
Abstract
Recent studies demonstrated that the endogenous expression level of Sox2, Oct-4, and c-Myc is correlated with the pluripotency and successful induction of induced pluripotent stem cells. Periodontal ligament cells (PDLCs) have a multilineage differentiation capability and ability to maintain the undifferentiated stage, which makes PDLCs a suitable cell source for tissue repair and regeneration. To elucidate the effect of an in vitro culture condition on the stemness potential of PDLCs, we explored the cell growth, proliferation, cell cycle, and the expression of Sox2, Oct-4, and c-Myc in PDLCs from the passage 1 to 7 with or without the addition of recombinant human bone morphogenetic protein-4 (rhBMP4). Our results revealed that BMP-4 promoted cell growth and proliferation, arrested PDLCs in the S phase of cell cycle, and upregulated the propidium iodinate value. It was revealed that without the addition of rhBMP4, the expression of Sox2, Oct-4, and c-Myc in PDLCs only maintained the nucleus location until passage 3, and then lost the nucleus location subsequently. The mRNA expression in PDLCs further confirmed that the level of Sox2 and Oct-4 peaked at passage 3 and then decreased afterward, whereas c-Myc maintained consistently the upregulation along the passages. After the treatment with rhBMP4, the expression of Sox2, Oct-4, and c-Myc in PDLCs maintained the nucleus location even at passage 7, and the mRNA expression of Sox2 and Oct-4 significantly upregulated at the passages 5 and 7. These results demonstrated that addition of rhBMP-4 in the culture medium could improve the current culture condition for PDLCs to maintain in an undifferentiated stage.
Introduction
The endogenous expression level of Sox2, Oct-4, and c-Myc has been shown to be correlated with the successful induction of iPSCs and the pluripotency [4 –6]. MSCs from dental tissues have shown potential to be reprogrammed by introducing Oct-4/Sox2/Klf4/c-Myc or Oct-4/Sox2/Nanog/Lin28 [4 –7]. It is also known that cellular stresses (including heat shock, oxidative stress, and hypoxia) are able to activate the signal that leads somatic cells to reprogramming and regeneration [8,9]. In our previous study, it has been demonstrated that the ischemic culture stimulated the expression of Oct-4 and ABCG2 in human dental pulp cells [10]. Therefore, it is possible that the pluripotency network can be re-established or activated in somatic cells [11].
Bone morphogenetic protein-4 (BMP-4), a member of the transforming growth factor-beta (TGFβ) superfamily, can control vasculogenesis and induce embryonic and extraembryonic mesoderm formation in embryo [12]. BMP-4 stimulates the formation and outgrowth of pericytes in blood vessel formation and maintenance, which may be the origin of stem cell niche [13]. Further, BMP-4 increases the proportion of Oct-4-positive cells in parthenogenic embryos during maturation, indicating that BMP-4 may provide a suitable microenvironment to maintain cells in an undifferentiated stage [14]. However, as of yet, the specific role of BMP-4 in maintaining stemness of PDLCs has not been elucidated yet. In the present study, we investigated cell growth, proliferation, cell cycle, and altered expression of reprogramming markers (Sox2, Oct-4, and c-Myc) in PDLCs at various passages with or without the addition of recombinant human BMP4 (rhBMP4) in the culture medium.
Materials and Methods
Isolation and expansion of human PDLCs
The protocols used (EC00171) were approved by the Human Ethics Committee from the Queensland University of Technology (QUT). Normal human premolars were extracted from healthy young adults (12–30 years) undergoing orthodontic treatment, after obtaining informed consent from each patient. PDLCs from 3 individual patients were used in each experiment, and each experiment was repeated 3 times. The PDLCs from each patient were not pooled; instead, they were cultured and studied separately according to the previous publications [15]. No significant variation was found among various patients. PDLCs were obtained from periodontal ligament tissues by an explant culture as previously described [15]. PDLCs were cultured in the Dulbecco's modified Eagle medium with low glucose (Invitrogen) supplemented with 10% fetal bovine serum (HyClone), 10 U/mL penicillin G, and 10 mg/mL streptomycin (Invitrogen). PDLCs were incubated at 37°C in 5% CO2 and characterized to the multilineage differentiation capability, the expression of cell surface markers STRO-1, CD146, CD44, etc., and the differentially expressed proteins and genes according to our previous studies [2,3]. rhBMP4 (R&D system) at the optimum concentration of 10 ng/mL indicated by previous studies was added to the culture medium throughout the suspension period [16]. Cells cultured in a normal medium served as controls. The medium was changed every 3 days. The images of the cell morphology were captured under an inverted light microscope.
