
Abstract
Ectopic expression of certain transcription factors induces reprogramming of somatic cells to a pluripotent state. A number of studies have shed light on the reprogramming capacity of various cell populations. As a result, it has been shown that stem/progenitor cells derived from organs of all germ layers exhibit a superior reprogramming efficiency compared to their differentiated progeny. Although proliferative capacity and endogenous expression levels of pluripotency factors are likely to be involved in this superiority, the detailed molecular understanding remains elusive so far. Recently, we have shown that the BAF-complex (BAF155 and Brg1), mediating epigenetic changes during reprogramming, is critical for the increased reprogramming efficiency of liver progenitor cells. In this review, we summarize recently acquired findings of the increased reprogramming capacity of adult stem/progenitor cell populations compared to their differentiated counterparts and discuss the potential mechanisms involved.
Introduction
However, a significant drawback of this technology for use in regenerative therapies is the potential risk of insertional mutagenesis associated with the genomic integration of lenti- or retroviral-based expression systems, potentially giving rise to cellular transformation and tumor formation [6]. In addition, generation of iPS cells is limited due to low reprogramming efficiency [1] (<1%) and longer duration [7] (several weeks) of reprogramming in contrast to nuclear transfer or cell fusion [8]. To bypass virus-mediated reprogramming, many efforts have been made to deliver reprogramming factors into target cells either by nonintegrating viruses [9,10], directly via mRNA transfection [11] or by recombinant proteins [12]. In addition, a defined set of small molecules has been shown to substitute for certain reprogramming factors [13]. Another quite robust approach is the excision of the exogenous DNA by, for example, recombinant cre-recombinases [14 –18]. Nevertheless, the overall efficiency was severely reduced compared to viral integration.
Beside the deficits of an optimal reprogramming platform, it has been hypothesized that the cell type of origin impacts the reprogramming process in terms of a bias toward the respective lineage [19,20]. Moreover, the reprogramming kinetics differs among the engaged cell types. Thus, the identification of certain subpopulations that are more amenable to cellular reprogramming and exhibit unbiased differentiation could shed light on the underlying process. Taken together, this groundwork should pave the way for the efficient generation of safe and ideal iPS cells.
Notably, adult stem cells share common features with their embryonic counterparts, such as extended differentiation potential and self-renewal [21,22]. In line with this, epigenetic modifications in adult- or tissue-specific stem cells more closely resemble those in ESCs compared to somatic cells [23 –27]. This is reflected by an enhanced reprogramming efficiency of tissue stem cells. In fact, others and we have shown that stem/progenitor cells from the skeletal muscle [28], the brain [29], the hematopoietic system, and the liver [30] are superior in cellular reprogramming compared to their differentiated counterparts. However, the intricate mechanisms orchestrating these effects require further exploration. Certainly, the endogenous expression of the reprogramming genes in combination with cellular proliferation rates play a definite role in the reprogramming process, but are not the only contributors to this phenomenon. Our own study has shown that BAF complex members, which are known to enhance reprogramming by chromatin remodeling of pluripotency loci, are highly expressed in liver progenitor cells (LPCs). The current review summarizes the efforts of reprogramming various adult stem cell populations and discusses the potential mechanisms underlying their highly efficient reprogramming capacities.
Stem/Progenitor Cells: The Ultimate Cell Type of Origin?
In recent years, various cell types were subjected to reprogramming. In particular, cell subtypes derived from the same organ were reported as being highly variable in their reprogramming capacity [28 –34]. In fact, accumulating evidence from systematic studies highlights that certain stem or progenitor cells in distinct tissues and compartments (muscle, blood, liver, skin, and brain), are more amenable to reprogramming compared to their more defined cellular counterparts. [28,30,31]. These findings will certainly aid the scientific community in their search for the perfect cell type for reprogramming and downstream applications and importantly, should provide deeper insights into the minefield, that is, reprogramming kinetics (Fig. 1).
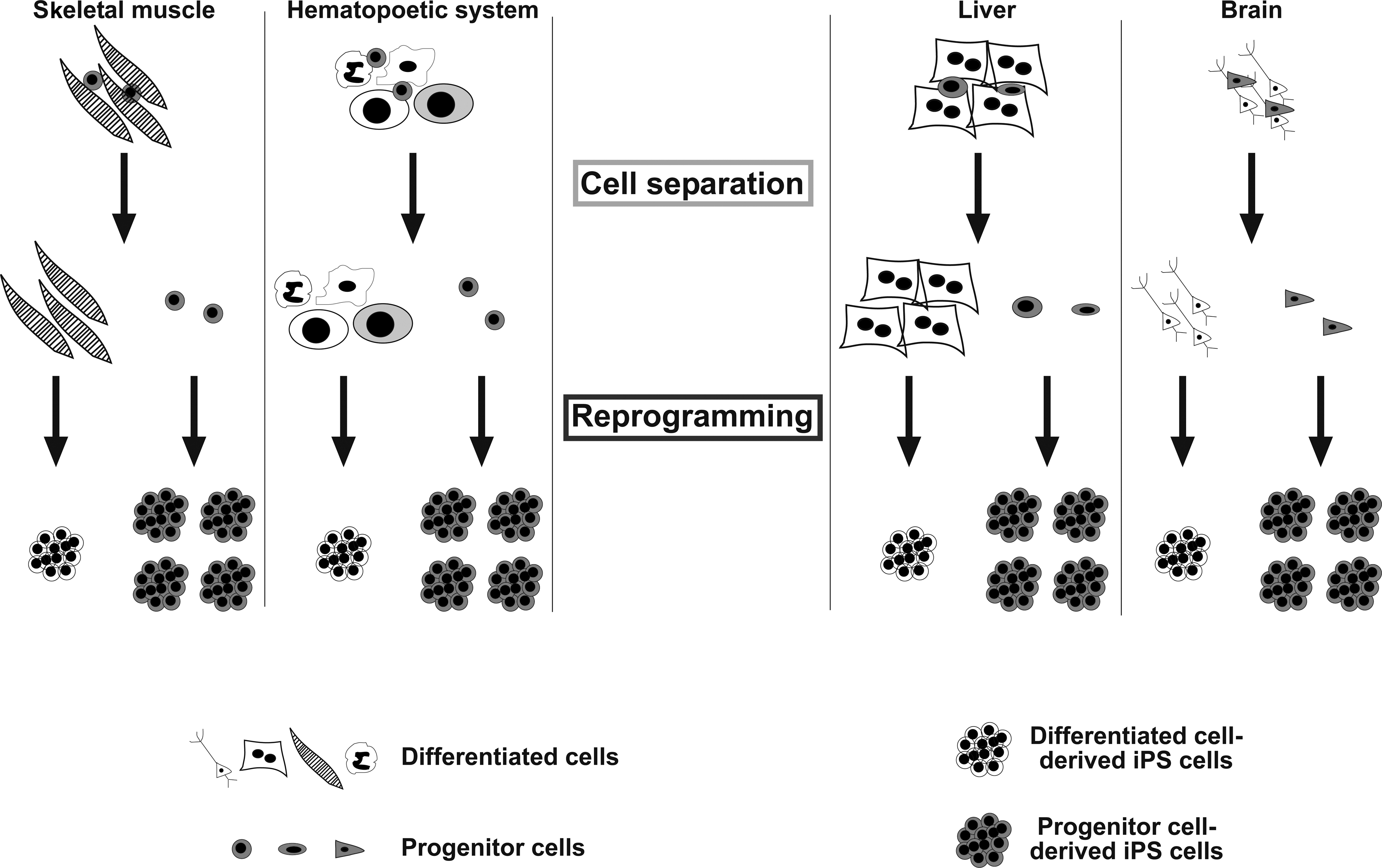
Model of a progenitor cell population within certain organ compartments, which is prone to reprogramming. The outline focuses on the 4 best-characterized compartments, skeletal muscle, hematopoietic system, brain, and liver. Small cells represent the stem/progenitor cells, which are more amenable to cellular reprogramming than their differentiated counterparts (black/white colors). To experimentally assess reprogramming capacity in a head to head comparison of both compartments, a separation step is required.
