
Abstract
Human umbilical cord-derived perivascular cells (PVCs) are a recently characterized source of mesenchymal stromal cells that has gained much interest in the field of cellular therapeutics. However, very little is known about the changes in fate potential and restrictions that these cells undergo during gestational development. This study is the first to examine the phenotypic, molecular, and functional properties of first trimester (FTM)-derived PVCs, outlining properties that are unique to this population when compared to term (TERM) counterparts. FTM- and TERM-PVCs displayed analogous mesenchymal, perivascular, and immunological immunophenotypes. Both PVCs could be maintained in culture without alteration to these phenotypes or mesenchymal lineage differentiation potential. Some unique features of FTM-PVCs were uncovered in this study: (1) while the gene signatures of FTM- and TERM-PVCs were similar, key differences were observed, namely, that the Oct4A and Sox17 proteins were detected in FTM-PVCs, but not in TERM counterparts; (2) FTM-PVCs exhibited a greater proliferative potential; and (3) FTM-PVCs were more efficient in their in vitro differentiation toward selective mesenchymal cell types, including the chondrogenic and adipogenic lineages, as well as toward neuronal- and hepatocyte-like lineages, when compared to TERM-PVCs. Both PVCs were able to generate osteocytes and cardiomyocyte-like cells with similar efficiencies in vitro. Overall, FTM-PVCs show more plasticity than TERM-PVCs with regard to fate acquisition, suggesting that a restriction in multipotentiality is imposed on PVCs as gestation progresses. Taken together, our findings support the idea that PVCs from earlier in gestation may be better than later sources of multipotent stromal cells (MSCs) for some regenerative medicine applications.
Introduction
M
Recent molecular and phenotypic comparisons have revealed important differences between MSC sources, which may impact their utility [13 –16]. Therefore, the growth, immunological, paracrine, and differentiation properties of MSCs from different sources need to be defined to identify the best source for specific cellular therapies. Cells derived from the perivascular region of the human umbilical cord (HUC) [perivascular cells (PVCs)] represent an accessible and promising source of young MSC populations for autologous or allogeneic cell therapy. Notably, previous studies have shown that PVCs have a greater proliferative potential, capacity for osteogenic, chondrogenic, and adipogenic differentiation when compared to BM-derived MSCs (BM-MSCs) [17]. Here we hypothesized that PVCs have increased stem-like properties in earlier stages of gestation, and that fate restrictions are imposed on PVCs during fetal development, thereby decreasing their multipotency. As such, PVCs isolated from early stages of gestation could be more effective than term counterparts, and by correlation BM-MSCs, for some regenerative medicine applications. We present our findings on the comparison of molecular growth and in vitro differentiation potential of PVCs derived from early [first trimester (FTM)] and late (TERM) gestational stages.
Materials and Methods
Isolation and culture of HUC-PVCs and cell culture
TERM-PVCs were isolated as previously described [18] from term newborns (n=9) and obtained with informed consent through Tissue Regeneration Therapeutics (TRT) or Life Line Stem Cell. HUCs from FTM (8–12 weeks of gestation; n=13) were obtained from consenting patients who underwent elective pregnancy termination (REB No. 454-2011; Sunnybrook Research Institute). To isolate FTM-PVCs, the whole FTM umbilical cord was digested with 1 mg/mL collagenase type I (Sigma) for 2 h at 37°C. The denuded vessels were discarded after removal of PVCs during digestion. The cell suspension was diluted with phosphate-buffered saline (PBS; Gibco) and centrifuged at 250 g for 5 min. The pellet was resuspended and seeded into six-well tissue culture plate wells with alpha MEM (Gibco) supplemented with 10% fetal bovine serum (FBS; Hyclone, Lot No. KTJ32091), 2 mM L-glutamine, and 300 ng/mL amphotericin B (Sigma) at 37°C in 5% CO2. Subconfluent (80%–90%) cells were dissociated by treatment with 0.05% trypsin-EDTA (Gibco) and replated at a density of 2.4×103 cells/cm2. The culture medium was replaced every 3 days.