Beta-galactosidase staining for cell senescence
Cytochemical staining for the senescence-associated β-galactosidase assay was performed by seeding PDLCs from passages 1, 3, 5, and 7 at the cell density of 1×103 cells/well in a 24-well plate. The cells were allowed to attach overnight and then were washed with PBS, fixed, and incubated overnight at 37°C with an X-gal chromogenic substrate at pH 6.0 according to the protocol provided by the β-galactosidase staining kit (Cell Signaling Technology). The images of cell morphology were captured under an inverted light microscope.
Cell counting kit 8 assay for cell proliferation
The cell growth was measured by means of cell counting. Cell proliferation was investigated by the cell counting kit 8 (CCK8) assay. The cultures of PDLCs were serum-deprived for 24 h before the induction. A total of 104 cells per well were plated in 96-well plates and cultured for 24 h in normal conditions with/without rhBMP4 induction. Cell proliferation of PDLCs from passage 1 to 7 was evaluated using the CCK8 (Dojindo) according to manufacturer's instructions. Briefly, 10 μL of the CCK8 solution was added to the culture medium, and incubated for additional 3 h. The absorbance was determined at 450-nm wavelength.
Flow cytometry for cell cycle analysis
The cultures of PDLCs were serum-deprived for 24 h before the induction. About 1×105 PDLCs from passage 1 to 8 with/without rhBMP4 treatment were harvested by trypsinization, washed twice in cold PBS, and fixed in 70% alcohol for 30 min on ice. After washing in cold PBS 3 times, cells were incubated with 0.5% propidium iodinate (PI) for 30 min at 4°C. The cells were analyzed using a FACSCalibur flow cytometer (BD Biosciences). Each condition was repeated in triplicate. Data were analyzed using FCSExpress software.
Immunofluorescent staining and nucleus expression ratio of reprogramming markers in PDLCs at various passages
PDLCs from the passages 1 to 7 were cultured in chamber slides (Nunc) until 80% confluence, and fixed with 3% paraformaldehyde for 15 min. The slides were rinsed in PBS 3 times for 5 min, respectively, then permeabilized with 0.1% Triton for 20 min, and incubated with 10% swine serum for 1 h at room temperature. Slides were transferred to a humidified chamber and stained with Sox2 (1:50; R&D system), Oct-4 (1:100; Chemicon), and c-Myc (1:50; Santa Cruz) antibodies overnight at 4°C. Samples were washed 3 times in PBS and incubated with a fluorochrome-labeled secondary antibody (1:150; Invitrogen) for 3 h, and then mounted for analysis. The images were captured under a Zeiss fluorescent microscope. The number of positive cells expressing Sox2, Oct-4, and c-Myc was measured in 3 randomly chosen areas from 3 different patients. The expression ratios of reprogramming markers in the nucleus were determined by the positive cell numbers dividing with the total cell numbers.
Quantitative real-time reverse transcription–polymerase chain reaction
Total RNA was isolated from PDLCs at passages 1 to 7 using Trizol reagent (Invitrogen) following the manufacturer's protocol. The concentration and quality of RNA samples were measured with spectrophotometers and gel electrophoresis. The first-strand cDNA was synthesized from 1 μg of total RNA using SuperScript III (Invitrogen) in a total volume of 20 μL. About 2.5 μL of the reaction mixture was incubated with 2×SYBR Green I Master Mix (Applied Biosystems) in a total volume of 25 μL. Primers used for detection are listed in Table 1. The conditions for polymerase chain reaction (PCR) were as follows: 95°C for 10 min for activation, followed by 40 cycles of denaturation at 95°C for 15 s each, and finally, primer extension at 60°C for 1 min. Sox2, Oct-4, c-Myc, BMP-4, and 18s mRNA were quantified using an ABI Prism 7000 sequence detection system (Applied Biosystems). Each plate contained 18s as the housekeeping gene to normalize the PCR data. All experiments were repeated 3 times from 3 separate samples. Raw data were acquired and processed to calculate the threshold cycle (Ct) value and relative gene expression values. The delta-delta Ct method was performed to analyze the result.
