
Abstract
Adult stem cells are critical for the healing process in regenerative medicine. However, cigarette smoking inhibits stem cell recruitment to tissues and delays the wound-healing process. This study investigated the effect of nicotine, a major constituent in the cigarette smoke, on the regenerative potentials of human mesenchymal stem cells (MSC) and periodontal ligament-derived stem cells (PDLSC). The cell proliferation of 1.0 μM nicotine-treated MSC and PDLSC was significantly reduced when compared to the untreated control. Moreover, nicotine also retarded the locomotion of these adult stem cells. Furthermore, their osteogenic differentiation capabilities were reduced in the presence of nicotine as evidenced by gene expression (RUNX2, ALPL, BGLAP, COL1A1, and COL1A2), calcium deposition, and alkaline phosphatase activity analyses. In addition, the microRNA (miRNA) profile of nicotine-treated PDLSC was altered; suggesting miRNAs might play an important role in the nicotine effects on stem cells. This study provided the possible mechanistic explanations on stem cell-associated healing delay in cigarette smoking.
Introduction
Adult stem cells can be considered as resident tissue-restricted stem cells, and they participate in tissue maintenance and regeneration [11]. Mesenchymal stem cells (MSC) are well-studied adult stem cells. They are multipotent and can differentiate into multiple mesodermal lineages, such as osteoblasts, chondrocytes, and adipocytes [12]. Osteogenic differentiation of MSC upregulates several marker genes, such as runt-related transcription factor 2 (RUNX2), alkaline phosphatase (ALPL), osteocalcin (BGLAP), and type I collagen (COL1A1 and COL1A2) [13]. MSC were first identified in adult marrow, but can also be found in most postnatal organs, including the periodontal ligament. Periodontal ligament-derived stem cells (PDLSC) contain a heterogeneous population of MSC and neural crest stem cells. PDLSC are capable of differentiating into the neurogenic, cardiomyogenic, chondrogenic, and osteogenic lineages [14]. Both adult stem cells, MSC and PDLSC, can differentiate, participate, and enhance the wound-healing process [15,16]. However, cigarette smoking inhibits recruitment of MSC to tissues [17]. Active components in cigarette smoke, such as nicotine, will influence stem cell recruitment to the injury site and lead to delayed wound-healing phenomena. Recently, we showed that nicotine will affect the biophysical properties of human MSC, rendering the cells less responsive to mechanoinduction and other physical stimuli [18]. The finding provides further evidence that nicotine can alter stem cell properties and affect their response to tissue injury.
Nicotine exerts its cellular functions through the nicotinic acetylcholine receptors. The subunits of the nicotinic acetylcholine receptor are present in human MSC (α3, α5 and α7) [19] and PDL fibroblasts (α7) [20]. Therefore, we hypothesized that the delayed wound healing due to nicotine exposure is the outcome from its effects on these adult stem cells through genetic and microRNA (miRNA) regulation. In this study, we investigated the effect of nicotine exposure on cell proliferation, cell migration, and osteogenic differentiation potential of the human adult stem cells, MSC, and PDLSC. In addition, the miRNA expression profile of the nicotine-treated human PDLSC was also examined by microarray analysis.
Materials and Methods
Cell culture
Human MSC were purchased from ScienceCell Research Laboratories (Cat #7500; ScienceCell Research Laboratories), whereas human PDLSC lines were established previously [14]. These cells were cultured in the Dulbecco's modified Eagle's medium (Gibco BRL) supplemented with 10% heat-inactivated fetal bovine serum (Gibco BRL). The medium was changed daily to maintain a constant level of nicotine. Cells with passage 3–5 were used for the experiments. Each group would have triplicate samples for each experiment.
Cell proliferation analysis
The proliferation of MSC and PDLSC was evaluated by MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] assay (TACS® MTT Cell Proliferation Assay; Trevigen). 1,500 cells per well were seeded on a 96-well plate (Becton Dickinson), and the cells were treated with or without 1.0 μM nicotine (Sigma-Aldrich). The analysis (8 wells per group) was performed everyday over a 5-day period according to the manufacturer's protocol. Absorbance at 600 nm with a reference of 650 nm was measured by a spectrophotometer (Dynex Technologies).
