly higher on ECA than on Matrigel at both time points (Fig. 2C, D). No difference in speed of cell division was detected, and the size of the colonies growing on ECA and Matrigel was similar (Fig. 2E). These results show that culture on ECA generates more cells based on better attachment and survival after dissociation and plating.
Validation of binding specificity
To assess the specificity of the cell–lectin interaction in supporting stem cell attachment to the growth surface, we performed inhibition experiments with specific disaccharide inhibitors and control disaccharides. Lactose (composed of galactose β1,4-linked to glucose) inhibited cell attachment effectively at 100 mM concentration, while the same concentration of saccharose (fructose α1,1-linked to glucose) had no inhibitory activity. Further, lacto-N-neotetraose oligosaccharide, which contains the β1,4-linked galactose epitope, was as effective as lactose (P<0.001) (Fig. 3). The inhibition experiments thus demonstrated that initial cell attachment to the ECA matrix was dependent on specific interaction of the surface-bound lectin with stem cell glycan ligands. The experiments were performed either in the presence (Fig. 3A) of absence (Fig. 3B) of pinacidil. Even if the effect of the inhibitors was similar in both conditions, the total number of attached cells was 4-fold higher with pinacidil.
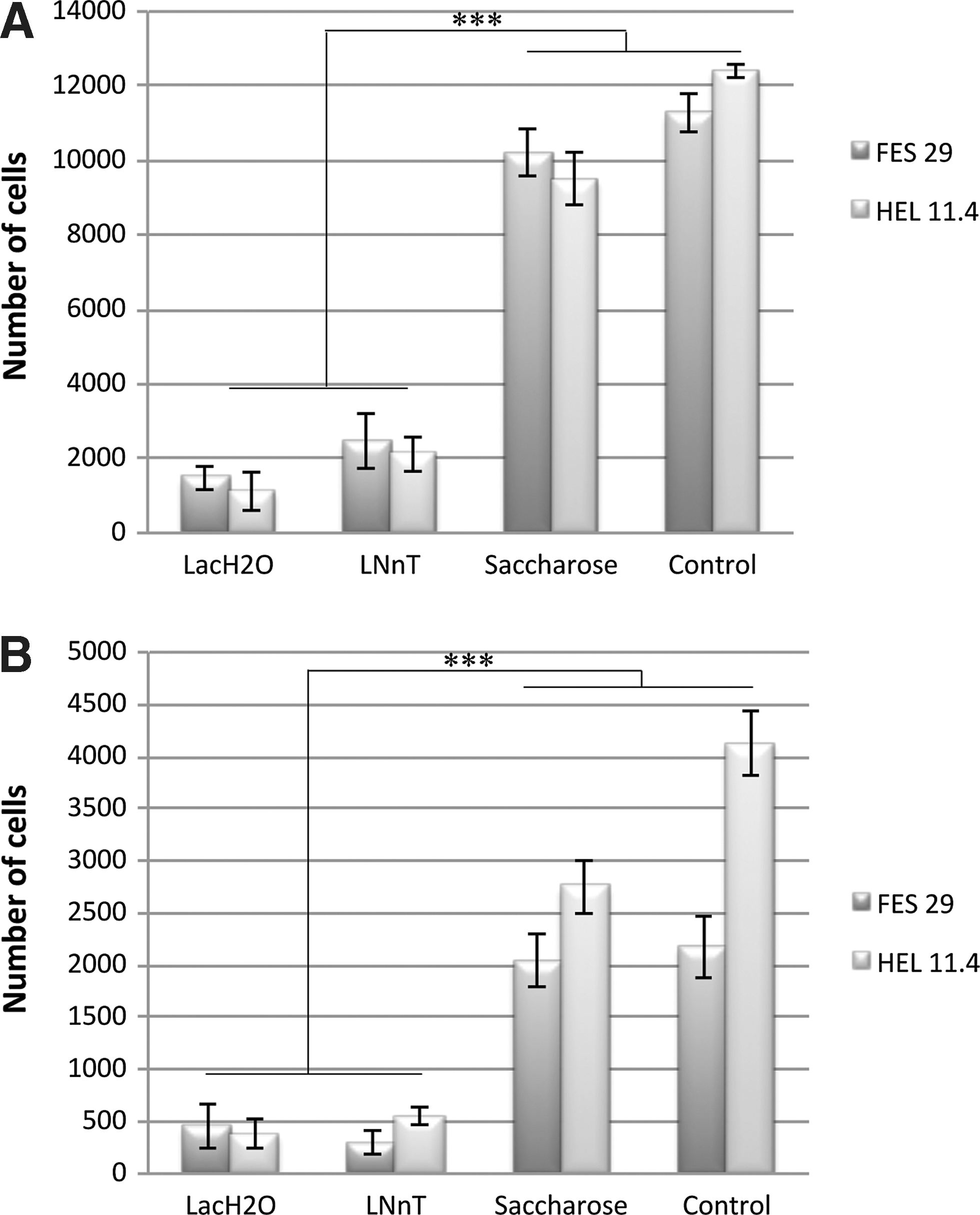
Validation of the binding specificity of hESC and hiPSC to ECA using specific competitive inhibitors either in the presence
Hepatocyte differentiation