Statistical analysis
All experiments were repeated at least 3 times. The SPSS16.0 software package (SPSS, Inc.) was used for the statistical tests. One-way analysis of variance was applied to compare the differences among passages. If equality of variances could be assumed, which was also concluded by a hypothesis test, Bonferroni test was performed. The difference was considered as being of statistically significant at P<0.05. The rhBMP4-treated cells were compared with the control cells at each passages using the Student's t-test. *P<0.05 was considered to be significant.
Results
Morphological alternations and cell senescence of PDLCs at various passages
PDLCs at passage 1 displayed well-spread attachment to the flask and showed a fibroblast-like morphology and single monolayer growing pattern (Fig. 1A). PDLCs at passage 3 (Fig. 1B), passage 5 (Fig. 1C), and passage 7 (Fig. 1D) showed signs of cell senescence, including alternations in the cell morphology with enlarged size and an increased nuclear/cytoplasm ratio. No difference in the morphology was observed using the microscopic observation between the normal and rhBMP4-treated groups at early passages (passage 1 to 3, data not shown). However, PDLCs with rhBMP4 treatment at passage 7 (Fig. 1E) showed a smaller cell size and decreased nuclear/cytoplasm ratio and cell secretion compared with the untreated groups (Fig. 1D).

Morphological characteristics of periodontal ligament cells (PDLCs) at various passages. PDLCs at passage 1 showed a fibroblast-like morphology and a single-monolayer growing pattern well spread attaching to the flask
To further authenticate the replicative senescence state of PDLCs at various passages, their senescence-associated β-galactosidase activity (SA-β-gal) was assessed. The β-galactosidase is an enzyme expressed by senescent cells. This assay revealed that PDLCs at passage 7 were strongly positive for SA-β-gal activity, which stained an intense blue color (Fig. 1I), and PDLCs from passage 5 (Fig. 1H) showed blue staining, but not as intense as the cells from passage 7. However, PDLCs from passage 1 (Fig. 1F), passage 3 (Fig. 1G), and the rhBMP4-treated group (passage 7, Fig. 1J) did not show any obvious blue staining, indicating that PDLCs from early passage were presenescent, and BMP-4 may maintain PDLCs at a presenescent state.
Cell growth, proliferation, and cell cycle of PDLCs at various passages
To investigate the biological effect of rhBMP, the cell number, proliferation, and cell cycle of PDLCs at various passages were investigated after treatment. The cell number of PDLCs with rhBMP4 treatment was significantly increased at various passages compared with the control groups (Fig. 2I, *P<0.05). As shown in Fig. 2J, treatment with rhBMP significantly stimulated the cell proliferation of PDLCs at passages 5 and 7 (Fig. 2J, *P<0.05), although no significant difference at passages 1 and 3 (Fig. 2J, P>0.05). These data indicate that cell growth and proliferation can be significantly stimulated by BMP-4.