Cell migration analysis
MSC or PDLSC (7,000 cells) were seeded on a 35-mm dish (Sarstedt, Inc.). The cells were synchronized in a serum-deprived medium, and they were treated with or without 1.0 μM nicotine (Sigma-Aldrich) for 7 days. Bovine serum albumin (0.1 mg/mL; Genotech) was used to stimulate cell migration at the day of imaging. MSC were visualized using a time-lapse microscope (Nikon NIS-Elements AR 3.2), and the cells were maintained at 37°C throughout the imaging period. Images were taken every 15 min over a 2-h period. 30 cells from 5 different areas in the images were traced and measured. The cell nucleus served as the reference point between frames. The distance travelled (μm) was measured using the software of the microscope. Speed (nm/s) was calculated by dividing the distance travelled with time.
Osteogenic differentiation analysis
Osteogenic differentiation was induced by addition of 10 μM β-glycerophosphate (Calbiochem), 10 nM dexamethasone (Sigma-Aldrich), 50 ng/mL BMP-2 (PeproTech), and 50 μg/mL ascorbic acid (Sigma-Aldrich) in the culture medium. For the nicotine-treated group, the osteogenic differentiation medium was supplemented with 1.0 μM nicotine (Sigma-Aldrich).
MSC or PDLSC (9,000 cells) were seeded on a 4-well chamber slide (Thermo Scientific). The cells were treated with the osteogenic differentiation medium for 10 days. For calcium deposition, the cells were fixed in 10% formalin and evaluated by Alizarin Red S staining (BDH Chemicals Ltd.). For alkaline phosphatase activity, the cells were fixed in ice cool acetone and evaluated by Burstone's staining protocol for alkaline phosphatase.
miRNA microarray and data analysis
Human PDLSC were treated with or without nicotine (0, 0.5, and 1.0 μM; Sigma-Aldrich) for 3 days. Total RNA, including the miRNA fraction, was collected and extracted in the TRIzol reagent according to the manufacturer's protocol (Invitrogen). The RNA concentration and quality were measured by Nanodrop 2000, whereas the RNA integrity was determined by the Agilent 2100 Bioanalyzer. The SurePrint G3 Human v16 miRNA Array Kit (8x60K, Release 16.0; Agilent) containing probes for 1205 human and 144 human viral miRNA from the Sanger miRBase v16.0 was used. GeneSpring GX 11.5 software (Agilent) was used for value extraction. A 2-tailed Student's t-test was then used for the calculation of the P value for each miRNA probe. Significance was defined by the fold change greater than 2 and the corrected P value less than 0.05. The Benjamini–Hochberg false discovery rate was used for the multiple testing correction. Principal component analysis was performed to provide a visual impression of how various sample groups are related.
Downstream mRNA targets of the miRNAs were predicted by TargetScan (
Validation of microarray results was performed by semiquantitative PCR. Total RNA (20 ng) was reverse transcribed using the TaqMan MicroRNA Reverse Transcriptase kit (Applied Biosystems). The resultant products were quantified using the appropriate TaqMan MicroRNA Assays (Applied Biosystems) on a BioRad MyiQ™ Real-Time PCR Detection System. Results were all normalized to U6 expression. The independent T-test was used for statistical analysis.
Gene expression analysis
MSC and PDLSC were treated with or without 1 μM nicotine (Sigma-Aldrich) for 3 days. Total RNA was collected and extracted in the TRIzol reagent according to the manufacturer's protocol (Invitrogen). The gene expression analysis was performed using the Sybr green PCR master mix (Applied Biosystems) on a real-time PCR machine (Stratagene). Genes for cell migration and osteogenic differentiation (PTK2, RUNX2, ALPL, BGLAP, COL1A1, and COL1A2) were examined with specific primers (Supplementary Table S1; Supplementary Data are available online at
Statistical analysis
All statistical analyses, except microarray analysis, were performed by commercially available software (SPSS, version 16.0; SPSS, Inc.). The independent T-test was used to compare the means between samples. Significance was defined as P<0.05.
Results
Nicotine reduced human adult stem cell proliferation
Stem cell regenerative potential is determined by different factors. One such factor is related to the availability and abundance of stem cells present to exert a regenerative effect. This potential can be determined by the proliferative rate of the cells. The proliferation of human MSC in the control medium was significantly higher (P<0.05) than that in the nicotine-containing medium (Fig. 1A). The proliferation of nicotine-treated MSC reached a plateau at day 4, while control cells kept an increasing trend in cell number throughout the testing period. Similarly, the proliferation of human PDLSC was also significantly higher (P<0.01) in control media than in the nicotine-treated group (Fig. 1B). The proliferation of nicotine-treated PDLSC reached a plateau at day 3. Both the PDLSC- and MSC-treated groups showed a greater than 2-fold decrease in the total number of cells at day 5 compared with the control group, indicating the loss of proliferative potential while under nicotine treatment.