Flow cytometry
For flow cytometric analysis of freshly isolated TERM-PVCs, red blood cells were first depleted using the RBC lysis solution (Miltenyi Biotech, Inc.). For analysis of cultured cells, single-cell suspensions were obtained by dissociation with TrypLE (Invitrogen) at 37°C for 5 min and resuspended in 1% FBS/PBS. The cells were filtered through a 70-μm cell strainer (BD Bioscience). For intracellular staining of Oct4A, Sox2, Sox17, HLA-G, Nanog, albumin, and alpha-fetoprotein (AFP), the Cytofix/Cytoperm buffer (BD Bioscience) was used in accordance with the manufacturer's instructions. Antibodies used are listed in Supplementary Materials and Methods section (Supplementary Data are available online at
Immunohistochemistry and immunocytochemistry
All immunohistochemistry (IHC) was conducted at The Centre for Modeling of Human Disease, Pathology Core, Mount Sinai Hospital (Toronto, ON, Canada). For Oct4, CD146, and NG2 IHC, umbilical cords (n=2 and n=3, FTM- and TERM-HUC, respectively) were fixed with 4% paraformaldehyde for 2 h, and embedded in paraffin. For SSEA4 IHC, FTM and TERM umbilical cords (n=3 and n=2, FTM- and TERM-HUC, respectively), were set in Tissue-Tek O.C.T compound (Sakura Finetek) and snap-frozen in liquid N2. Serial 20-μm cryosections were obtained, and blocked in 0.3% hydrogen peroxide for 30 min and in protein block (Dako) for 30 min, before incubation at 4°C overnight in a primary antibody (Supplementary Materials and Methods section). The ABC kit (Vector Labs) and DAB chromogen were used for detection. Hematoxylin counterstaining was performed. Images were captured at 40×bright field using The Hamamatsu Whole Tissue Slide scanner (Olympus) and analyzed using NanoZommer 2.0 RS software.
For immunocytochemistry, cultured cells were rinsed twice before fixation with 4% paraformaldehyde for 20–30 min, and then permeabilized with 0.5% saponin in PBS. The cells were blocked with 10% normal goat serum (Jackson Immunoresearch Laboratories) in PBS for 30 min and incubated at 4°C overnight with primary antibodies (Supplementary Materials and Methods section). The cells underwent three 5-min washes with PBS and were incubated at room temperature for 1 hr with secondary antibodies (Supplementary Materials and Methods section). The nuclei were counterstained with Hoechst (1:1,000; Invitrogen) for 10 min followed by three 5-min washes with PBS. Fluorescence images were observed and captured using the EVOS fluorescence microscope (AMG).
Cell proliferation assay
The growth properties of FTM- and TERM-PVCs were analyzed by direct cell count to determine the log and lag phases. The cells were seeded in six-well tissue culture plates, at a cell density of 2.4×104/well [17], and viable cells were counted using trypan blue dye exclusion every 24 h over 7 days using the Countess® Automated Cell Counter (Life Technologies). Accumulated cell numbers and population doubling times (PDT) were assessed by plating 2.4×104 FTM-PVCs and TERM-PVCs (passage 2) into six-well tissue plates and calculated using the following algorithm:
PDT=t×log2/(logN t−logN 0), t=the culture time (hours), where N 0 is the number of cells seeded, and N t is the number of cells harvested. The cells were dissociated by trypsin-EDTA and replated every 7 days with medium replacement every 2 days. The expanded number of live cells over nine passages was counted and plotted from passage 2 to 9.
Karyotyping analysis
GTG-banding analysis was performed by the Procrea Cliniques Cytogenetics Laboratory, Montreal, QC. Four hundred bands per haploid chromosome in 3 cells were analyzed per sample (n=7 at passage 3, n=3 at passage 9).
Clonogenic assays
One hundred ninety two single cells from passage 6 FTM-PVC (n=5) and TERM-PVC (n=4) lines were seeded in 96-well plates (Nunc) using an AriaII-GC BRU cell sorter (SickKids UHN Flow Cytometry Facility, Toronto, Ontario) and cultured in maintenance media for 15 days. Media were changed every 4 days. Cell morphology and density were documented using bright field microscopy (Olympus).