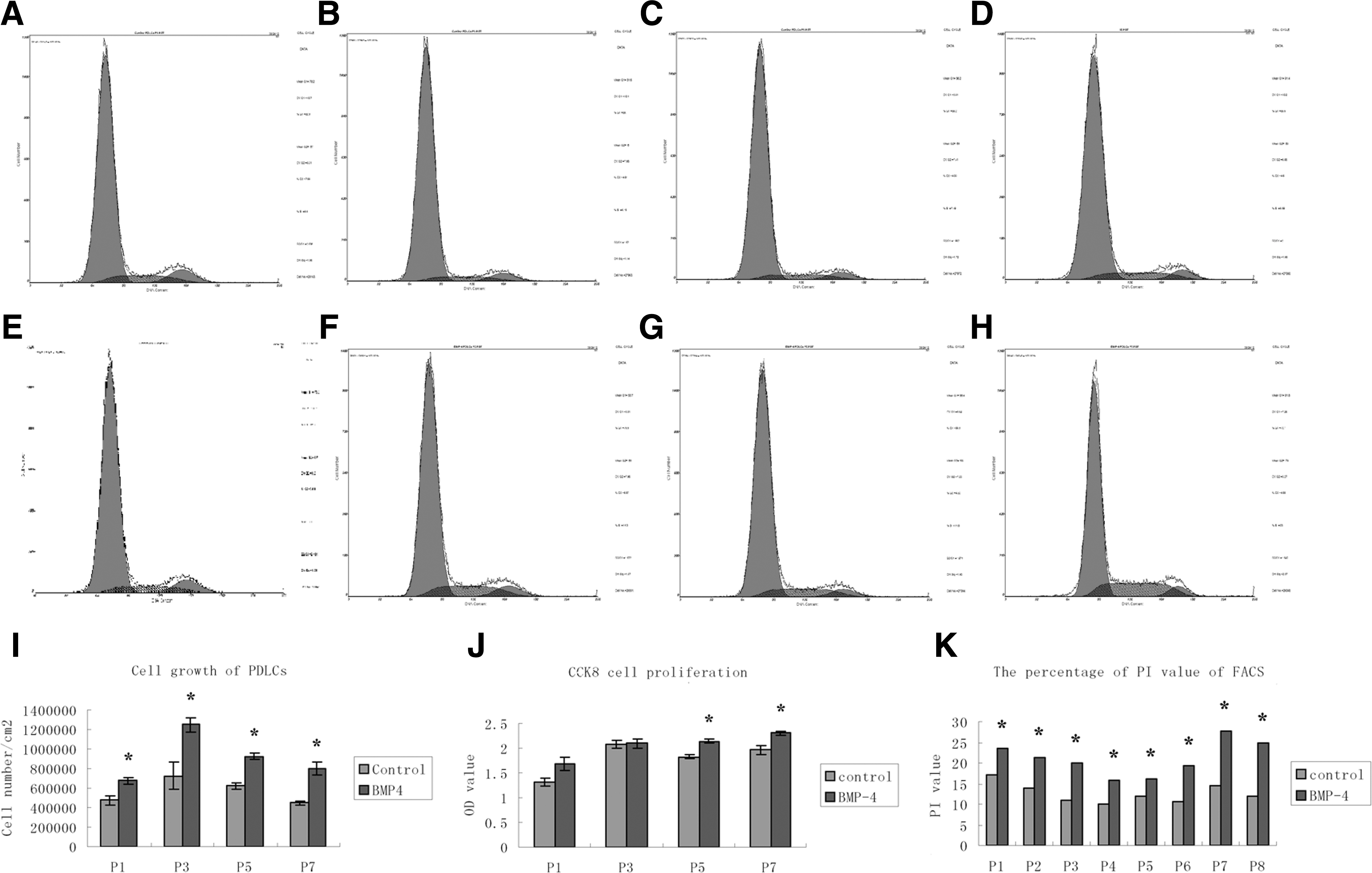
Cell growth, proliferation, and cell cycle of PDLCs at various passages. The cell number of PDLCs with rhBMP4 treatment was significantly upregulated at various passages compared with the control groups
Since the effect of BMP-4 on the stimulation of cell growth and proliferation may result from cell cycle effects, the cell cycle distribution of the rhBMP4-treated and control group PDLCs at various passages was investigated by flow cytometry. As shown in Fig. 2A–H, K and Table 2, rhBMP4 arrested PDLCs from various passages in the S phase of the cell cycle, as demonstrated by a significant increase in the percentage of cells in the S phase (passages 1 to 8, Fig. 2A–H, K & Table 2, *P<0.05, **P<0.001) and G2/M (passages 1, 2, and 6, Fig. 2A–H, K & Table 2, *P<0.05) with a corresponding decline in the G0/G1 phase (passage 1 to 8, Fig. 2A–H, K & Table 2, *P<0.05). Notably, the percentage of PI=(S+G2/M)% in rhBMP4-treated PDLCs from passages 1 to 8 were significantly higher than the control group (Figs. 2A–H and 3C & Table 2, *P<0.05, **P<0.001). These results imply that BMP-4 may promote cell growth and proliferation of PDLCs, especially from late passages, through arresting cells in the S and G2/M phase of cell cycle and enhancing the PI value.