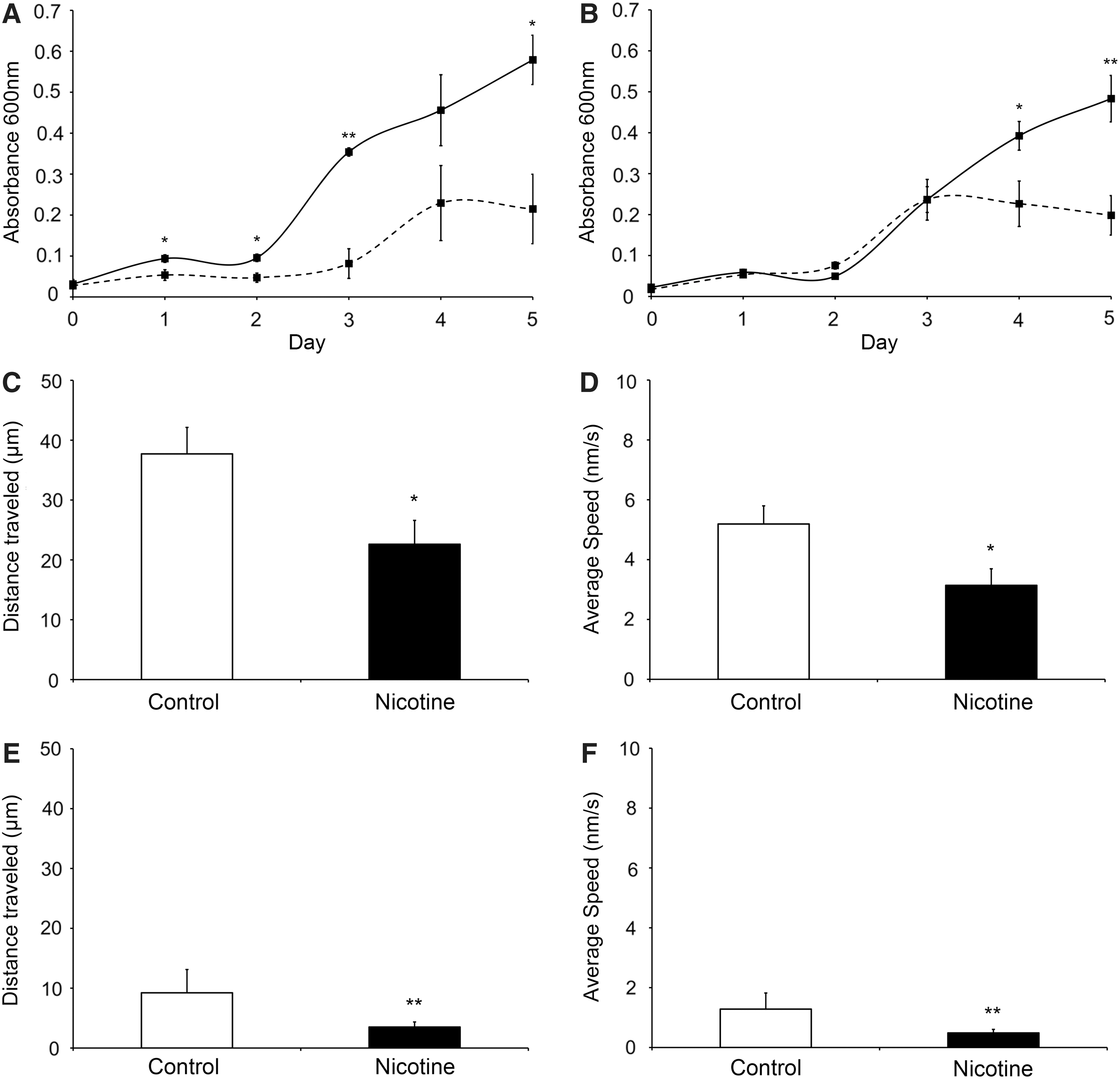
Cell proliferation and migration analysis of human MSC and PDLSC with or without nicotine treatment.