During mammalian development, stem cells are responsible for the specification of the embryo and tissue/organ development. Thereby, stem cells with varying differentiation capacity or potency are left behind in time. Stem cells, which reside in adult tissues mostly play a crucial role in tissue maintenance and repair, and are known as adult stem cells or somatic stem cells. They are distinctly localized and the surrounding microenvironment is defined as their particular stem cell niche within the respective organ. Nevertheless, adult stem cells from different tissues and different niches exhibit large variations in their differentiation potential and status [21,22]. These biological variations define their proximity to the most naive stem cells of all, ESCs, with respect to their proliferation capacity, epigenetic memory, and potency (Fig. 2).
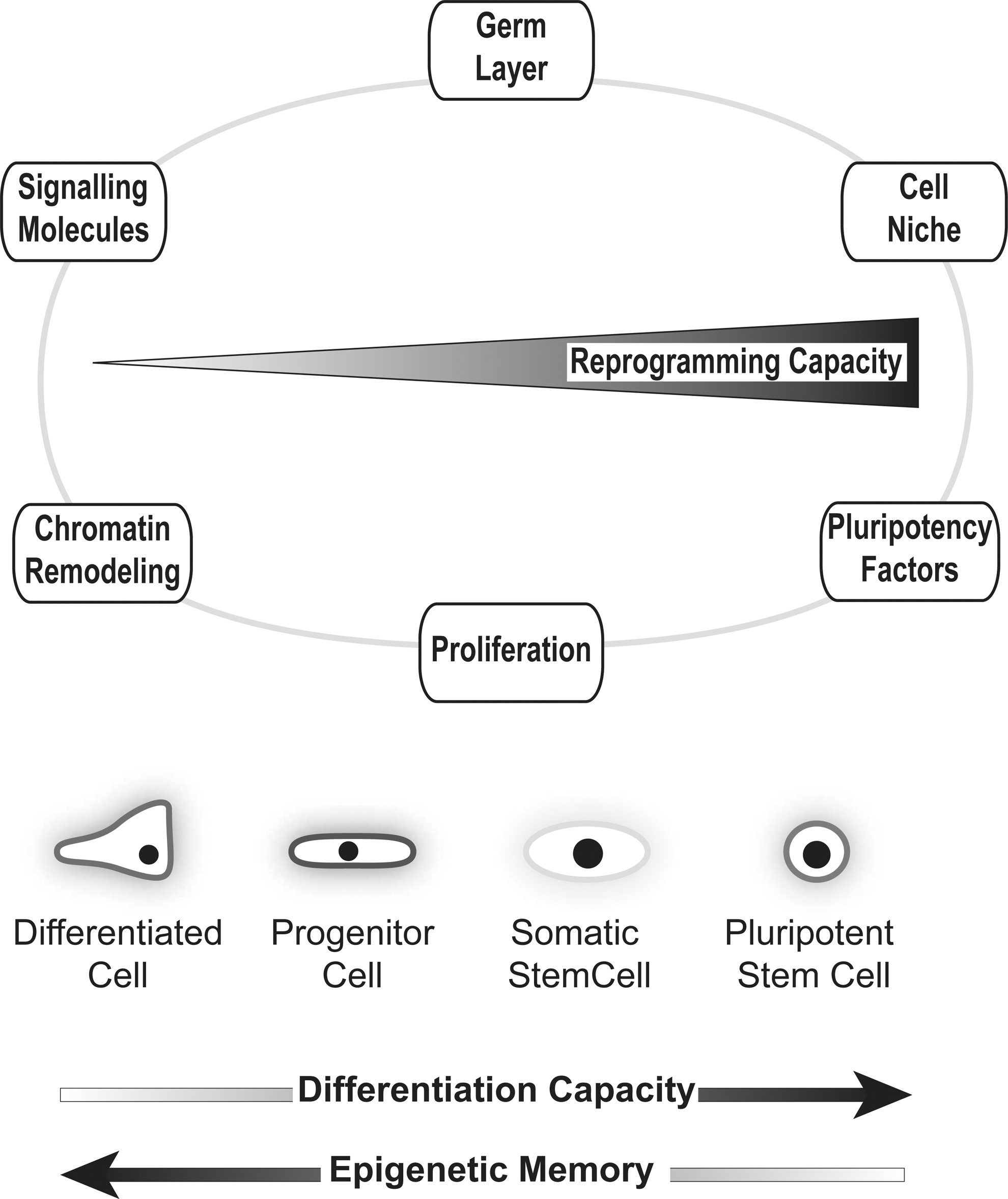
Different factors influence cellular reprogramming toward pluripotency. The circle represents the respective factors either positively or negatively influencing reprogramming. Below, the different cell types, pluripotent stem cells, adult stem or progenitor cells, and differentiated cells are shown. Kinetics in reprogramming capacity, differentiation capacity, and epigenetic memory are outlined by arrows in the figure.
