
Abstract
Human embryonic stem cell (hESC)-derived oligodendrocyte progenitor cells (OPCs) are being studied for cell replacement therapies, including the treatment of acute spinal cord injury. Current methods of differentiating OPCs from hESCs require complex, animal-derived biological extracellular matrices (ECMs). Defined, low-cost, robust, and scalable culture methods will need to be developed for the widespread deployment and commercialization of hESC-derived cell therapies. Here we describe a defined culture system that uses a vitronectin-derived synthetic peptide acrylate surface (VN-PAS; commercially available as Corning® Synthemax® surface) in combination with a defined culture medium for hESC growth and differentiation to OPCs. We show that synthetic VN-PAS supports OPC attachment and differentiation, and that hESCs grown on VN-PAS are able to differentiate into OPCs on VN-PAS. Compared to OPCs derived from hESCs grown on ECM of animal origin, higher levels of NG2, a chondroitin sulfate proteoglycan expressed by OPCs, were observed in OPCs differentiated from H1 hESCs grown on VN-PAS, while the expression levels of Nestin and PDGFRα were comparable. In summary, this study demonstrates that synthetic VN-PAS can replace complex, animal-origin ECM to support OPC differentiation from hESCs.
Introduction
A
Interactions with extracellular matrix (ECM) have been shown to affect OPC survival, proliferation, migration, and maturation [15]. Virtually, all protocols to differentiate OPCs from hPSCs include culture of progenitor cells on ECM-coated surfaces [7,8,16,17]. The ECM proteins laminin and fibronectin have been used for OPC differentiation, either alone or in combination with poly-D-lysine, as has Matrigel™, a complex ECM derived from the murine Engelbreth-Holm-Swarm with laminin as the main constituent [7,8,16,18,19]. Laminin is a potent promoter of oligodendrocyte survival and myelination whose receptor α6β1 integrin is expressed on OPCs and plays a critical role in mediating growth factor signaling to control oligodendrocyte development [20,21]. Recently, vitronectin, a multifunctional ECM glycoprotein that is expressed in the ventral part of developing human spinal cord, has been shown to promote oligodendrocyte differentiation from hESCs by synergistically interacting with sonic hedgehog protein [22].
ECMs have also been extensively investigated for the culture of undifferentiated hPSCs. Various complex ECMs, including Matrigel, human serum, and a mixture of multiple ECM proteins, were found to support the propagation of hESCs in culture [23 –25]. This was followed by the identification of purified ECM proteins that support hPSC growth and expansion, including laminin, fibronectin, or vitronectin [26 –29], and the recombinant versions of these proteins (i.e., laminin 511, laminin 111, and vitronectin) [30 –33]. Perhaps the most promising advancements for eventual widespread clinical use of hPSCs have been the recent development of defined peptide and synthetic polymer surfaces that support hPSC expansion [34 –38]. These surfaces are of defined composition, devoid of animal origin materials, and potentially scalable for commercial manufacture of hPSC-derived cell products; however, little is known about the capacity of lineage-specific differentiation for hPSCs grown on these surfaces. Characterization of the defined surfaces is critical because small differences in the culture systems may have significant impact on the ability of hPSCs to differentiate into the desired cell populations [39,40]. In our previous study, synthetic peptide acrylate surface (PAS), one of the few surfaces able to support long-term self-renewal of hESCs in a defined medium, was shown to support hESC differentiation into cardiomyocytes [38]. However, to date, no synthetic surface has been reported to support OPC differentiation from hPSCs.
In this study, the ability of vitronectin-derived synthetic peptide acrylate surface (VN-PAS, commercially available as Corning® Synthemax® surface) to support the differentiation of hESCs to OPCs was evaluated. VN-PAS is shown to support OPC differentiation of hESCs maintained on either a complex ECM (Matrigel) or VN-PAS. The combination of VN-PAS and Ultra-low Attachment surface, together with defined media, provides an integrated, defined culture system for hESC expansion and OPC differentiation. To our knowledge, this is the first report that describes the use of a synthetic, vitronectin peptide-containing surface to support OPC derivation from hESCs as well as the use of a defined culture system for both hESC expansion and the subsequent OPC differentiation.