Nicotine inhibited human adult stem cell migration
Another determining factor is related to the movement of stem cells and their capacity to migrate to injury sites. This characteristic can be examined by cell migration analysis. The distance travelled by the nicotine-treated MSC (22.61±3.98 μm) was significantly shorter than that by the control MSC (37.71±4.43 μm, P=0.022; Fig. 1C). The average speed of MSC migration was also slower in the nicotine-treated group (3.14±0.55 nm/s) than in the control group (5.19±0.61 nm/s, P=0.024; Fig. 1D). Analogously, the nicotine-treated PDLSC (3.50±0.86 μm) migrated less than the control PDLSC (9.23±3.89 μm, P=0.008; Fig. 1E). Furthermore, the nicotine-treated PDLSC (0.49±0.12 nm/s) also moved slower than the control PDLSC (1.28±0.54 nm/s, P=0.008; Fig. 1F).
Nicotine reduced human adult stem cell osteogenic differentiation
The ability of stem cells to differentiate into a designated mature cell type is the driving concept behind stem cell regeneration. Under conditioned media stimulation, MSC or PDLSC differentiated into osteoblasts, and their calcium deposition increased (Fig. 2A). However, reduced calcium deposition was observed in the osteogenic differentiation medium supplemented with nicotine. Coherently, blue diazonium salt stain indicating alkaline phosphatase activity was accumulated in the cells under osteogenic differentiation (Fig. 2B). The amount of blue stain was reduced in cells treated with the osteogenic differentiation medium with nicotine. These findings suggest that nicotine can inhibit the differentiation of osteoblasts from MSC or PDLSC.
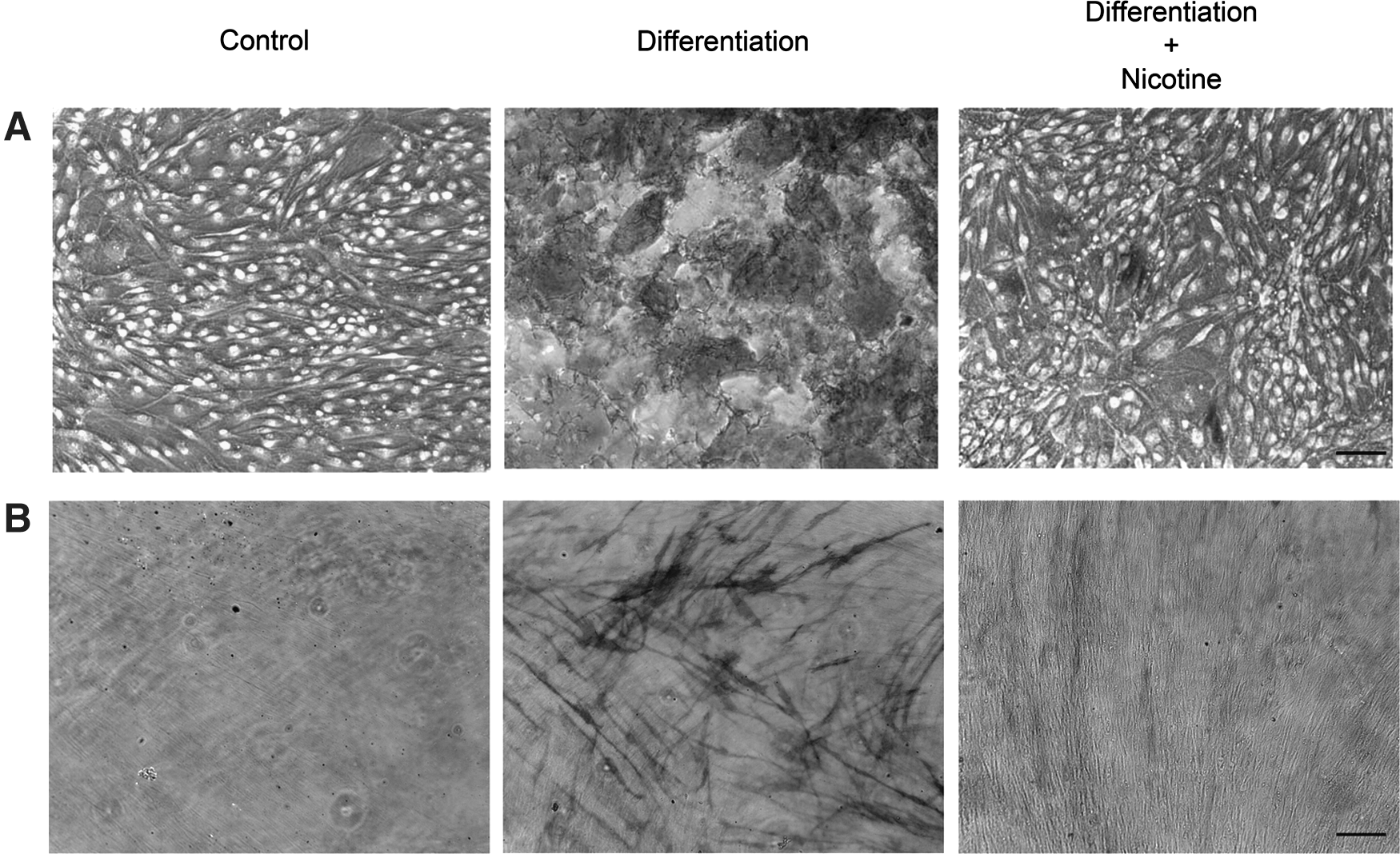