Thus, the hierarchical organization of tissue stem cells, progenitor cells, transit amplifying cells, and their differentiated progeny could provide the blueprint for a cell in its propensity to regress back through the normal developmental stages (see also final paragraph). Another characteristic of adult stem cells is their endogenous expression of genes that are crucially involved in the pluripotency network (Fig. 2). It is still a matter of debate to what degree endogenously expressed factors of the pluripotency network act in concert during reprogramming and if successful reprogramming even depends on the relation of the targeted cell to the PSC. It has become clear that all 4 reprogramming factors possess unique abilities to drive or maintain pluripotency, yet, most known pluripotency factors exhibit various, context-dependent additional functions during cell differentiation and tissue development. This further points to the hypothesis, that the endogenously expressed factors may change their function during reprogramming to support the exogenously expressed ones [30]. Although pluripotency factors crucially regulate self-renewal, endogenous expression of stem cell factors in nonpluripotent cells does not necessarily indicate a proximity to ESCs. This has been shown for various core factors of the pluripotency network, such as Sox2 or Tbx3, which play a critical role for the maintenance of somatic stem cells, but in an entirely different context. However, to what extent the stemness of tissue stem cells impacts their capacity to give rise to iPSCs is poorly understood. Likewise, there is a lack of knowledge about the mechanisms of pluripotency itself.
So far, a limited number of tissue stem cells have been investigated in this context. The largest efforts to reveal the exact properties of adult stem cells have been undertaken in the brain, the blood, the skeletal muscle, and the liver (Fig. 1) [28 –34]. Another element necessary to consider when discussing reprogramming is the epigenetic memory of the differentiated cells. Epigenetic memory is based on DNA methylation and the Polycomb–trithorax group (Pc-G/trx) protein complexes at certain tissue-specific gene loci. Thus, the overall DNA methylation pattern of the genome defines a distinct differentiation and functional status of the cell [35]. While reprogramming a cell erases most of this methylation pattern, it has been shown that at least low passage iPS cells from individual tissues harbor particular DNA methylation signatures characteristic for their cell type of origin [26]. This epigenetic background or memory is also thought to be responsible for varying differentiation capacities among different iPS cell lines derived from various tissues [36]. Furthermore, it has been hypothesized and discovered that somatic cells like neural stem cells (NSCs) and dermal papilla cells possess epigenetic characteristics similar to that of the ES cells with regard to the demethylation patterns of the Sox2 and Nanog locus. These similarities within epigenetic status may also play an important role in making the cell more susceptible to reprogramming as these cells might require less remodeling of their epigenome during iPS cell formation [34,37]. Of note, although iPS cells derived from different organs and from cells with different maturation status (even of different ages) [38] show an incoherence of their differentiation capacities, the overall pluripotency and naivety is similar and even senescence is reversible [39]. It also became clear in recent studies that these capacity variations are more dependent on the cell donor than on the cell type reprogrammed [40]. Thus, deficiencies in DNA damage repair mechanisms appear to be an important underlying background for cell age-dependent differences in differentiation capacity of stem cells [38] and also for reprogramming efficiency.
The brain
Initially, in a series of studies, Schöler and colleagues reprogrammed NSCs from mice and men using different combinations of reprogramming factors [29,34,41]. In these studies, the endogenous expression of reprogramming genes in NSCs allowed the authors to omit certain factors from the reprogramming cocktail [29,34,41]. Indeed, the first gene, which could be easily withdrawn from the reprogramming combination, was c-myc. However, the omission of c-myc strongly reduced the reprogramming efficiency in fibroblasts compared to NSCs [41,42]. Another striking observation in NSCs was the high endogenous expression of Sox2, one of the master pluripotency genes, making it an additional candidate to be spared. Intriguingly, Sox2 expression not only plays a crucial role in the pluripotency circuitry by regulating the expression of Oct4, it later becomes important as a major regulator of neural tissue development and neural cell fate determination [43,44]. However, the kinetics and efficiency of NSC reprogramming in the absence of exogenous Sox2 were significantly reduced to 0.11% when compared to 3.6% with all the 4 reprogramming genes. This provided a good hint that endogenous Sox2 expression levels do not alone correlate with the reprogramming process, but still were sufficient for the successful reprogramming. The third factor that was withdrawn from the NSC reprogramming cocktail was the Krüppel-like factor 4 (Klf4). In PSCs, Klf4 functions both as a transcriptional repressor and activator to maintain pluripotency [36,45]. Being endogenously expressed in NSCs, although at much lower levels than in ESCs, Klf4 withdrawal still resulted in the generation of iPS cells with a reprogramming efficiency of 0.014%. Finally, the ultimate challenge to the reprogramming system is removal of Oct4. Until now, only a handful reports show that other factors can successfully replace Oct4 [46 –48] pointing to its critical role for the process itself. It is noteworthy that with the decreasing reprogramming efficiency due to less reprogramming factors, the latency of colony formation is also strongly increased. Intriguingly, NSCs reprogrammed using solely Oct4 (1 factor iPS cells) performed worse in a variety of differentiation assays [49], which lead to the hypothesis that levels and time frame of exogenously and endogenously expressed factors could change the integrity of the stemness circuitry (Table 1; Figs. 1 and 2).
These efficiencies contrast with fully differentiated postmitotic neurons, which reprogrammed with the efficiency of 0.8%, but required the expression of additional factors. More precisely, postmitotic neurons regained entrance to the cell cycle via repression of p53 together with the repression of lineage-specific genes [33]. This strongly underpins the hypothesis that a differentiated epigenome requires more effort and time for a complete reset. Obviously, this phenomena is also dependent on the differentiated cell type itself as not all fully differentiated cells require the additional factors for reprogramming [10,50 –53].
The blood
Similar, but more systematic data were provided for the hematopoietic system [31]. Technically, due to the inimitably well-characterized surface marker expression profiles of hematopoietic cells, also defining different maturation stages, it is possible to perform even single-cell reprogramming in a reasonably unbiased setting. A recent study combined these unique features of the hematopoietic system [31] with a genetically homogeneous secondary system to express the 4 reprogramming factors (Oct4, Sox2, cMyc, and Klf4; OKSM). In that respect, MEFs were infected with inducible virus constructs carrying the respective reprogramming factors and were subsequently reprogrammed into iPS cells. In a further step, mice were generated from these iPS cells resulting in the genomic existence of the inducible exogenous factors in every cell of the body. Hematopoietic cells harvested from these animals, could finally be secondarily reprogrammed, simply by activation of the constructs. Based upon surface marker isolation of mature and immature cells and comparing the reprogramming kinetics, an inverse correlation between the differentiation stage and reprogramming capacity was noted as hematopoietic stem cells (HSCs) performed even slightly worse in reprogramming compared to myeloid progenitors. Even though, the HSCs reprogrammed worse than their differentiated counterparts, the overall reprogramming was still strikingly higher with 7%–28% in the undifferentiated cells compared to 0.02%–0.6% in the differentiated cells (Table 1). Given the simplicity of isolating myeloid progenitors or HSCs by either a bone marrow puncture or a simple blood withdrawal after granulocyte colony-stimulating factor stimulation and subsequent surface marker staining, the blood system is likely to be one of the best candidates for making benefit of a potential organ hierarchy in reprogramming. It would be of particular interest to learn whether these differences are simply based on lineage-specific signatures in the differentiated blood cells, which are harder to erase, or whether there are particular factors present in the stem/progenitor cell compartment, which facilitate reprogramming just as BAF complex factors do in LPCs (paragraph below). Our reanalysis of published transcriptome sets at least pointed toward certain factors being present in undifferentiated hematopoietic cells compared to T cells, which share similar transcriptional distances from ESCs, but show much worse reprogramming efficiencies than HSCs (Fig. 3).