Materials and Methods
Human ESC culture
H1 hESCs were cultured under feeder-free conditions as described previously [27,41]. Briefly, cultures were passaged by incubation in 200 U/mL collagenase IV (Invitrogen) for 5–10 min at 37°C, mechanically dissociated, and seeded as clumps onto T-75 flasks coated with 1:30 diluted growth factor-reduced (GFR) Matrigel (BD Biosciences) or VN-PAS T-75 flasks (Corning Synthemax Surface; Corning). The cells were maintained in X-VIVO™ 10 medium (Lonza) supplemented with basic fibroblast growth factor (bFGF, at 80 ng/mL; Invitrogen) and transforming growth factor-β1 (TGFβ1, at 0.5 ng/mL; R&D Systems). For this study, hESCs from a cell bank cryopreserved at passage 30 were used and the cells were cultured for up to 20 passages after thawing. Triplicate T-75 flasks were maintained for each condition with one flask sacrificed for cell counting using a Vi-Cell XR analyzer (Beckman Counter) after dissociation with 0.05% trypsin/EDTA. The seeding density was ∼1×105 cells/cm2 based on a cell count obtained from a parallel T-75 flask. The cells were passaged every 5–7 days. The seeded cell numbers and the harvested cell numbers at each passage were used for the calculation of apparent doubling time.
Vitronectin-derived synthetic peptide acrylate surface
The detailed procedure for VN-PAS production is described elsewhere [38] and VN-PAS vessels are commercially available as Corning Synthemax Surface from Corning Incorporated. Synthemax Surface is a synthetic surface comprised of an acrylate polymer conjugated to the biologically active vitronectin peptide Ac-KGGPQVTRGDVFTMP. For this study, T-75 flasks with Synthemax Surface were used.
OPC differentiation
H1 hESCs grown on GFR-Matrigel-coated T-75 flasks or Synthemax T-75 flasks were differentiated into OPCs according to a 41-day protocol with some modifications [7,9]. Briefly, the cells were dissociated into small clumps using collagenase IV treatment plus mechanical scraping. The cells formed embryoid bodies (EBs) and cultured in Ultra-low Attachment T-75 flasks (Corning) for 27 days followed by 14 days of adherent culture. On day 0, cell clumps were cultured in Transition Media (50% differentiation medium and 50% XVIVO 10) with 4 ng/mL bFGF and 20 ng/mL epidermal growth factor (EGF; Sigma-Aldrich). The differentiation medium consists of DMEM/F12 (Invitrogen), 2% B27 (Invitrogen), and 40 ng/mL triiodothyronineT3 (Sigma-Aldrich). On day 1, about 10 μM all-trans retinoic acid (RA; Sigma-Aldrich) was added to Transition Medium together with 2 ng/mL bFGF and 20 ng/mL EGF. During days 2–8, EBs were fed daily with differentiation medium supplemented with EGF and RA. From day 9 to 26, EBs were fed 3 times a week in the presence of EGF only. No cell dissociation was performed during days 1–26. On day 27, EBs were transferred at 1:2 ratio onto GFR-Matrigel-coated T-75 flasks or Corning Synthemax T-75 flasks and cultured for 7 days. The plated cells were dissociated using 0.05% trypsin/EDTA (Invitrogen) and reseeded at 3×106 per T-75 flasks. After another 7 days, OPCs were harvested with 0.05% trypsin/EDTA and cryopreserved. Cell counting on days 34 and 41 was performed using a Vi-Cell XR analyzer (Beckman Counter). The OPC doubling ratio was calculated as the cell number at day 41 divided by the cell number at day 34.
Flow cytometry
Live cells were analyzed for surface markers mouse IgG1 anti-NG2 (Zymed) and mouse IgM anti-Tra1-60 (Millipore). Cells stained with the DNA-intercalating dye ethidiummonoazide bromide (EMA; Sigma-Aldrich), followed by paraformaldehyde fixation and methanol-permeablization, were analyzed for intracellular markers, including mouse IgG1 anti-Oct-4 (Millipore), mouse IgG1 anti-Nestin (Millipore), and polyclonal rabbit PDGFRα (Santa Cruz). Briefly, 5×105 cells were incubated in blocking buffer (10% goat serum in D-PBS) followed by primary antibodies (0.5 μg per sample) or the corresponding isotype controls. A single marker was stained for each sample. After washing, cells were incubated with corresponding secondary antibodies (0.25 μg per sample), including Alexa Fluor® 488 goat anti-mouse IgG1, Alexa Fluor 647 goat anti-mouse IgM, and Alexa Flour 488 goat anti-rabbit IgG (all from Molecular Probes). Before acquisition, live cells were stained with 2 μg/mL propidium iodide (PI) for viability assessment of surface markers. A total of 30,000 events (gate was set for PI or EMA negative populations to exclude dead cells) were acquired using FACS Calibur (BD Biosciences). The viable cell populations were analyzed for the marker expression against corresponding isotype control using FlowJo software. The percentages of positive cells were calculated with the subtraction of isotype background.