Calcium deposition and alkaline phosphatase analysis of human MSC or PDLSC under osteogenic differentiation with or without nicotine treatment. Osteogenic differentiation was induced by addition of 10 μM β-glycerophosphate, 10 nM dexamethasone, 50 ng/mL BMP-2 and 50 μg/mL ascorbic acid for 10 days. For the nicotine-treated group, the osteogenic differentiation medium was supplemented with 1.0 μM nicotine.
The miRNA signatures of nicotine-treated human PDLSC
To understand the molecular mechanism of the nicotine-induced effect, global miRNA expression patterns (miRNome) of nicotine-treated PDLSC were analyzed by microarray. A total of 225 miRNAs were differentially expressed at a 2-fold difference between nicotine-treated PDLSC when compared with control. The miRNA expression profiles of the control and 1.0 μM nicotine-treated groups were differentially clustered and separated from each other by either principle component analysis (Fig. 3A) or hierarchical clustering (Fig. 3B). The miRNA profile of the 0.5 μM nicotine group lied intermediately between the control and the 1.0 μM nicotine group. Compared to the control group, 16 miRNAs were differentially expressed in the 1.0 μM nicotine group (Pcorr <0.05 and fold change >2; Table 1). 12 of them (hsa-miR-7, hsa-miR-18b, hsa-miR-30d, hsa-miR-137, hsa-miR-374b, hsa-miR-505, hsa-miR-543, hsa-miR-1305, hsa-miR-1914*, hsa-miR-1973, hsa-miR-3198, and hsa-miR-3659) were upregulated, and 4 of them (hsa-miR-210, hsa-miR-762, hsa-miR-1915, and hsa-miR-4281) downregulated. All of these miRNAs showed dose-dependent changes from 0.5 to 1.0 μM nicotine. The top 10 highly expressed miRNAs in Table 1 were selected for validation and showed a similar expression change to the microarray results. Interestingly, 5 miRNAs related to osteogenesis (hsa-miR-29b, hsa-miR-30d, hsa-miR-137, hsa-miR-424, and hsa-miR-1274a) showed Pcorr <0.05 (Table 2).