The cells were differentiated into hepatocyte-like cells (HLCs) on ECA side by side with Matrigel® as a control. For hepatocyte differentiation, we used a 3-step protocol, modified from the one established by Hay et al. [26]. During the course of differentiation, the expression of the pluripotency marker gene OCT4 was efficiently downregulated while the endoderm marker FOXA2 and anterior definitive endoderm (DE) marker Cerberus (CER1) were strongly upregulated (Fig. 3A). The cells on both coatings changed into morphologically typical DE cells and stained positive for FOXA2 (Fig. 3B). DE induction yielded on average 68%±5% cells positive for CXCR4 on ECA, while using Matrigel, the average was 78%±17% (Fig. 3C). However, cells detached easier from ECA than from Matrigel during the DE stage. The DE cells were further differentiated into hepatocyte progenitors with 5 days of dimethyl sulfoxide treatment. The cells formed hepatic endoderm with α-fetoprotein-positive progenitors on both matrices (data not shown). When the cells were matured into HLCs with hepacyte growth factor and Oncostatin M treatment (d10–d20), the more mature hepatocyte marker albumin became strongly expressed as shown by qPCR and immunocytochemistry (Fig. 4D, E). Taken together, hPSCs were successfully differentiated into HLCs on ECA-matrix, and no significant difference was detected when compared to cells differentiating on Matrigel.
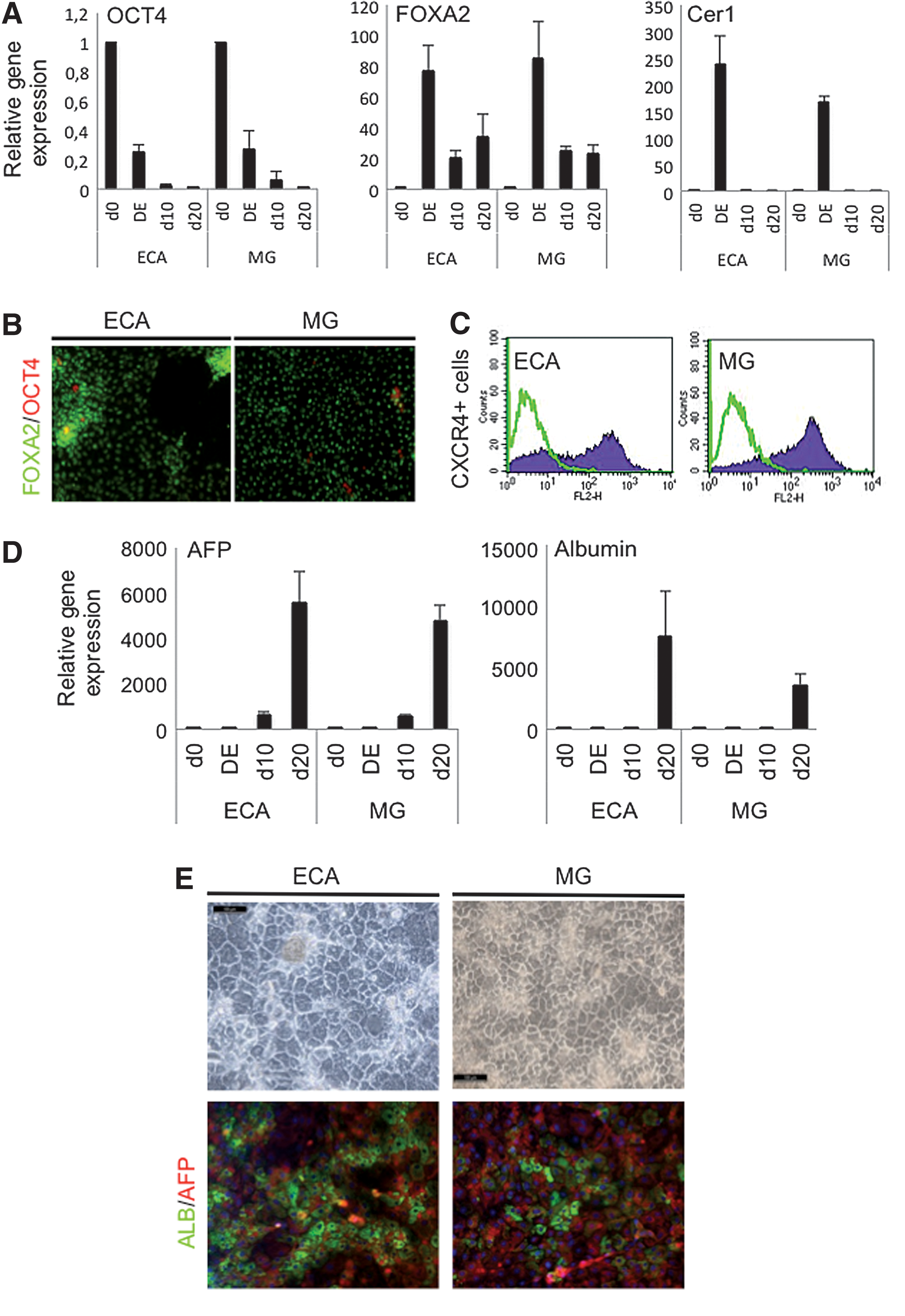
Hepatic differentiation of HEL11.4 cells cultured on either ECA or Matrigel (MG).
Discussion