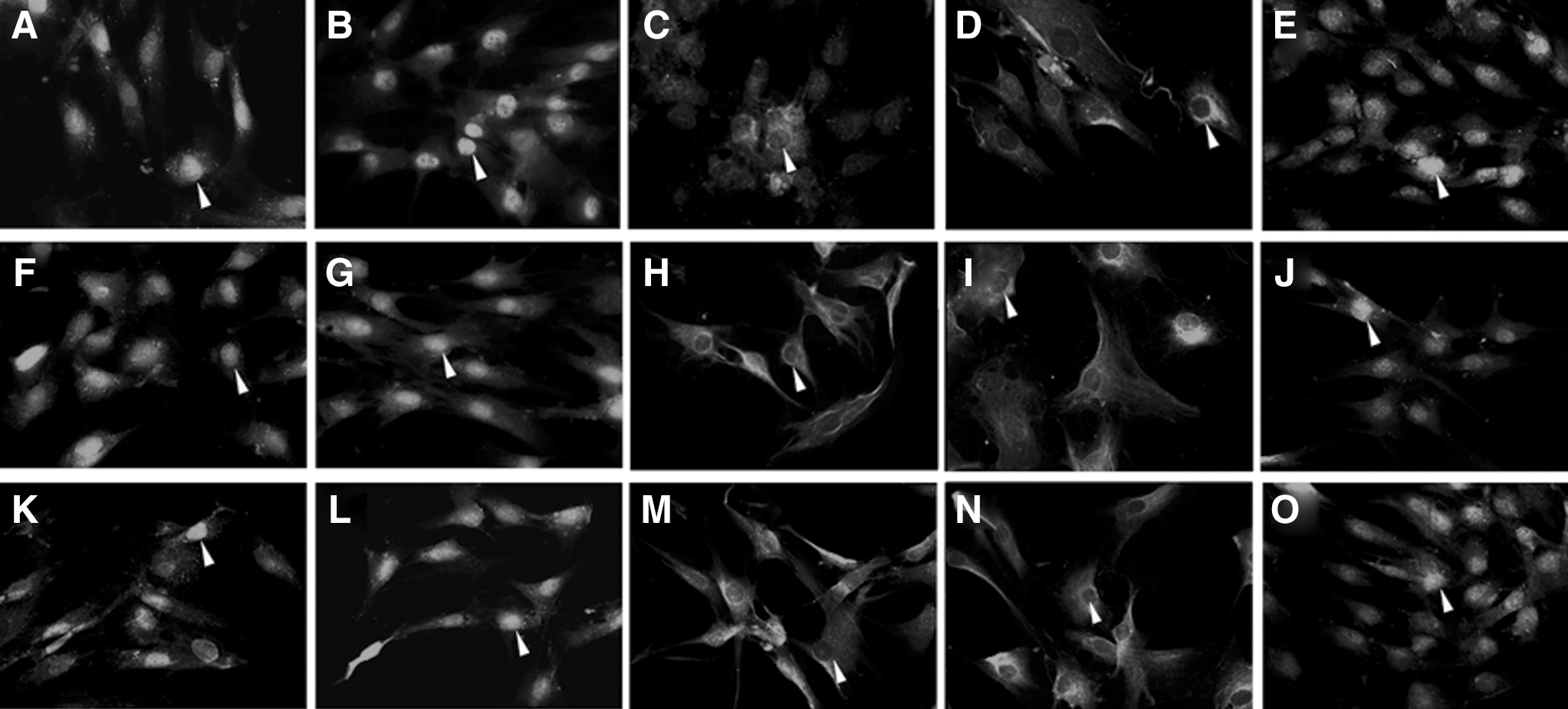
Immunofluorescent staining of Sox2, Oct-4, and c-Myc in PDLCs at various passages. Tense staining of Sox2
A significant increase in the percentage of periodontal ligament cells in the S phase (passages 1 to 8, a P<0.05, b P<0.001) and G2/M (passages 1, 2, and 6, a P<0.05), with a corresponding decline in the G0/G1 phase (passages 1 to 8, a P<0.05) was presented.