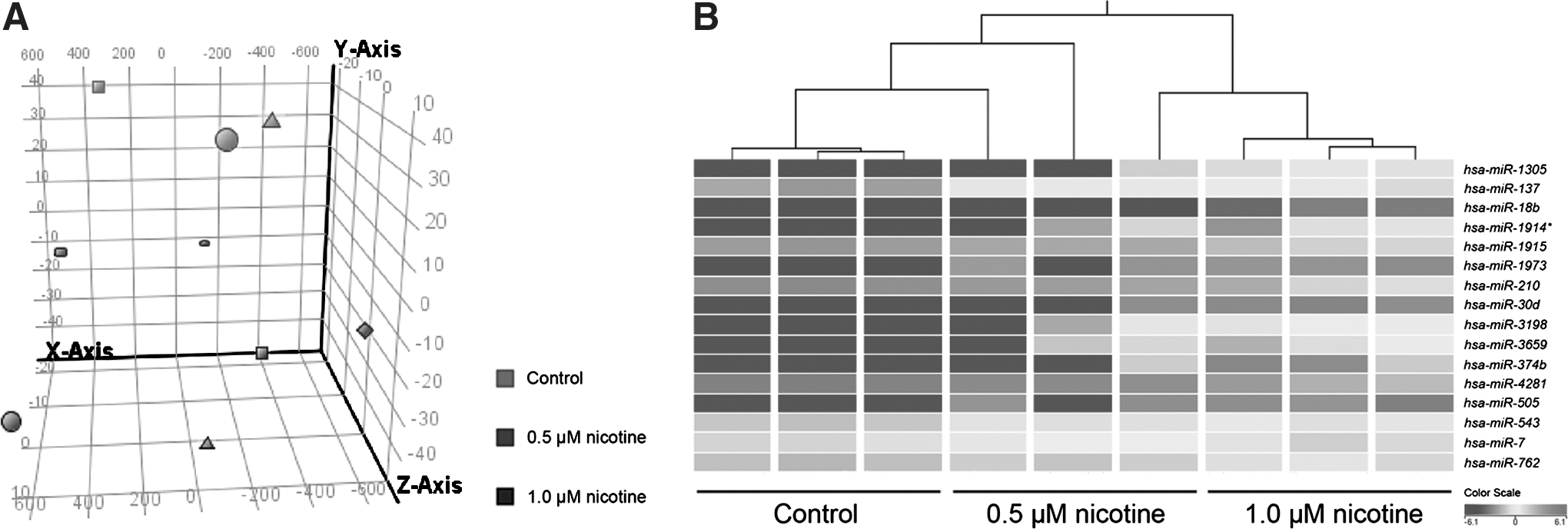
Discrimination of the microRNA microarray profiles in PDLSC with or without nicotine treatment.
Pcorr represents the P value corrected by Benjamini–Hochberg false discovery rate.
denotes the less predominant form.
Bold, indicates the 5 significant microRNAs related to osteogenesis, as described in the Results section.
Subsequently, a global target gene list of the nicotine-associated miRNAs (2,629 genes) was generated by the TargetScan in the GeneSpring (Agilent) platform. Gene ontology of this gene list, analyzed by DAVID (2485 DAVID identities), revealed that nicotine-associated miRNAs might target the genes involved in cell migration, cell division, and bone development (Table 3).
Enrichment score greater than 1.3 was considered as significant.
Gene expression analysis of nicotine-treated human adult stem cells
From the predicted target gene list, RUNX2 associated with bone development (Supplementary Table S2) and PTK2, associated with cell migration (Supplementary Table S3), were predicted as the targets of differentially expressed miRNAs, including hsa-miR-1305, hsa-miR-30d, and hsa-miR-137, which were upregulated in the nicotine-treated group. Subsequent gene expression analysis by semiquantitative PCR revealed that both PTK2 and RUNX2 were significantly downregulated in the nicotine-treated MSC (by 3.0 and 9.1-fold, respectively; P<0.01) and PDLSC (by 2.1 and 2.1-fold, respectively; P<0.01). Nicotine-induced miRNA upregulation might be correlated with the downregulation of PTK2 and RUNX2. In addition, other osteogenic genes (BGLAP, COL1A1, and COL1A2) were also downregulated in the nicotine-treated cells (by 2.0–5.3-folds; P<0.05) compared to the control group, further supporting the inhibitory effect of nicotine on MSC and PDLSC osteogenic differentiation potential (Fig. 4).