Karyotype analysis
Cytogenetic analysis of 20–50 cells was performed using GTG-banding, by the Medical Genetics Cytogenetics Laboratory at Children's Hospital, Oakland, CA.
Statistical analysis
The hESC expansion experiments were repeated 5 times and the data from one typical experiment are presented. OPC differentiations were repeated 5 (comparison of surface combinations MMM, VMM, and VVV) or 6 times (comparison of surface combinations MMM and MVV). Average values of the marker expression from 5 or 6 independent experiments are presented. For statistical analysis, the percentages of OPC markers were analyzed using analysis of variance (ANOVA) followed by paired Tukey-Kramer analysis (JMP® 9.0 software) to determine the statistical significance. A P value<0.05 was considered as significant.
Results and Discussions
Our previous study has shown that VN-PAS supports the expansion of undifferentiated H1 and H7 hESCs, and that H7hESCs can be thawed, expanded, and differentiated to cardiomyocytes on VN-PAS [38,42]. For neural lineage, vitronectin has been shown to promote oligodendrocyte differentiation from hESCs [22], and we have shown previously that hESCs can differentiate to OPCs using a 41-day protocol [41]. Consequently, the hypothesis of this study is that VN-PAS, comprised of an acrylate polymer conjugated to a vitronectin-derived peptide, should be a surface suitable to support OPC differentiation of hESCs. To test this hypothesis, this study is divided into 3 sections to determine if (i) H1 hESCs can be thawed directly onto VN-PAS and expanded at the undifferentiated state; (ii) VN-PAS supports the OPC differentiation of H1 hESCs maintained on GFR-Matrigel; (iii) VN-PAS supports the OPC differentiation of H1 hESCs maintained on VN-PAS.
The production of OPCs from cryopreserved hESCs can be divided into 2 stages: hESC thaw/expansion and OPC differentiation (Fig. 1). The differentiation process is further divided into 3 phases: EB Formation (days 0–27), Plating and Maturation (days 27–34), and Re-plating and Maturation (days 34–41). Adherent cultures during days 27–41 were performed on GFR-Matrigel-coated vessels or VN-PAS vessels. The following surface combinations were evaluated in this study (Fig. 2): hESC thaw/expansion, plating/maturation, and re-plating/maturation