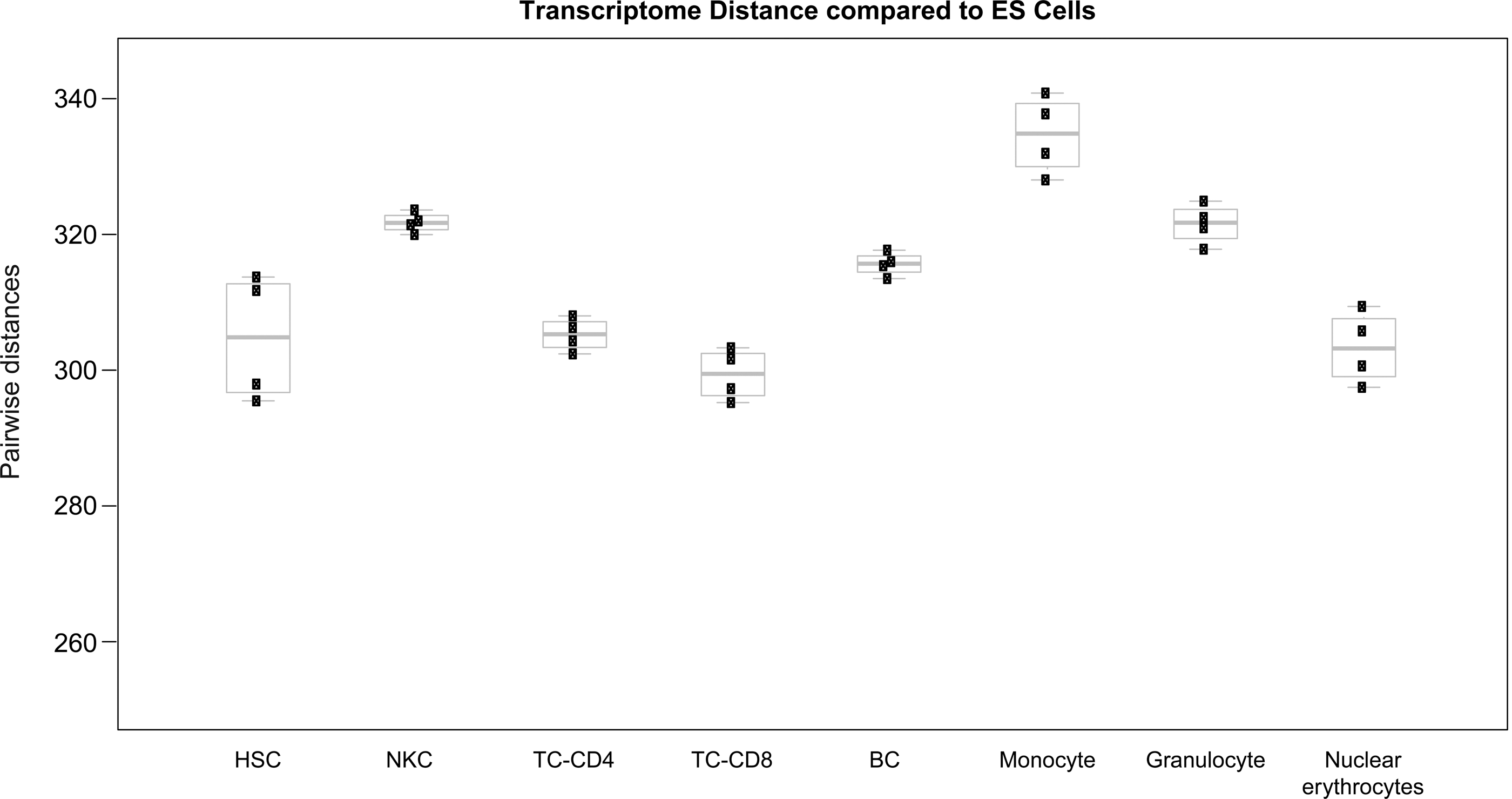
Pairwise distance calculation of different hematopoietic cell types from established embryonic stem cells upon whole transcriptome profiling. Note that there is no direct connection between transcriptional distance from ES cells and reprogramming capacity [31]. Abbreviations are as follows: Hematopoietic stem cells (HSC); natural killer cells (NKC), B cells (BC), T cells (TC-CD4, TC-CD8), monocytes, granulocytes, and nucleated erythrocytes. The difference in gene expression of the various hematopoietic cell types from ES cells is shown as boxplots, which give pairwise Euclidian distances of gene expression profiles between the respective groups. Data sets were downloaded from GEO (
The skin
Another progenitor cell compartment, which was subjected to cellular reprogramming were dermal papilla cells, a specialized mesenchymal cell type thought to instruct epithelial stem cells during hair morphogenesis and regeneration. This cell type was successfully reprogrammed with as few as 1 factor, namely Oct4. Again, it became clear that their reprogramming capacity exceeds the one ascribed to more differentiated cells (1.38% using 4 factors; Table 1). However, the authors did not perform a head to head comparison of the undifferentiated cells to their differentiated counterparts using the same culture conditions or reprogramming combinations [37,54]. This head to head comparison is generally hampered by the lack of surface markers defining distinct, but different cell types in a particular organ. In fact, the authors used a fluorescent reporter labeling the dermal papilla cells for their isolation. Nevertheless, the current knowledge points toward an intrinsic reprogramming capacity of tissue stem cells. Looking at a cell type that is not a stem cell type, but is located in the same niche as dermal papilla cells, hair keratinocytes from the hair outer root sheath, have not only been shown to exhibit a great reprogramming efficiency, but also express the factor Klf4 at high levels [55 –57]. Surprisingly, these cells could not be successfully reprogrammed upon omission of Klf4 [56].