In vitro multilineage differentiation assays
At least three independent FTM-PVC and TERM-PVC lines (passage 6 unless otherwise stated) were used for multilineage differentiation assays. P3 to P6 were determined to be ideal for differentiation assays based on amplification of lines and maintenance of proliferative and immunophenotypical properties. For all assays, the medium was changed every 3 days unless otherwise indicated.
Adipogenic
FTM-PVCs and TERM-PVCs were cultured as a monolayer in the presence of adipogenic supplements (STEMCELL Technologies) or based on [19], for 3 weeks, at which point they were fixed with 10% formaldehyde and stained with Oil Red O (Sigma) to detect lipid droplets. The dye from lipid droplets was extracted by isopropanol elution and quantified by spectrophotometer at A540 and the percentage of area stained with Oil Red O was measured by using the ImageJ analysis program.
Osteogenic
To induce osteogenic differentiation, FTM-PVCs and TERM-PVCs were seeded at 5×103 cells/cm2 and incubated in the low-glucose DMEM for 4 days, followed by culture in the Osteogenesis Differentiation Medium (Invitrogen) for 21 days. On day 21, cell were processed for Alizarin Red S staining to assess the induction efficiency and quantified by spectrophotometry at A405.
Chondrogenic
For chondrogenic differentiation, micromass cultures were generated by seeding 5-μL droplets of cell solution (1.6×107 cells/mL) in the center of wells. These were incubated at 37°C for 2 h, and then cultured in the Chondrogenesis Differentiation Medium (Invitrogen) for 14 days. Chondrogenic pellets were processed for Alcian Blue staining and positively stained pellets were counted and captured using bright field microscopy (Olympus).
Cardiomyogenic differentiation
For cardiogenic differentiation, three independent FTM- and TERM-PVC lines (passage 3 and 4) were cocultured with neonatal rat cardiomyocytes. Neonatal rat cardiomyocytes were obtained and prepared as previously described [20]. The isolated cardiomyocytes were replated at 5×104 cells/cm2 to a 3.0-μm pore size collagen-coated membrane transwell (Corning) that is only permeable to secreted molecules and incubated for 24 h. About 5×104 FTM- or TERM-PVCs were seeded to the bottom well and cocultured for 14 days. The medium was replaced every 2 days. The cardiomyogenic induction rate was calculated as the percentage of cardiac troponin T-positive (cTnT+) or alpha-sarcomeric actinin-positive (α-SA+) cells over total cells (Hoechst-positive nuclei) in randomly selected fields.
Hepatic differentiation
Hepatic differentiation of FTM-PVCs was performed using a modified two-step protocol based on [21,22]. Briefly, upon confluence, independent lines of FTM-PVCs (n=4) and TERM-PVCs (n=3) each at passage 5 and passage 6 were treated with the differentiation medium consisting of the DMEM (low glucose) supplemented with HGF (50 ng/mL; PeproTech), bFGF (10 ng/mL), dexamethasone (10−8 M), and ITS (50 mg/mL) for 16 days (Step 1). Cells were then induced with the maturation medium consisting of the DMEM/F12 medium supplemented with oncostatin M (20 ng/mL; PeproTech), dexamethasone (10−6 M), and ITS (50 mg/mL) for 16 days (Step 2). The medium was changed every 4 days. Cells were harvested for flow cytometry and QPCR analysis at day 8, 15, 22, and 32. Morphological changes were documented every 4 days using bright field microscopy (Olympus).
Neurosphere assay
The neural colony-forming assay was performed as described previously [23], with minor adaptations for PVCs. In brief, independent lines of FTM-PVCs (n=10) and TERM-PVCs (n=4) at passage 5 or 6, were harvested using either trypsin-EDTA or TrypLE, washed in serum-free media: TESR1 or 2 (STEMCELL Technologies) or NeuroCult media (STEMCELL Technologies), and then plated in the same serum-free media containing either human LIF (1,000 U/mL, Millipore) or ROCK inhibitor (Y-27632, STEMCELL Technologies; 10 μg/mL) and a combination of epidermal growth factor (20 ng/mL; Santa Cruz Biotech), basic fibroblast growth factor (10 ng/mL, PeproTech), heparin (2 mg/mL; BioShop), and B-27 supplement (50×Gibco). Cells were allowed to grow for 5–14 days. Cell viability and quantification were assessed by trypan blue dye exclusion, with cells seeded at a concentration of 10 cells/μL to ensure clonality. On day 5–7, all spheres that were approximately≥50 μm, were counted, and then individually isolated and plated on a Matrigel substrate in TESR1 or 2 or NeuroCult media containing 1% FBS and incubated for 5–7 days to assess their differentiation potential.