on GFR-Matrigel-coated vessels (MMM); hESC thaw/expansion on GFR-Matrigel-coated vessels, plating/maturation and re-plating/maturation on VN-PAS (MVV); hESC thaw/expansion on VN-PAS vessels, plating/maturation and replating/maturation on GFR-Matrigel-coated vessels (VMM); hESC thaw/expansion, plating/maturation, and replating/maturation on VN-PAS vessels (VVV). From these combinations, the effects of the culture surface on hESC expansion stage or on OPC differentiation stage were able to be delineated.
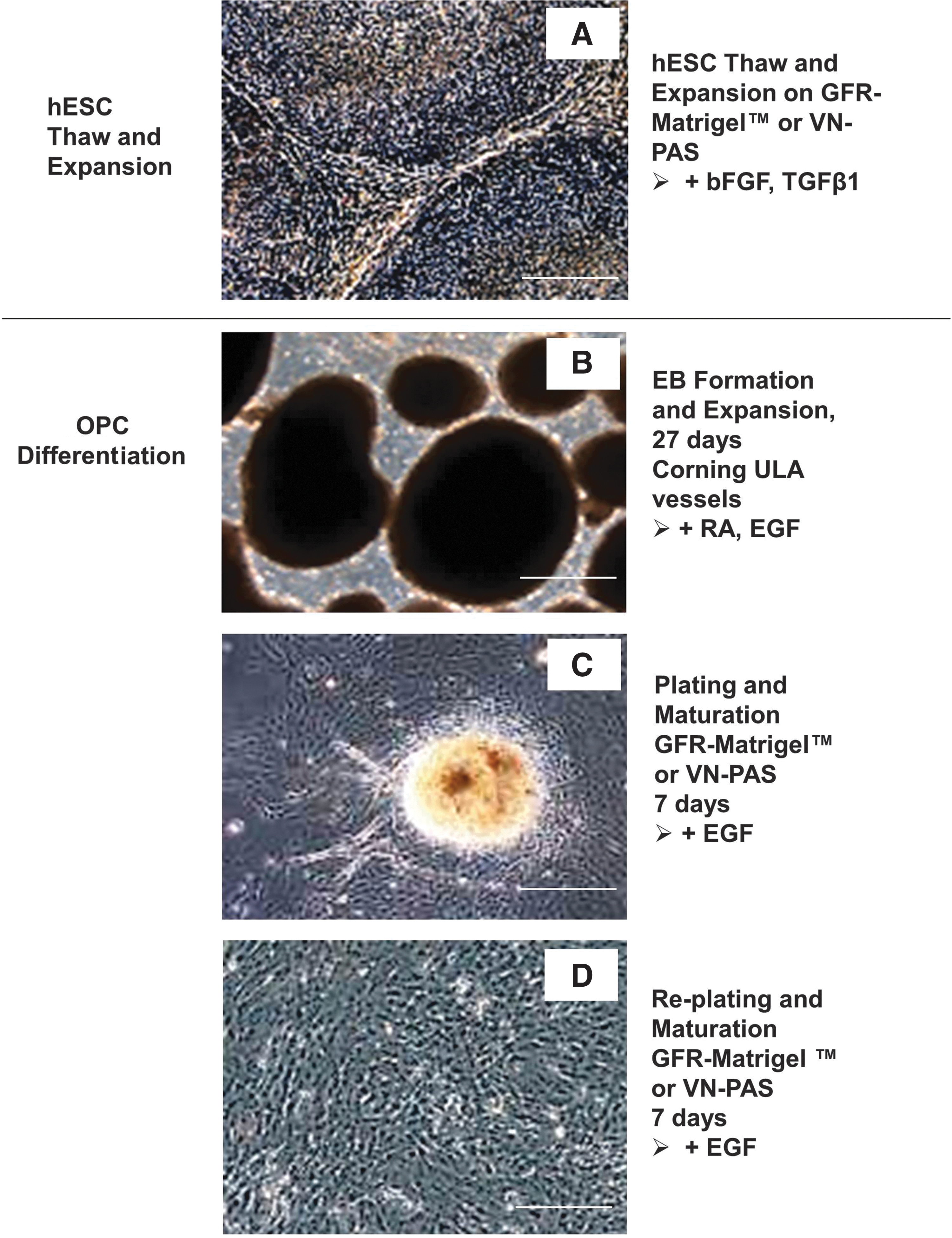
Illustration of oligodendrocyte progenitor cell (OPC) differentiation from human embryonic stem cells (hESCs). Stages to generate hESC-derived OPCs: hESC thaw/expansion and OPC differentiation. The differentiation process is divided into 3 phases: embryoid body (EB) formation (days 0–27) in nonadherent culture, plating and maturation (days 27–34), and re-plating and maturation (days 34–41) in adherent culture.

Schematic description of different surface conditions for OPC differentiation. MMM: all 3 steps of hESC thaw/expansion, plating/maturation, and re-plating/maturation were performed on GFR-Matrigel-coated vessels. MVV: hESC thaw/expansion on GFR-Matrigel-coated vessels, plating/maturation and re-plating/maturation on VN-PAS. VMM: hESC thaw/expansion on VN-PAS vessels, plating/maturation and replating/maturation on GFR-Matrigel-coated vessels. VVV: all 3 steps of hESC thaw/expansion, plating/maturation, and replating/maturation were performed on VN-PAS vessels.
VN-PAS supports hESC thaw and expansion
Cryopreserved H1 hESCs were thawed and resuspended in the culture medium. The cells were divided and seeded onto GFR-Matrigel-coated vessels or VN-PAS vessels to eliminate vial-to-vial variability for the comparison. The cells were serially passaged and maintained in the defined medium for more than 10 passages. The apparent doubling times (include plating efficiency and growth) from 2 VN-PAS cultures were compared with GFR-Matrigel control. All 3 cultures showed consistent and comparable doubling times (most were around 60–80 h) over 13 passages (Fig. 3A) amidst the standard passage to passage variability evident in each of the cultures. This observed variability was within the range of our routine hESC cultures. The