In this study, we have demonstrated the ability of ECA–lectin to support hESC and hiPSC cell self-renewal in mEF-CM and in commercial defined media. We also show that when passaged with pinacidil, the expansion of the cells is more efficient than on Matrigel without any impairment of quality. As ECA is a small-sized single protein that can be easily produced recombinantly [27], and therefore it is a suitable matrix for a good manufacturing practice use.
It has been reported in many studies that culture adaptation is an evident phenomenon in hPSC cultures. Adapted stem cells proliferate better and quickly become a dominant population in the culture [28 –30]. Extreme adaptation may lead to impaired differentiation capacity of the cells and enrichment of chromosomal abnormalities [31]. The cells cultured on ECA maintained a normal karyotype and pluripotent differentiation capacity. An important feature of the current ECA-based culture method is use of Pinacidil (or ROCK inhibitor) at the time of passaging. Pinacidil is a small molecule that has been widely used for years as a vasodilating drug. It mimics efficiently the effect of ROCK inhibitors in supporting the attachment and viability of dispersed stem cells [32 –34]. Even after, more than 15 passages on ECA, the benefit from use of Pinacidil remained unchanged. This suggests that our culture method does not select for an adapted subpopulation that over time survives on the single-compound matrix. The teratoma formation and hepatocyte differentiation experiments suggest that long-term culture on ECA does not alter the differentiation capacity of the cells. However, further studies of various differentiation pathways are needed to confirm this.