The muscle and the liver
Two other microenvironments, namely, the muscle [28] and liver [30] have been thoroughly investigated in a more comprehensive manner. The advantages of both studies are given by the facts that (i) culture conditions for differentiated and undifferentiated cells have been characterized extensively and that (ii) a set of cell surface markers discriminating the respective populations exist [58 –61]. Thus, using a defined secondary system, Wagers and colleagues could show that unipotent skeletal muscle precursors and bipotent nonmyogenic mesenchymal progenitor cells gave rise to iPS cells with an efficiency of around 23%–25% which by far exceeded the one from the differentiated myogenic compartment [28]. Again, the authors investigated the endogenous expression of c-myc and Klf4 and found both expressed in the stem and nonstem cell compartment. However, the expression pattern could not explain the reprogramming differences (Table 1).
The hitherto described studies focused mainly on mesodermal- or ectodermal-derived stem cell compartments. In consequence, our study concentrated on an endoderm-derived solid organ, namely, the liver. Beforehand, hepatic progenitor populations have been identified in fetal and adult mouse liver tissue [58,59,62 –64]. Thus, similar to the hematopoietic system, it became possible to isolate LPCs based on the expression of distinct surface markers. This subset of cells exhibits a bipotent differentiation potential in cell culture experiments, generating cells of the hepatic and biliary lineage. For comparative purposes, we also isolated differentiated hepatocytes according to standard methods from livers of the same animals to allow a precise head to head comparison in the reprogramming capacity of differentiated and undifferentiated liver cells [30]. Using equally titered polycistronic viruses encoding 3 and 4 reprogramming factors, we were able to reprogram LPCs with efficiencies of 22% (OSK) and 30% (OSKM), respectively. These reprogramming efficiencies were significantly higher compared to those of differentiated hepatocytes, namely, 0.08% (OSK) and 0.34% (OSKM) (Table 1). Moreover, we assessed the reprogramming kinetics of LPCs and differentiated cells and found a pronounced acceleration in the whole process showing fully reprogrammed iPS cell colonies derived from the liver progenitor cells as early as 3 days after infection. The grade of reprogramming was evaluated upon reactivation of the endogenous Oct4 locus based on a reporter allele [30]. In both studies, muscle and liver progenitors were fluorescent activated cell sorting (FACS) purified. However, cell reattachment after FACS purification does not occur with absolute efficiency. Thus, it is likely that reprogramming efficiencies were even underestimated bearing in mind that usual plating efficiency ranks around 20% for both FACS sorted muscle and LPCs [28,30].
To date, there are several organs and systems, which allow a systematic dissection of a reprogramming hierarchy, based on robust cell separation assays, for example, blood, skeletal muscle, and liver. We summarize the different types of progenitor cells used in the above-mentioned approaches, the various ways used to reprogram these cells, and the obtained efficiencies in a tabular format (Table 1). Understanding why progenitor or adult stem cells perform superior in reprogramming will help answering the following important questions: (i) which mechanism decides about the enhanced reprogramming process in stem cells? (ii) Is a globally shorter distance between progenitor cells and pluripotent cells responsible or are there particular factors responsible for this phenomena (Figs. 1 and 2)? In summary, further systematic and global analyses in defined settings within the same organ will help us to gain deeper insights into the reprogramming process. We have tried to address, at least in part, these issues by a deeper analysis of LPCs to identify factors ascribing their reprogramming fitness and to investigate how proliferation impacts on the process. Thus, the following 2 paragraphs summarize and discuss the current knowledge of proliferation and other reprogramming factors in terms of their impact on progenitor cell reprogramming.
Nothing Else, but Proliferation?
Recent findings provide evidence that reprogramming requires a series of stochastic events that are highly dependent on the simple rate of cell proliferation [7,65]. Live-cell imaging revealed that reprogramming correlates with an indicative change in cell morphology occurring within one round of cell division after exogenous expression of pluripotency genes [66]. Based on these studies, it has been proposed that nearly every cell can give rise to an iPS cell via a stochastic process, which is determined by 2 critical parameters, namely, the time frame of cell division and mechanisms being independent of proliferation [7]. Pluripotent cells are defined by a high number of cells in S-phase with a short G1-phase in their cell cycle profile [67]. Thus, a cell type resembling this cell cycle profile is more likely prone to reprogramming than others. Jaenisch and colleagues explained these differences in proliferation as changes in the reprogramming time until colony formation. This time has been termed as latency. By means of a genetically homogenous reprogramming system, they postulated that in case of a reprogramming process exclusively driven by proliferation, a doubling in the proliferation rate leads to an ∼2-fold shortening in latency [7].
However, proliferation and reprogramming are vaguely connected. In fact, a series of studies, including our own, have shown that proliferation does not uniquely impact on the reprogramming process [30,68]. This seems to be particularly true in the case of stem cells. Assuming the above-described scenario that proliferation-dependent events lead to a delayed, but similar efficient reprogramming of cell types over time [7,65], we performed analyses to calculate the reprogramming latency for LPCs and their differentiated counterparts [30]. We observed a more than 10-fold increased latency for differentiated liver cells. Since proliferation rates in LPCs were only increased by 15%–20% compared to non-LPCs, proliferation rates unlikely accounted exclusively for these differences in reprogramming. In addition, we simulated a doubling in proliferation rates in the differentiated liver cells, but were still not able to make the curves collapse (Fig. 4, adopted from [30]). A similar, but more biological approach has been used to modify the proliferation of HSCs. Hochedlinger and colleagues cultured HSCs either in the quiescent stage or under cycling conditions by the addition or removal of particular cytokines. Afterward, they assessed the reprogramming capacity and found virtually no difference [31]. Thus, at least for the liver and blood system, proliferation is likely to have a relatively low impact on the reprogramming fitness of the stem/progenitor cells.