Low-density lipoprotein uptake assay
Adherent cells were rinsed with PBS and incubated with 10 μg/mL of 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine-labeled acetylated low-density lipoprotein (Dil-ac-LDL) (1:100; Molecular Probes) in prewarmed media for 4 h at 37°C in 5% CO2 and washed with 1% FBS/PBS, according to the manufacturer's instructions. Dil-ac-LDL uptake was observed and captured using an EVOS fluorescence microscope (AMG).
RNA isolation, RT-PCR, and QPCR
RNA samples were prepared using the RNAeasy kit (Qiagen) according to the manufacturer's instructions, followed by DNase treatment to eliminate traces of genomic DNA. RNA was reverse transcribed into cDNA using the RT2 First Strand Kit (Qiagen) according to the manufacturer's instructions. For QPCR, commercial Human RT2 Profiler™ PCR Arrays (PAHS-405R and PAHS-508R; SABiosciences) and QPCR primer assays (SABiosciences; Supplementary Table S1) were used. The experiment was performed following the manufacturer's instructions on the Rotor-gene 6000 instrument (Corbett) using 20–25 ng total cDNA per reaction. For the QPCR arrays, gene expression levels were determined using PCR Array Data Analysis Centre (SABbiosciences) using the delta–delta Ct (ΔΔCt) method and reported as the fold change relative to TERM-PVCs. NTERA2 cells were used as a positive control. For the QPCR primer assay analysis, standard curves were generated using positive control cDNA dilutions and the PFAFFL method was used to report fold change between undifferentiated and differentiated FTM-PVCs and TERM-PVCs. GAPDH or RPL13A were used as normalizers. All assays were done in triplicates for at least three independent PVC lines (at passage 4 for arrays; passage 6 for lineage differentiation QPCR primer assays) in all conditions tested, with the exception of neural differentiation when n=1 representative line. Genes with normalized Ct≥33 were considered as not detected.
Data analysis
All results were generated from at least three independent experiments and were presented as mean±SD, unless otherwise indicated. Statistical significance was determined using the Student's t-test, unless otherwise indicated, and differences were considered significant when P<0.05.
Results
A predominant population of cells with mesenchymal and pericytic characteristics is maintained in FTM-PVC and TERM-PVC cultures
Cells isolated and cultured from the perivascular region of HUCs at birth express the combination of cell surface markers that define MSC populations [24,25]. We asked if cells isolated from FTM-PVCs have similar immunophenotypical characteristics. IHC of FTM and TERM umbilical cord tissue sections shows a discrete population of cells surrounding the endothelial cell layer expressing NG2 and high levels of CD146, two pericyte markers. CD146 expression was also detected in the adjacent smooth muscle cell layers, with weaker staining in the connective tissue (Fig. 1A). Flow cytometric analysis of cells freshly isolated from FTM umbilical cords revealed that greater than 98% of cells expressed the representative MSC marker CD90 at the time of isolation, and 73%±2.2% also expressed the pericyte marker CD146 (11.4%±0.7% CD146+bright and 61.6%±2.1% CD146+dim). Endothelial and hematopoietic markers, CD31 and CD45, were not detected (Supplementary Fig. S1). When freshly isolated TERM cells were analyzed for the same markers, ∼90% were CD90+and ∼15% expressed CD146 (4.9%±0.3% CD146+bright and 10.8%±0.3% CD146+dim) (Supplementary Fig. S1). A heterogeneous population, including an enriched source of pericyte-like cells can thus be