BMP4, bone morphogenetic protein-4; PI, propidium iodinate.
Subcellular localization and nucleus expression ratio of Sox2, Oct-4, and c-Myc in PDLCs at various passages
Immunofluorescent staining of Sox2, Oct-4, and c-Myc was investigated in PDLCs from passages 1 to 7; representative figures at passage 1, 3, 5, and 7 are provided (Fig. 3). The expression of Sox2 (Fig. 3A–D), Oct-4 (Fig. 3F–I), and c-Myc (Fig. 3K–N) was obviously detected in the nucleus and slightly expressed in the cytoplasm of PDLCs at passages 1 and 3. Along with passages, Sox2, Oct-4, and c-Myc all lost the nucleus location and maintained weak cytoplasm staining at passages 5 and 7. The nucleus expression ratio of Sox2 (Fig. 4A), Oct-4 (Fig. 4B), and c-Myc (Fig. 4D) showed a similar expression pattern and altered in a passage-dependent manner, displayed no significant difference between passages 1 and 3 (P>0.05), but significantly decreased at passages 5 and 7 (*P<0.05).
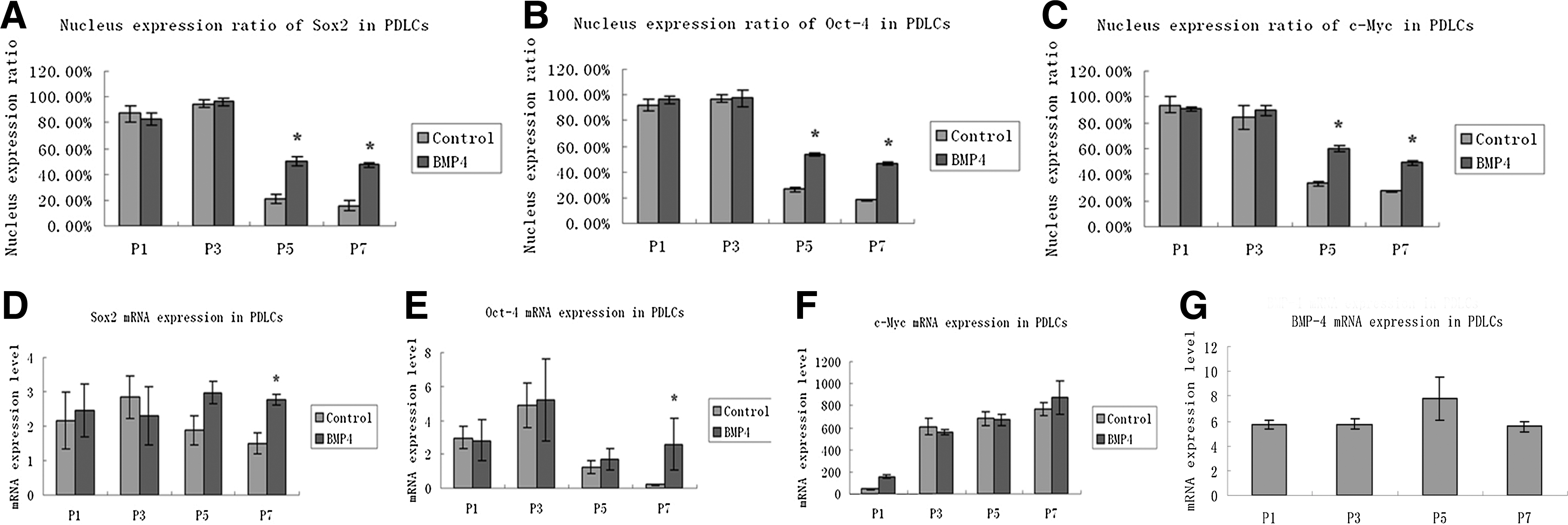
Nucleus expression ratio and mRNA expression of Sox2, Oct-4, c-Myc, and BMP-4 in PDLCs at various passages. The nucleus expression ratio of Sox2
After treated with rhBMP4, Sox2 (Fig. 3E), Oct-4 (Fig. 3J), and c-Myc (Fig. 3O) maintained the nucleus location even at passage 7, whereas the control groups at passage 7 (Fig. 3D, I, N) all lost the nucleus location without rhBMP4 induction. The nucleus expression ratio showed that there was no significant difference between the rhBMP4-induced and control group at passages 1 and 3 (Fig. 4A–C, P>0.05), although a significantly higher expression ratio of all 3 markers at passages 5 and 7 compared with the control groups (*P<0.05).