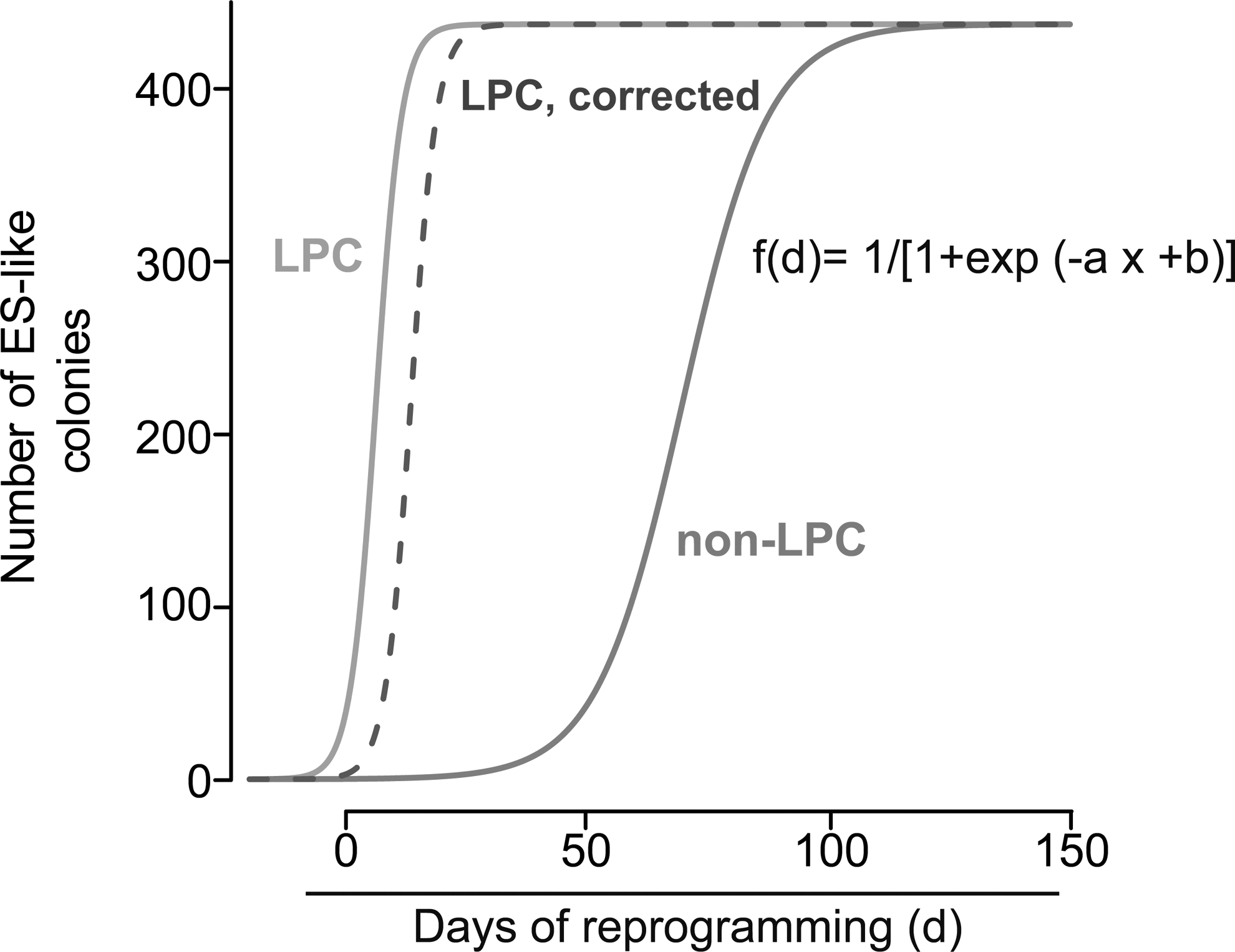
Latency calculation for liver progenitor cells (LPCs) and non-LPCs. Assuming that stochastic events would lead to a delayed, but similar efficient reprogramming of both LPCs and non-LPCs over time [7,65], curve fitting was used to calculate the latency for LPCs and non-LPCs to reach the plateau of maximum reprogramming based on the observed data (for details see also [30]). All experimental data were fitted (R version 2.11, function nls) with sigmoidal functions of the form: f(d)=1/[1 + exp(−a x + b)]. Parameter values were a=0.372, b=2.311 (LPCs, light gray) and a=0.1116, b=7.7868 (non-LPCs, dark gray). Inflection points were calculated to be at day 6.21 and day 69.77. Light gray line: reprogramming kinetics of LPCs. Dotted line: predicted effect of a 50% decrease in cell proliferation on the delay in reprogramming of LPCs. Dark gray line: calculated reprogramming kinetics of non-LPCs based on the measured data (from Kleger et al., [30]).
However, increased proliferation rates could also be indirectly linked to improved reprogramming by the expression of cell cycle-associated genes acquiring alternative functions in a reprogramming context. One of these factors, serving as a critical driver of the cell cycle machinery, is c-myc. On the one hand, the oncogene c-myc strikingly improves iPSC formation, but on the other hand, it significantly increases tumor formation in iPSC-derived chimeric mice. Thus, c-myc could play a dual role during reprogramming: First, the suppression of differentiation-associated genes and, second, the activation of genes being expressed in highly proliferative cells, including cancer cells, iPSCs, and ESCs [69]. However, the knowledge about the functional role of c-myc remains underdeveloped as the reprogramming kinetics and efficiencies strongly differ upon omission of c-myc in different cell types. For example, using MEFs, the overall efficiency was reduced between 1 and 2 orders of magnitude and kinetics were 5-fold delayed when c-myc was not exogenously expressed [42]. Moreover, endogenous c-myc levels of liver progenitors and dermal papilla cells already strongly exceed those of MEFs and even ESCs. In line, when c-myc was omitted from the reprogramming cocktail of liver progenitors, the reprogramming efficiency just slightly dropped (30% to 22%) and the kinetics remained virtually similar compared to 4-factor reprogramming. Hypothetically, endogenous saturation of c-myc expression in LPCs allowed high reprogramming continuously even without exogenous c-myc. Alternatively, in the context of reprogramming, c-myc clears the reprogramming path for other genes instead of simply keeping up cell division to increase the probability of reprogramming (Fig. 2).
In summary, higher proliferation rates certainly have an impact on the reprogramming capacity of particular cell populations. Nevertheless, our mathematical modeling data [30], published biological data [30,31], and various other facts [7,65,66] point to a reprogramming fitness in adult stem cells being mainly independent of proliferation. Thus, the responsible endogenous factors that are expressed in certain stem cell populations have to be identified.