Lectins, the carbohydrate-binding proteins, are identified in a great number of animal cells. The carbohydrate structures to which ECA binds, terminal N-acetyl lactosamine (Galβ1-4GlcNAc) and 2′-fucosyl-N-acetyl lactosamine (Fucα1-2Galβ1-4GlcNAc) are frequently found in cell glycoproteins and glycolipids of mammalian origin, also in undifferentiated hESC and their early differentiation derivatives [23,35,36]. Also galectins, the mammalian counterparts of ECA have been implicated in many biological phenomena, including cell–cell and cell–ECM adhesion [37].
Human galectin-1 has binding specificity related to ECA and in preliminary experiments, we were able to support both hESC attachment and undifferentiated proliferation by using recombinant galectin-1 (unpublished results). In general, the adhesion activity of lectins correlate with the presence of its receptors on the cell surface [38]. However, the affinity between a lectin and its receptor may vary a great deal due to small changes in the carbohydrate structure of the receptor and it is difficult to identify the key interactions mediating a cell's binding to ECA.
Various specific glycan structures are known to be expressed in stem cells in developmentally regulated fashion [23,39]. However, little is known about their potential biological functions, but a plausible mechanism is interaction with cellular lectins. Adhesion molecules, including fibronectin and CD44, contain functionally important lectin domains mediating interaction with the ECM. The use of lectins as cell culture matrixes is not self-evident. For example, lectins binding to mammalian cell surface can be mitogenic [21,22], and the β-galactoside-binding lectin ricin is highly toxic. This was also demonstrated by our initial functional experiments; only few of the tested lectins that bound to hESCs supported their initial attachment, and only ECA was able to support continuous cell culture. The capacity of ECA to act as a support for undifferentiated growth is likely to depend on high-affinity cell-type specific adhesion, which is not associated with any toxic or differentiation-inducing effects. There is also evidence for the activation of specific growth-stimulating signaling pathways after binding of mouse ESCs to a β-galactoside-binding lectin, galectin-1 [40]. However, we did not observe direct mitogenic effects for ECA.
The primary role of a cell culture matrix is to stimulate the cell-dependent attachment response. In addition, the matrix provides signals for growth and differentiation of the cells. However, these functions may be less important in a complete complex culture medium. Adhesive epitopes, which often are glycans, and their density on the substrate as well as combination with other ligands influence the cell adhesion. Previous reports have indicated that at least laminins 511 and 111 as well as vitronectin and fibronectin are effective for the attachment and growth of undifferentiated hPSC [11,14,15,41]. However, these large and complex proteins are difficult and expensive to produce. Using currently commercially available preparations, the cost of plate coating is approximately similar with ECA and Matrigel (1:20). When comparing costs and benefits of ECA and Matrigel, the cell expansion is clearly more effective on ECA, saving both money and time. In our hands, 10,000 hESCs plated on ECA produce over 2 million cells during 8–10 days while on Matrigel this takes 12–14 days. For many purposes, clonal expansion is essential, and this is significantly better on ECA than on Matrigel.
Overall, our results suggest that lectins may serve as a natural platform for culture and differentiation of stem cells. Lectins may also provide a simple and cost-effective platform for studies of additional signals in self-renewal and differentiation. Based on the efficient expansion of karyotypically and functionally intact cells, recombinant ECA is a useful GMP-applicable alternative as a defined coating substrate for hPSC culture.
Footnotes
Acknowledgments
The authors are grateful to Eila Korhonen, Heli Mononen, Jaan Palgi, and Jarkko Ustinov for help with cell culture and analysis. This study was supported by the Finnish Funding Agency for Technology and Innovation (TEKES), the ESTOOLS project of the EU 6th FP, and the LIV-ES project of the 7th FP. Further support was received from the Academy of Finland and the Sigrid Jusélius Foundation.
Author Disclosure Statement
T. S., J.N., and J.S. are shareholders of Glykos Finland Ltd.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.