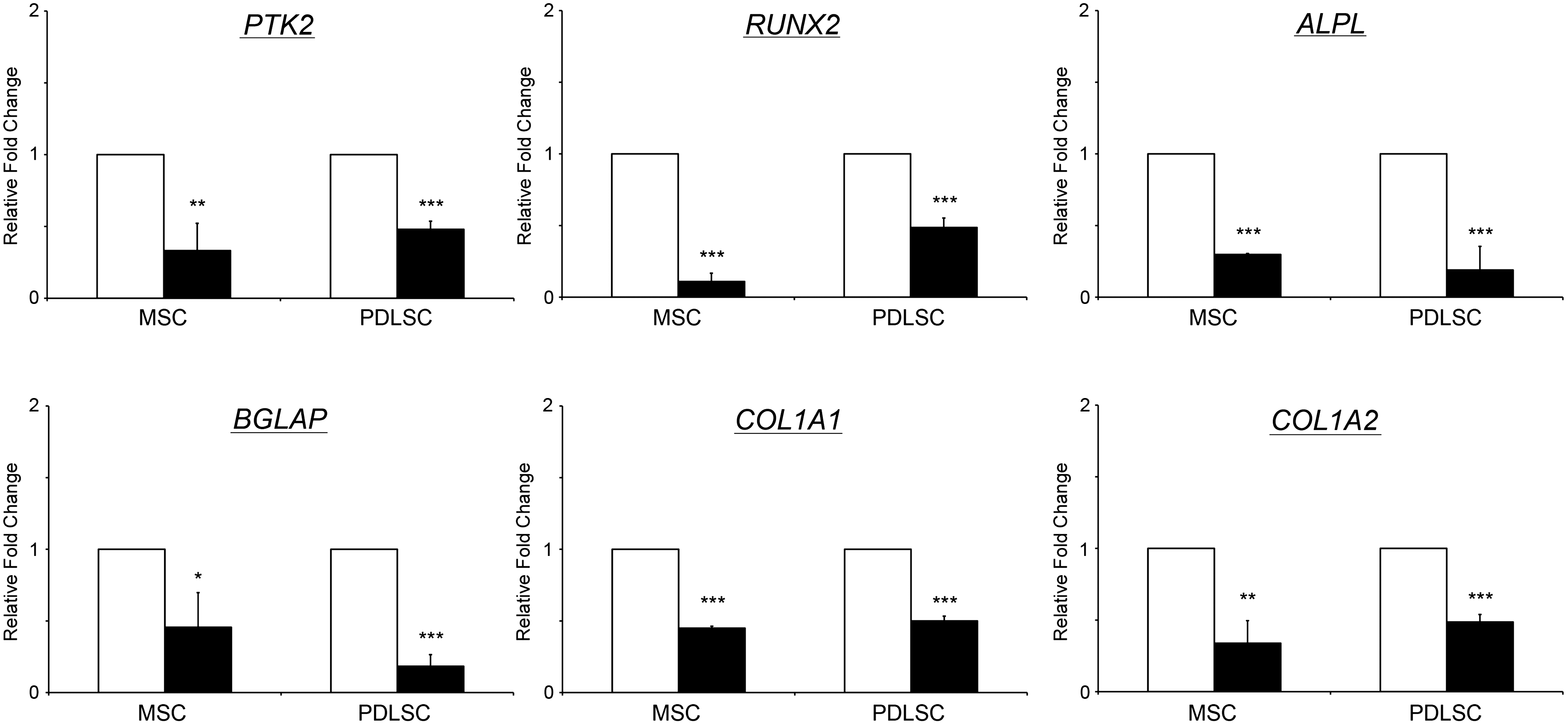
Gene expression analysis of human MSC and PDLSC with or without nicotine treatment. Human MSC (left panel) and PDLSC (right panel) were treated with or without 1 μM nicotine for 3 days. Total RNA was collected and extracted in TRIzol reagent. The gene expression analysis was performed using the Sybr green PCR mater mix on a real-time PCR machine. Genes for cell migration (PTK2) and osteogenic differentiation (RUNX2, ALPL, BGLAP, COL1A1, and COL1A2) were examined. GAPDH was used for normalization. The data represented are the mean±standard deviation. White bars: the control group; black bar: the nicotine treatment group. *P<0.05, **P<0.01, ***P<0.001.
Discussion
Cigarette smoking delays healing process at the injury site [4 –7]. A recent review suggested that nicotine from a cigarette at least, partly, contributes to this delayed healing process [10]. One μM nicotine was applied in this study since its effect on biophysical properties of human MSC was greatest in our previous analysis [18]. Adult stem cells are known to actively participate in this regenerative process [11]. Stem cell regeneration is determined by 3 critical processes: cell proliferation, migration, and differentiation [22]. Cell proliferation determines the amount of stem cells present within the body capable of exerting a regenerative effect. Smoking has been shown to decrease the level of circulating CD34+ progenitor cells in blood [23]. This reduced level could be caused by reduction in cell proliferative rates or induction in apoptosis. Nicotine attenuates cell proliferation of human alveolar bone marrow-derived MSC [24] and PDL fibroblasts [25]. Moreover, nicotine induces apoptosis in PDLSC [26]. Our results further proved that nicotine inhibits cell proliferation in both human MSC and PDLSC (Fig. 1A, B). However, we did not observe a difference in MSC death between the 1 μM nicotine treatment group and the control group in our previous study [18]. Hence, the inhibition of cell proliferation by nicotine might lead to a reduction of available adult stem cells in people smoking cigarettes.