obtained from the perivascular region of the FTM umbilical cords, and is in accordance with previous reports on term umbilical cords [25]. We then wanted to assess the immunophenotype of cultured FTM-PVCs in greater detail and in comparison to previously characterized term counterparts [24]. Cells harvested from FTM (n=13, 9.1±1.1×105 cells/cord) and TERM umbilical cords (n=9, 1.9±1.99×106 cells/cord) were plated in standard MSC culture conditions. While more cells were harvested per cm of FTM versus TERM cords (P<0.05, Fig. 1B), all lines of PVCs were maintained as monolayers for at least nine passages and exhibited homogeneous elongated spindle-like cell morphologies. Flow cytometric analysis at passage 1 showed that PVCs were negative for the hematopoietic (CD45, CD117) and endothelial (CD34 and CD31) cell markers (Fig. 1C and Supplementary Fig. S2A). The MSC-associated markers CD44, CD73, CD90, and CD105 were all detected on greater than 90% of total cell populations in both culture types. A major proportion of both FTM- and TERM-PVCs were positive for the pericyte-associated markers CD146 (90.4±10.3 vs. 85.6±0.4 CD146low), NG2 (65.9±5.6 vs. 61.1±10.7), and PDGFRβ (91.7±4.2 vs. 75.8±9.7) (P>0.05) (Fig. 1C, and Supplementary Fig. S2A). The majority of FTM- and TERM-PVCs express low levels of the immunogenic MHC class I molecules HLA-A,B,C (82.6%±6.5% vs. 93.5%±0.8%, respectively, P>0.05), although we suspect that this antibody cross-reacts with the immunoprotective molecule HLA-G [26]. HLA-G was detected in ∼70% of the cells in FTM- and TERM-PVC cultures, while HLA-DR/DP/DQ molecules were not detected (Fig. 1C and Supplementary Fig. S2A). PVCs were maintained for at least nine passages without any significant alterations in their immunophenotypes (Supplementary Fig. S2B). These results indicate that cultured FTM- and TERM-PVCs are very similar with regard to their mesenchymal, pericytic, and immunological molecular profiles and these properties are maintained in culture, with the notable exception of the CD146bright population, which is detected in freshly isolated PVCs, but lost or altered in standard MSC culture conditions (Supplementary Figs. S1 and S2).

Isolation and phenotypic profiling of FTM-PVCs.
The pluripotency marker Oct4A is expressed in cultured FTM-PVCs, but not TERM-PVCs
We next investigated the expression of pluripotency- and stem cell-associated markers in FTM and TERM umbilical cord tissue sections (in situ), in freshly isolated cells, and in cells cultured under standard MSC conditions. Sox2, Nanog, TRA-1-60, and TRA-1-81 protein expression were examined by IHC in tissue sections of FTM- and TERM- umbilical cords, but were not detected (data not shown). SSEA4 was detected in a subpopulation of PVCs in both FTM and TERM umbilical cords, while nuclear and cytoplasmic Oct4 was detected in PVCs of FTM, but not TERM umbilical cords (Fig. 2A). Flow cytometric analysis using SSEA4- and Oct4A-specific antibodies revealed that ∼8% vs. 20%–30% of freshly isolated FTM and TERM cells expressed SSEA4, and 85% vs. 0%–2.5% expressed Oct4A, respectively. We confirmed that the Oct4A+cells observed in some TERM samples were CD90(−) (Supplementary Fig. S1). Rare cells expressing Sox2 (1.86±0.1), TRA-1-60 (0.87±0.04), and TRA-1-81 (0.71±0.07) were also detected by flow cytometry in freshly isolated FTM cells (Supplementary Fig. S1). These proteins could not be detected by flow cytometry or immunocytochemistry in cultured PVCs, suggesting that this potential rare progenitor population is lost in standard MSC culture approaches (Supplementary Fig. S3).