slightly slower doubling time for VN-PAS may reflect the cell response to the new culture condition upon thaw. This apparent difference disappeared around passage 12–13 for the cell bank used in this study (Fig. 3A). Expression of Oct-4 and Tra1-60, markers for hESC pluripotency, was compared at selected passages by flow cytometry. Similar level of expression (70%–86% after passage 6) was observed for all 3 cultures [Fig. 3B, C; see Supplementary Fig. S1 (Supplementary Data are available online at
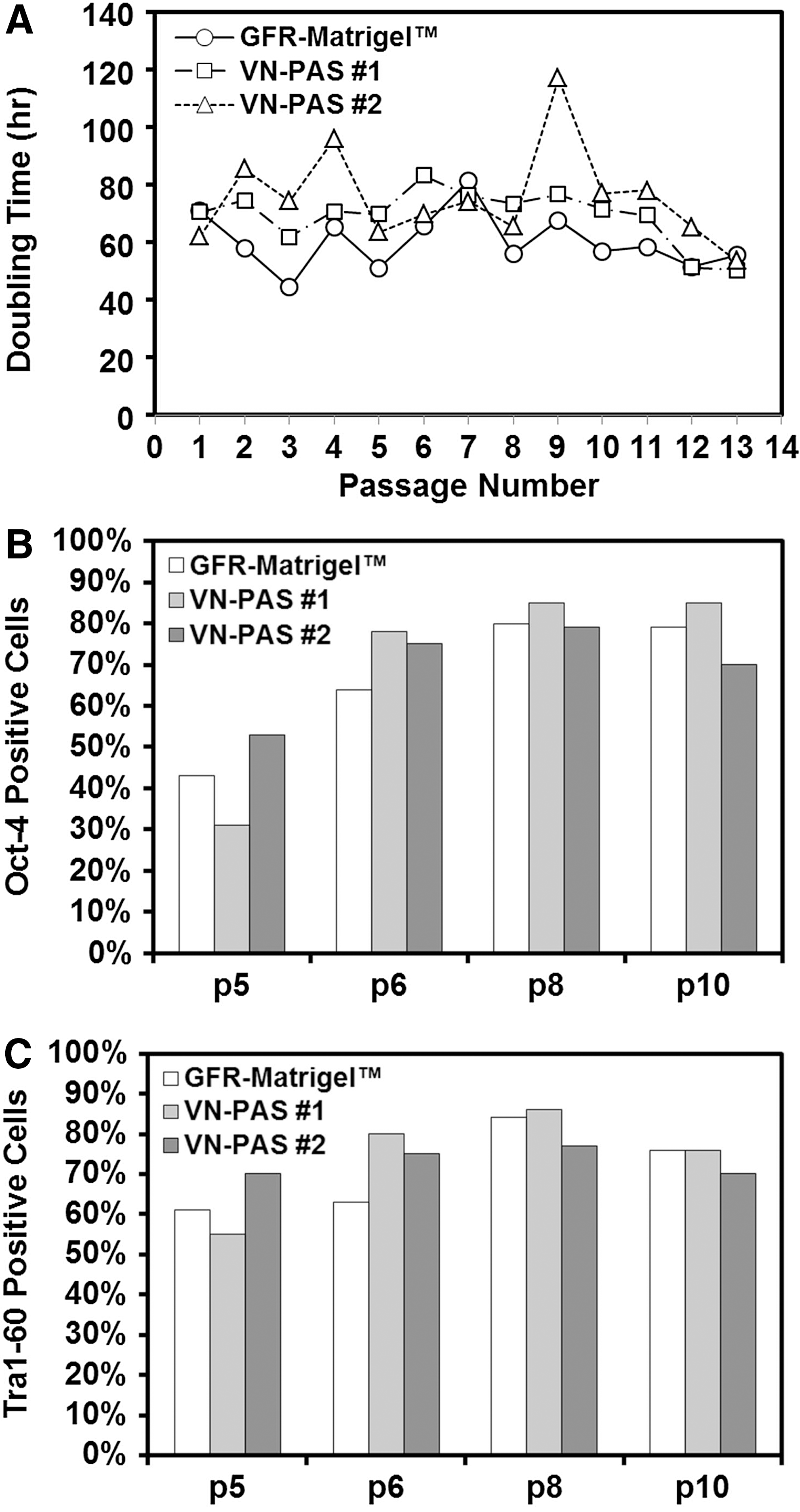
Long-term expansion of H1 hESCs on VN-PAS T-75 flasks. H1 hESCs were thawed and seeded onto GFR-Matrigel-coated T-75 flasks or VN-PAS T-75 flasks.
In our experience, hESCs are more sensitive to the changes in the culture condition upon thaw than the changes in the established cultures. The ability of VN-PAS to support the colony enrichment of H1 hESCs directly from thaw indicates that VN-PAS is a robust surface for hESC culture.
VN-PAS supports OPC differentiation
The OPC differentiation on VN-PAS was evaluated for H1 hESCs expanded on GFR-Matrigel-coated surface. After EB formation, cells were plated (day 27) and replated (day 34) onto either GFR-Matrigel-coated T-75 flasks (surface combination MMM) or VN-PAS T-75 flasks (surface combination MVV). The morphology of OPCs at day 41 on both surfaces was similar, with bipolar appearance characteristics of immature OPCs (Fig. 4A, B). Expression of OPC markers NG2, PDGFRα, and Nestin was also similar for OPCs differentiated on GFR-Matrigel (MMM) or VN-PAS (MVV; Fig. 4C). Flow cytometry histograms typical for these OPCs are shown in Fig. 4D. The expression pattern of these markers by immunocytochemistry for OPCs differentiated using this protocol was shown in an early report [41]. No Oct-4 and Tra1-60 expression was observed for the 2 groups (data not shown). Few cells were positive for β-tubulin III (neuron lineage) or glial fibrillary acidic protein (GFAP, astrocyte lineage) (Supplementary Table S1). Although the expression of NG2, a chondroitin sulfate proteoglycan expressed by OPCs, was more variable than Nestin and PDGFRα, similar variability was observed for both surface conditions. These results suggest that VN-PAS supports OPC differentiation from hESCs similarly to GFR-Matrigel.