Cell migration allows stem cells to actively move toward the injury sites and contribute to the healing process [27]. Nicotine inhibits the migration of human MSC [28] as well as the PDL fibroblast [29]. This is coherent to the inhibition of human MSC and PDLSC migration by nicotine at 1 μM as evidenced in this study (Fig. 1C–F). Moreover, the downregulation of the focal adhesion kinase (PTK2) in the nicotine-treated human MSC and PDLSC (Fig. 4) further supports the observation of a nicotine-induced reduction of stem cell migration [30]. In addition, this also suggests that reduced human adult stem cell migration by nicotine could be mediated through the downregulation of PTK2 expression.
Stem cell differentiation into tissue-specific cells is one of the mechanisms for stem cell repair at the injury site during the healing process [15,16,31]. In this study, we showed that exposure to nicotine decreased the calcium deposition as well as the alkaline phosphatase activity in human MSC and PDLSC during osteogenic differentiation (Fig. 2). In addition, the expression of the osteogenic genes (RUNX2, ALPL, BGLAP, COL1A1, and COL1A2) was downregulated in the nicotine-treated human MSC and PDLSC (Fig. 4). Nicotine could have affected the osteogenic differentiation potential of human MSC and PDLSC, which might be related to the delayed healing processes seen clinically.
Nicotine exerts its cellular functions in stem cells through the nicotinic acetylcholine receptors [19,20]. While nicotine in tobacco contributes partly to the delayed fracture healing by inhibiting TNF-α secretion [10], the molecular mechanisms of nicotine effects are still largely unknown; especially as it relates to nicotine's effect on miRNA regulation on stem cell differentiation and proliferation. Nicotine has been reported to increase miR-140* expression in the rat PC12 cell line [32]. Moreover, nicotine downregulates miR-133 and miR-590 levels in canine atrial fibroblasts [33] and miR-146a expression in human placental TCL-1 cells [34]. In addition, upregulation of miR-16 and miR-21 was found in nicotine-treated human gastric cancer cells [35]. In this study, we, for the first time, reported the miRNA profile of nicotine-treated PDLSC (Table 1). Gene enrichment analysis suggested that the predicted miRNA-target genes could be involved in regulating cell migration, cell division, and bone development (Table 3). Furthermore, our miRNA predicted target genes and gene ontology analysis correlated well with the effects of nicotine in the biological studies performed. PDLSC behave similarly to MSC [14]; therefore, the miRNA expression and their related function in this study could pave the biological pathway for the investigation of nicotine effect in other kind of stem cells.
The function of hsa-miR-1305 in PDLSC was not known. In this study, we examined the expression of 2 hsa-miR-1305 target genes, PTK2 and RUNX2, which were selected from the predicted target gene list (Supplementary Tables S2 and S3). Both PTK2 and RUNX2 were shown to be downregulated in the nicotine-treated human MSC and PDLSC (Fig. 4). This suggested that the upregulation of hsa-miR-1305 could be associated with the downregulation of PTK2 and RUNX2, although these 2 genes could also be regulated by other miRNAs (Table 2) [36 –39] and miR-138 on PTK2 [40]. hsa-miR-30d and hsa-miR-137 were also upregulated in the nicotine-treated PDLSC (Table 1). Further analyses are needed to validate their direct interactions. Our data revealed the novel findings that the reduced stem cell migration and osteogenic differentiation by nicotine was related to the respective downregulation of PTK2 and RUNX2.
In summary, this study showed that the proliferation, migration, and osteogenic differentiation of human MSC and PDLSC were inhibited by exposure to nicotine. In addition, we also revealed the altered miRNA expression in nicotine-treated PDLSC, further providing evidence that miRNAs are a key regulator in these nicotine-associated functional changes.
Footnotes
Acknowledgments
This work was supported, in part, by the VA Merit Review Grant and the Senior VA Research Career Scientist Award, Miami, and a block grant of the University Grants Committee Hong Kong and the Endowment Fund for Lim Por-Yen Eye Genetics Research Centre, Hong Kong
Author Disclosure Statement
The authors indicate no potential conflicts of interest.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.