BAF Complex and Potential Other Reprogramming Mediators
So far, we have discussed the influence of the OKSM reprogramming factors and proliferative capacity on the reprogramming fitness of different stem/progenitor cells. However, the current knowledge points toward additional mechanisms. At this point, one could envision 2 potential scenarios: (i) The transcriptome of stem/progenitor cells regenerating different organs share global signatures with pluripotent cells, thus, leading to a shorter epigenetic reset process. (ii) There are cell-intrinsic, molecular predicates such as specific, endogenously expressed factors in stem/progenitor cell populations helping in faster transition to the iPS state. These mechanisms act either in concert or independently of the OKSM reprogramming cocktail to reset the epigenome toward a pluripotent stage. The first scenario could be particularly present in stem cells, which share common factor expression signatures with the core pluripotency network such as NSCs do. In NSCs, there is a possibility that Sox2 switches its function while being already expressed endogenously, thus, sparing the time of resetting the locus for pluripotency. However, a precise analysis for this hypothesis is lacking so far and, therefore, remains a matter of speculation. From the above-discussed reports, as yet, only our study aimed for deeper insights into the mechanism of stem cell reprogramming fitness. To this end, we performed pairwise distance calculation within the differentiated liver cells and the less differentiated LPCs from pluripotent ESCs. Intriguingly, and to our surprise, we found a significantly greater distance in LPCs versus ESCs compared to non-LPCs versus ESCs. No matter how large these differences are, everything above equal transcriptional distance points toward the second hypothesis, namely, cell-intrinsic, molecular regulatory devices being present in LPCs, thereby mediating reprogramming fitness. Comparable results can be obtained dissecting the transcriptome of various hematopoietic cells starting from the stem cell level (Fig. 3). Obviously, no significant difference in the proximity to the ES cell transcriptome is present when HSCs are compared with differentiated immune cells of both the myeloid and lymphoid lineage. Indeed, T-Lymphocytes of different classes exhibit a signature, which is in closer proximity to ESCs, compared with all other hematopoietic cells.
In consequence, our search turned toward other factors regulating reprogramming fitness. So far, the following classes could be discriminated: (i) Transcription factors, which act downstream of one of the OKSM genes, thereby substituting an OKSM factor and/or enhancing the process. (ii) Signaling molecules, which activate one of the pluripotency pathways, such as Lif/Stat3 signaling, thereby enhancing the activation of the core pluripotency network. (iii) Molecules, which pioneer the path for the reprogramming factors, such as chromatin remodeling complexes, thereby shifting the balance from densely packed heterochromatin to loosely packed euchromatin at certain pluripotency loci.
The latter was addressed in a recent study from the Schöler laboratory [70]. They showed for the first time an elaborated picture in which the epigenetic machinery takes part in iPS reprogramming. In a large proteomics approach, they identified the chromatin remodeling SWI/SNF complex (ATP-dependent BAF chromatin-remodeling complex) to heavily influence somatic cell reprogramming. Interestingly, BAF complex-driven reprogramming was particularly obvious when c-Myc was omitted from the reprogramming cocktail. This underpins our data reporting still pronounced reprogramming efficiencies in LPCs upon c-Myc omission [30].
Mechanistically, the BAF complex has been reported to interact with DNA methylating enzymes [71 –73], thereby ascribing it the ability to modulate the methylation status of crucial stem cell genes. In the context of reprogramming, the SWI/SNF complex may regulate DNA methylation either directly—that is, by recruiting DNA methylases—or indirectly—that is, by altering the expression of other proteins affecting DNA methylation. For instance, Brg1, required for nuclear reprogramming, has been shown to interact with the DNA methylation enzyme Dnmt3a, a protein driving HSC differentiation in lymphosarcoma cells [71,74]. Also, the knockdown of Brg1 has been known to negatively impact on the levels of Oct3/4. Additionally, it was shown that esBAF complex components achieve a euchromatic chromatin state and enhance binding of cell state-specific transcription factors to their respective DNA loci, among them the binding of reprogramming factors onto key pluripotency gene promoters, thereby enhancing reprogramming [70,75 –77]. In former studies, it had already become evident that members of the BAF complex facilitate the expression of reprogramming genes, such as c-Myc by the interaction mediated between c-Myc and Ini1, a further component of the BAF complex [72,73,78,79]. esBAF also plays an important role by regulating Stat3 signaling thereby controlling LIF and BMP signaling pathways, which in turn are required for maintenance of self-renewal and pluripotency [73]. The other important component of the BAF complex, BAF 155, may enhance reprogramming by replacing baf170 thereby converting the endogenous BAF complex to an esBAF complex, which in turn facilitates chromatin remodeling [70]. In this context, they interact with binding sites of the respective genes within the genome and initiate the demethylation process [70]. This is underlined by the fact that the loss of the BAF complex leads to an abnormal hyper-methylation of these loci [70].
In our context, the BAF complex shows robust expression in self-renewing tissues, including the liver [80]. Intriguingly, Brg1 is differentially expressed during liver development showing a decline with the differentiation stage from fetal to adult liver [75]. Moreover, targeting Brg1 via RNAi revealed diminished albumin transcription [75]. Given this exciting function of Brg1 and other BAF complex members during pluripotency and liver development, we hypothesized the BAF complex as the potential driver of reprogramming fitness in LPCs. Gene expression analysis showed differential expression of Brg1 and BAF155 with a pronounced expression in adult and fetal LPCs compared to non-LPCs [30]. Reanalyses of recently published transcriptome data on LPCs isolated from adult mice confirmed these data reporting an increased BAF complex expression [81,82] compared with differentiated cells. The conserved pattern in adult/fetal liver points to a particular function of the BAF complex in LPCs and consequently for the differentiation/regeneration of the liver. To rigorously test the hypothesis that the BAF complex members Brg1 and BAF155 are the critical drivers of reprogramming fitness in LPCs, we knocked down both factors in LPC and non-LPC compartments. Interestingly, the reprogramming efficiencies of the LPCs resembled those of the differentiated liver after knockdown of the BAF complex. These dramatically reduced reprogramming efficiencies pointed toward a model (Fig. 5) where the BAF complex members govern the transcription of liver/LPC-specific genes, such as albumin, by keeping cell state-specific chromatin regions in an open state. However, after the application of the reprogramming factors, endogenous BAF complex members switch their function to an esBAF function to promote an ESC-specific transcription by enhancing binding of the reprogramming factors to key reprogramming gene promoters [70]. Given this intimate role of the BAF complex for reprogramming, it would be interesting to assess its function during human cell reprogramming. Human iPS cells are in closer proximity to the primed than to the naïve pluripotent state. In fact, this more epiblast-like state represents a major hurdle of human PSC research as certain applications, such as homologous recombination, occur less frequently. Future experiments must determine whether BAF-mediated human cell reprogramming could shorten this distance to the naïve state to overcome at least some obstacles of the primed state.