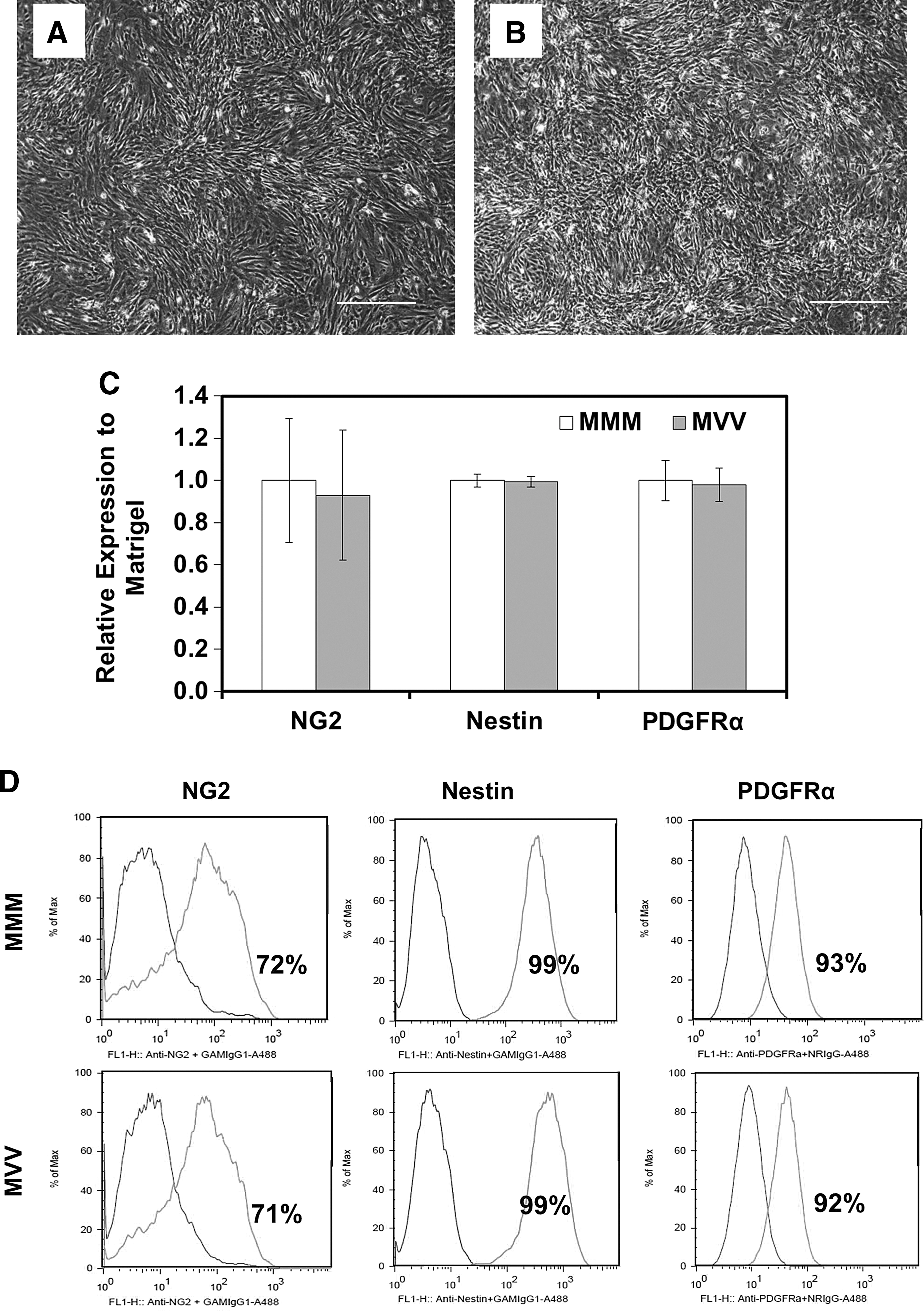
OPC differentiation on VN-PAS surface. Phase-contrast micrograph (40×) of day 41 OPCs on
ECM plays an important role for OPC differentiation from hPSCs in various EGF-dependent and platelet-derived growth factor (PDGF)-dependent OPC differentiation protocols [43]. Different ECM proteins, including fibronectin, laminin, and Matrigel, impact OPC proliferation, maturation, migration, and survival [15]. The major component of GFR-Matrigel is laminin, which binds α6β1 integrin expressed on oligodendrocyte precursors to regulate OPC proliferation and maturation [21]. Similarly, vitronectin receptors αvβ1, αvβ3, and αvβ5 are differentially expressed at different OPC developmental stages and play an important role in modulating OPC migration (early stage), proliferation (middle stage), and the final differentiation (late stage) [44]. Consistently, vitronectin has been shown to promote OPC differentiation from hESCs [22]. The VN-PAS reported in this study contains peptide Ac-KGGPQVTRGDVFTMP, which is derived from the active cell attachment domain of vitronectin interacting with αvβ5 [38]. From the results in this study, this peptide sequence is sufficient to support OPC differentiation from hESCs.