mRNA expression of Sox2, Oct-4, c-Myc, and BMP-4 in PDLCs at various passages
PDLCs from passage 1 to 7 were examined for Sox2, Oct-4, and c-Myc mRNA expression by real-time PCR, and the results of typical passages that showed the trend were presented (Fig. 4D–F). mRNA expression of Sox2 (Fig. 4D) and Oct-4 (Fig. 4E) increased and peaked at passage 3, and then decreased afterward. mRNA expression of c-Myc (Fig. 4F), however, showed an absolutely different pattern and was over 100 times higher expressed than Sox2 and Oct-4; cMyc also maintained consistently the upregulated levels at passage 7. Differences of Sox2, Oct-4, and c-Myc among the passages were statistically significant (P<0.05). Moreover, the mRNA expression level of Sox2 (Fig. 4D) and Oct-4 (Fig. 4E) showed significant upregulation in PDLCs with rhBMP4 treatment at passage 7(*P<0.05). Although, the mRNA expression level of c-Myc showed no significant difference compared with the control group (P>0.05, Fig. 4F), implying the similar expression pattern and potential cooperative interaction between Sox2 and Oct-4. We further investigated whether the basal endogenous BMP-4 level had any impact on the expression of Sox2, Oct-4, and cMyc in PDLCs. It was found that the basal endogenous BMP-4 mRNA level was relatively low in PDLCs, and no significant alternations were found in the mRNA expression of BMP-4 from passages 1 to 7 (P>0.05, Fig. 4G), indicating that there was no direct correlation between the endogenous BMP-4 level and the expression of Sox2, Oct-4, and c-Myc in PDLCs.
Discussion
Extended in vitro culture in the current methodology is known to cause morphologic alternations, loss of expression of pluripotent markers, and widespread senescence [17,18]. The PDLCs used in this study showed significant cell senescence and morphological changes at the late passages in the normal cell growth medium. It is known that the loss of regeneration capacity of MSCs can be rejuvenated by an altered extracellular microenvironment [18], and the microenvironment produces systemic factors to modulate the molecular pathways that control the regenerative properties and support tissue regeneration [19,20]. Since Oct4, Sox2, and c-Myc cooperatively maintain or induce the regulatory network responsible for self-renewal and pluripotency, it has been assumed that these genes have important and conserved functions in stem cells. However, the specific functions of these markers in various cells remain poorly defined. Sox2 belongs to the SRY-related HMG box transcription factor family and plays an essential role in regulation of pluripotency of ESCs during early embryonic development [21,22]. Oct-4 is an essential regulator of pluripotency in ESCs and iPSCs [23]. The Oct-4 and Sox2 levels in human ESCs are characteristic of the undifferentiated state, and loss of expression results in differentiation and progressive loss of pluripotency [11,24]. Sox2 and Oct-4, the master transcription factors for maintenance of the undifferentiated state and self-renewal capability, can interact cooperatively to activate the expression of pluripotency factors such as c-Myc, FGF4, and Nanog and determine the cell fate [21,25]. V-myc myelocytomatosis viral oncogene homolog (c-Myc), a frequent contributory oncogene in tumors, can mediate reprogramming through altering the telomere of a developmentally restricted cell [26].