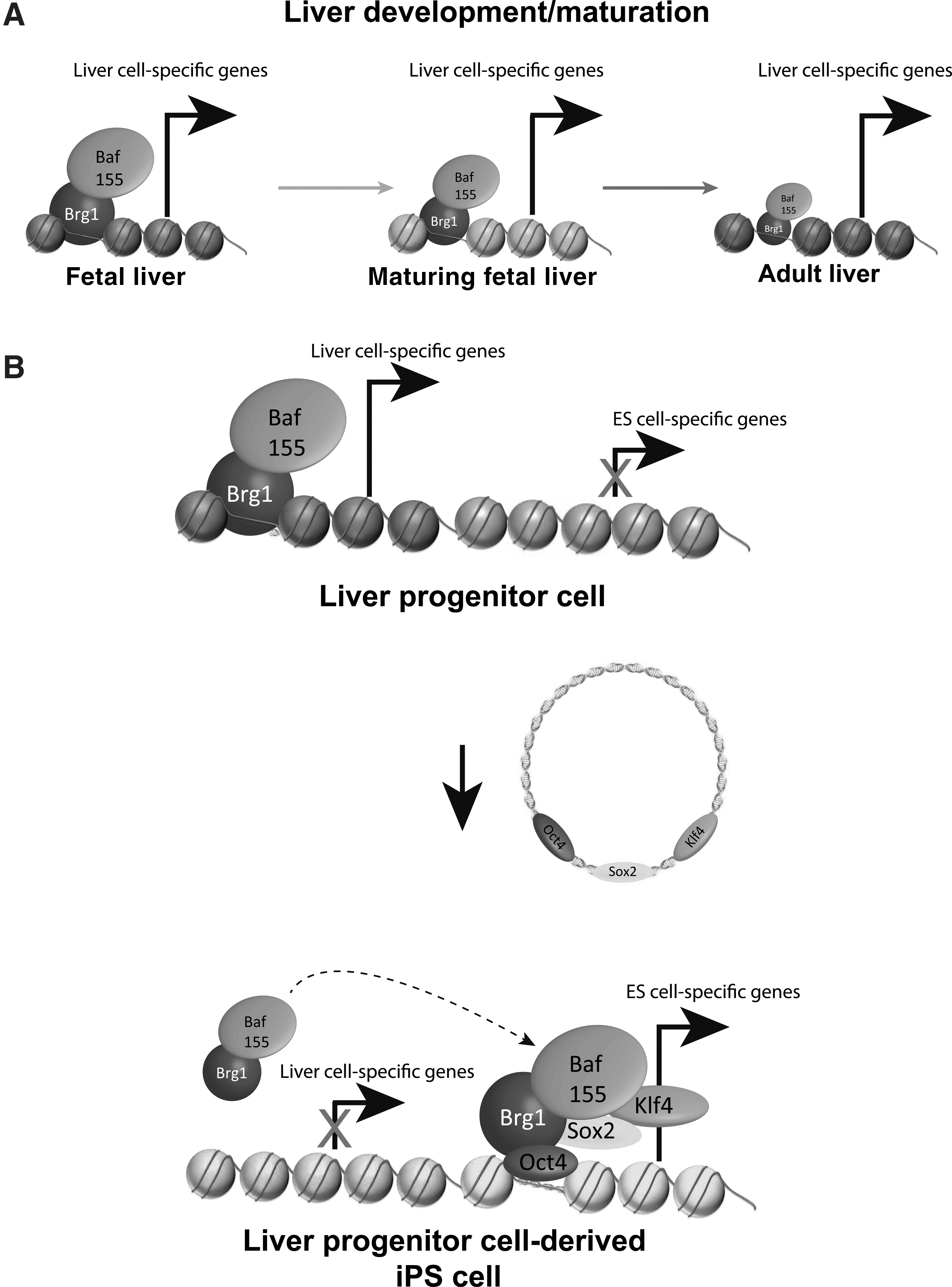
Assumed BAF complex expression behavior during liver development and LPC reprogramming.
In summary, this clearly strengthens previous observations in different types of donor populations that the epigenetic status of a particular cell type facilitates the higher reprogramming efficiencies by creating a favorable epigenome for the binding of stem cell genes. As adult stem cells are generally richer in euchromatin [23 –26,83], BAF-complex factors may define a novel molecular cornerstone in somatic stem cells, contributing to the increased reprogramming capacity of adult stem cells in general. These findings could also be relevant for the dedifferentiation of somatic stem cells during cancer formation.
Outlook
Tissue stem cells represent an attractive cell source for the generation of iPS cells as they carry several advantages over their differentiated counterparts. In almost all studies, including our own, it was observed that tissue stem cells show a higher degree of reprogramming efficiency compared to their differentiated progeny, which makes them a more suitable cell source for reprogramming. Additionally, at least in the hematopoietic system, cells are easily obtained.
Origin of a donor population really matters in the case of reprogramming efficiency due to the fact that donor cells from a particular niche show a characteristic gene expression profile, a specific epigenetic status and a certain level of endogenous expression of reprogramming factors. A variety of studies have been performed using a vast array of donor cells from different niches and most of these studies significantly correlated the origin of the donor cell to their reprogramming potential (Fig. 1). These controlled differences in reprogramming ascribed by endogenous variables allow 4 major conclusions at this point: (i) Adult stem and progenitor cells from different organs are likely to be the optimal cell type of reprogramming origin due to higher efficiencies, fewer necessary factors and less time required to reach the pluripotent state. (ii) Systematic analysis will allow the identification of new and endogenous factors (niche dependent and independent), which could drive reprogramming. (iii) BAF complex factors are differentially regulated chromatin remodelers, which could account for endogenous differences in reprogramming capacity independent of OKSM expression. (iv) Adult stem cells provide a platform, which allows the investigation of most of the relevant factors influencing reprogramming, such as differentiation stage (mature vs. immature), proliferation (high vs. low proliferation), niche (environmental impact), epigenetic memory, and germ layer affiliation (Fig. 2).
Footnotes
Acknowledgments
AK is supported by a fellowship provided by the Medical Faculty of Ulm University (Bausteinprogramm, L.SBR.0011) and a fellowship provided by the German Cardiac Society. This study was funded by the Deutsche Forschungsgemeinschaft (DFG, KL 2544/1–1, SL BO1718/4-1), the German Foundation for Heart Research (F/34/11; to AK and SL), the Else-Kröner-Fresenius-Stiftung (2011_A200; to AK and SL), Boehringer-Ingelheim BIU (to AK and SL), the Helmholtz Gesellschaft (VH-VI-510 to SL), and BMBF (MND-Net to SL). AK is indebted to the Baden-Württemberg Stiftung for support by the Elite program for postdocs. This work was funded in part by the German Federal Ministry of Education and Research (BMBF) within the framework of the program of medical genome research (PaCa-Net; project ID PKB-01GS08) to HAK, and the framework Gerontosys II (Forschungskern SyStaR, project ID 0315894A) to HAK and AK/TS.
Author Disclosure Statement
There is no potential conflict or competing interest to disclose.