An integrated hESC expansion and OPC differentiation culture on VN-PAS
Subsequent to the establishment of VN-PAS as a viable surface supporting not only the thaw/expansion of H1 hESCs but also the OPC differentiation of H1 hESCs maintained on GFR-Matrigel, this part of the study evaluated whether H1 hESCs thawed and maintained on VN-PAS were able to differentiate to OPCs. One vial of H1 hESCs was thawed and the cells were divided and plated on GFR-Matrigel-coated surface or VN-PAS. H1 hESCs expanded on VN-PAS were dissociated to make EBs for the suspension stage of OPC differentiation and then plated (day 27) and replated (day 34) onto VN-PAS (surface combination VVV). EBs were also plated and replated onto GFR-Matrigel-coated vessels (surface combination VMM) to decouple the effects of vitronectin peptide on the expansion or differentiation stages. Surface combination MMM was used as the control.
At the end of differentiation, the expression of Nestin and PDGFα was found to be comparable for all 3 conditions (Fig. 5A). However, higher levels (1.5–1.6-fold) of NG2 were observed for the surface combinations VMM and VVV compared to MMM (P<0.05), but not between VMM and VVV. Flow cytometry histograms from a typical experiment are shown in Fig. 5B. This observation indicates that the increased NG2 expression is a result of H1 hESC thaw and expansion on VN-PAS but not a result of the differentiation on VN-PAS. The mechanism of this phenomenon is not clear, and further studies are required to compare hESCs grown on GFR-Matrigel or VN-PAS at cellular and molecular levels. For OPC proliferation, the doubling ratio during days 34–41 of the differentiation procedure was compared. Similar doubling ratio (1.9–2.4) was observed for all the surface combinations (Fig. 6). The OPCs differentiated using this protocol and defined by the 3 markers, NG2, Nestin, and PDGFRα, were able to mature into cells with oligodendrocyte morphology (Supplementary Fig. S2). Some variations were observed in the EB size distribution and plating efficiency on day 27 (data not shown). Further investigations such as dynamic suspension culture in bioreactors will be important to provide more controlled and uniform culture conditions.

An integrated hESC thaw/expansion and OPC differentiation on VN-PAS surface.