Sox2, Oct-4, and c-Myc are transcriptional factors described as a nuclear protein. Although the molecular basis of these transcriptional factors to control cell fate is not clear, yet it has been demonstrated that nuclear import is a requirement for their function [4,6]. In these cases, when the cytoplasmic location is detected, the nuclear envelope may be disassembled, allowing the protein exchange between the nucleus and cytoplasmic compartments. Although the functional relevance of maintaining Sox2, Oct-4, and c-Myc in the nucleus is not known, the activity of Sox2, Oct-4, and c-Myc in PDLCs is associated with the multilineage differentiation potential and may be controlled by nucleus–cytoplasmic shuttling [27,28].
Adding growth factors to the culture solution and improving the culture condition are simple and effective ways of delivering the growth factor signals to the cells. As known from in vivo studies, bone morphogenetic factors, members of the TGF-β superfamily, have a notable value in development and differentiation [16]. Bone morphogenetic factors, presented in a wide variety of stem cell niches to maintain cell functions, can replace Klf4 to initiate reprogramming and boost the efficiency of the Oct-4/Sox2 complex and single-factor Oct-4-mediated reprogramming of mouse embryonic fibroblasts [29].
BMP-4 is reported to maintain cell proliferation and pluripotent state through Smad, PI3K/Akt, and Wnt1/beta-catenin in mouse ESCs [30 –32]. Interestingly, the microarray profiles of Oct-4 knockdown and BMP-4-treated H1 cells were compared. There are 1,395 Oct-4 responsive genes and 1,466 BMP-4 responsive genes identified, which showed a significant overlap [23]. Moreover, it is reported that mouse skin keratinocytes treated with BMP-4 can be reprogrammed to a more developmentally potent stage, and redifferentiate into neuronal-lineage cells through upregulating Oct-4, Sox-2, Nanog, Utf1, and Rex-1 [33]. Stromal fibroblasts from head/neck cancer tissues produce growth factors, including BMP-4, which affects the functional properties and remains undifferentiated stage of keratinocytes with in vitro culture, including promotion of epithelial–mesenchymal transition-like alterations. Oct-4 is reported to centralize on the nucleus of the human tongue OSCC cell line Tca8113 after induced by BMP-4, with upregulation of ABCG2, hTERT, CD44, and Bmi-1 [34]. This is consistent with our result that Sox2, Oct-4, and c-Myc maintained the nucleus location and relatively high mRNA expression even at late passages in PDLCs with rhBMP4 induction, revealing that BMP-4 may provide a suitable microenvironment to maintain cells in an undifferentiated stage [30,31]. This suggests that the effects seen with BMP-4 treatment may be due to transcriptional regulation through the Sox2, Oct-4, and c-Myc signaling pathway. Moreover, the present study revealed that BMP-4 promoted cell growth and proliferation, arrested PDLC in the S and G2/M phase of cell cycle, and enhanced the PI value. The present study indicates that although BMP-4 alone may not be sufficient to lead to direct reprogramming [4,7], it may play an essential role in modification of the microenvironment and culture system that maintain cell growth and the expression of multipotency markers. Therefore, BMP-4 may be an effective way to optimize the culture environment and improve stem cell properties of PDLCs during a long-term culture.
In summary, this study revealed that BMP-4 contributes to promote cell growth, proliferation through cell cycle modulation, and maintains the expression of reprogramming markers even at late passages. This is of great importance since it suggested that the microenvironment can manipulate key molecular pathways controlling stemness of PDLCs. However, future studies are required to investigate the molecular mechanisms of BMP4 in maintaining the pluripotency with a long-term in-vitro culture.
Footnotes
Acknowledgments
This study was supported by the National Natural Science Foundation of China (no. 81070830), the Specialized Research Fund for the Doctoral Program of Higher Education (no. 20120171120070), the Fundamental Research Funds for the Central Universities (no. 11ykpy51), the Bureau of Chinese Medicine of Guang Dong Province (no. 20121147), Natural Science foundation of Guangdong Province (no. 10451008901005923) and the Department of Health of Guang Dong Province (no. B2012141).
Author Disclosure Statement
No competing financial interests exist.