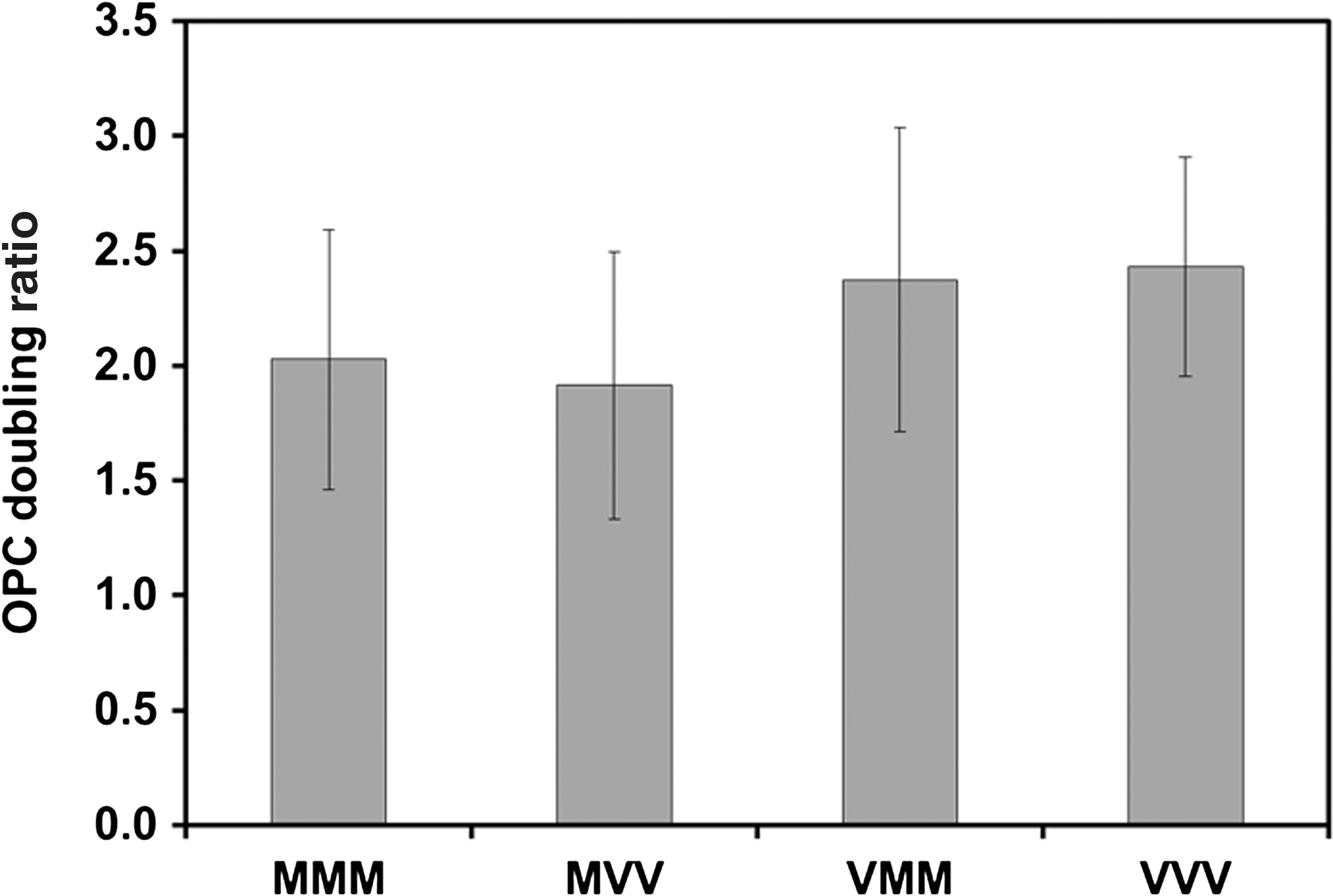
OPC doubling ratio during days 34–41 for different surface combinations. The OPC doubling ratio was calculated as the cell number at day 41 divided by the cell number at day 34 (n=5). The average values from 5 independent experiments are shown.
Defined, low-cost, robust, and scalable culture methods will need to be developed for the widespread deployment and commercialization of hESC-derived cell therapies. Several synthetic surfaces that support hPSC growth have been developed in recent years that could eliminate animal derived or recombinant ECMs, and are potentially scalable for commercial manufacture [34 –38,45]. Synthetic polymer surfaces that do not contain specific biological ligands have so far relied on the medium components to enhance cell adhesion [35,37,46]. For example, conditioned media or serum-free media with abundant bovine serum albumin have to be used; even so, only some hPSC lines were supported under these conditions [35,37,46]. Thus, synthetic peptide surfaces with biological ligands have been more robust than simple polymer surfaces to culture hPSCs in the defined serum-free medium [36,38]. From the results in this study, hESC expansion on VN-PAS resulted in OPCs with increased NG2 expression upon differentiation, highlighting the importance of hESC culture conditions on the cell characteristics during specific lineage differentiation. However, only a few studies have tested the synthetic surface for lineage-specific differentiation. While VN-PAS has been shown to support hESC differentiation into cardiomyocyte and a polypeptide surface was developed for early mesoderm differentiation [38,47], to our knowledge, this study is the first report to derive OPCs from hPSCs on synthetic peptide surfaces. Although further study is required for more detailed characterizations of OPCs derived on synthetic peptide surfaces, the integrated hPSC thaw/expansion and differentiation on VN-PAS simplifies the process of OPC production from hPSCs and moves one-step further toward the eventual commercial manufacture of hPSC-derived cell therapeutics [48].
Footnotes
Acknowledgments
We thank Mr. Melaku Teklemichael for his help on cell culture and Ms. Stacey Lawson for her help on reagent preparation. Funding support in part from Florida State University (FSU) FYAP award to YL is also acknowledged.
Author Disclosure Statement
No conflict of interests is declared. No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.