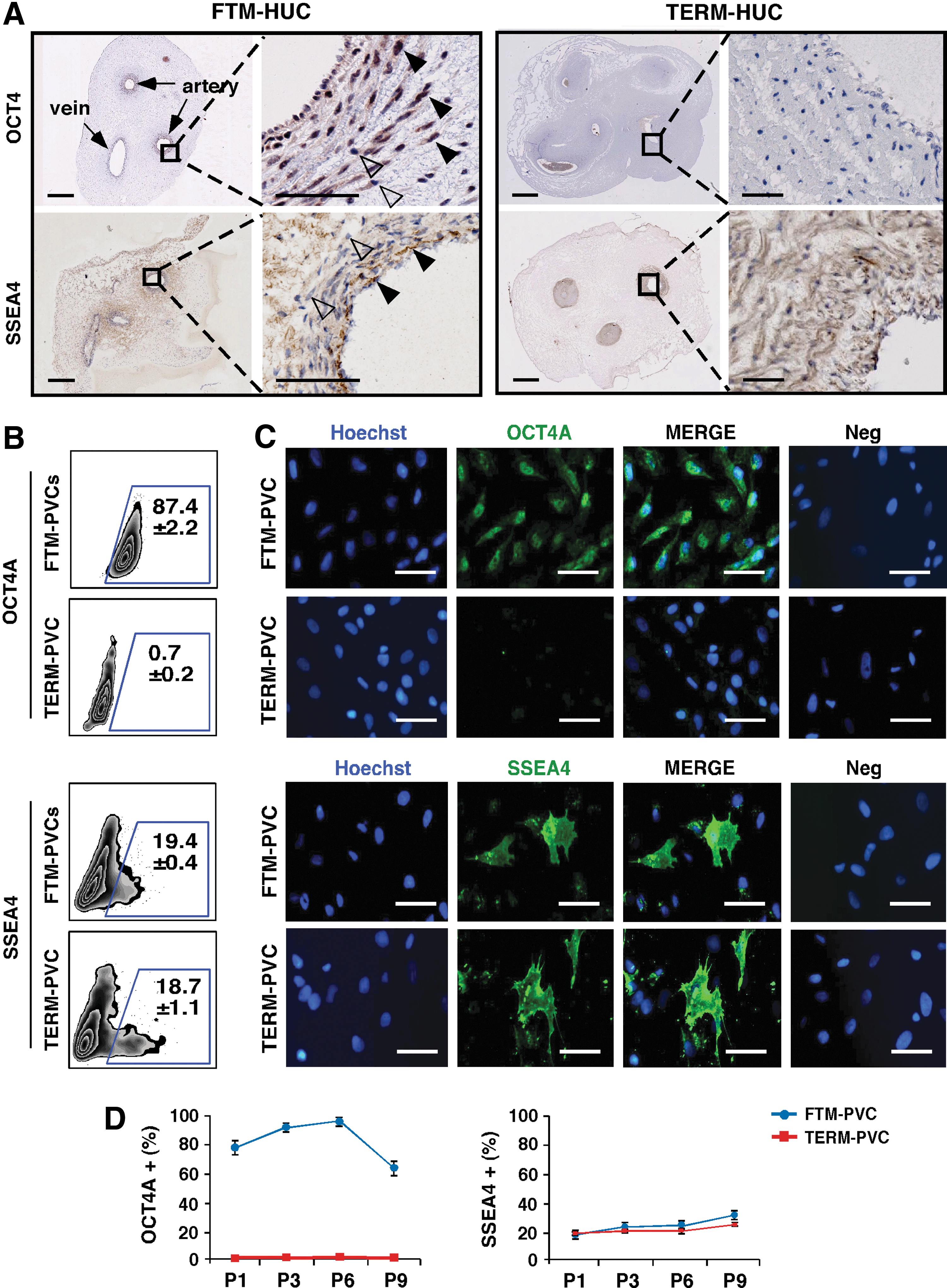
Comparison of stem cell-associated marker expression between FTM-PVCs and TERM-PVCs.
In culture (passage 1), the frequency of SSEA4+cells in FTM-PVCs and TERM-PVCs was comparable and represented ∼20% of PVCs (Fig. 2B and Supplementary Fig. S3), while low levels of the Oct4A protein were exclusively detected in a high frequency of FTM-PVCs (87.4±2.2 vs. 0.7±0.2 in TERM) (Fig. 2B and Supplementary Fig. S3). These findings were confirmed by immunocytochemistry of cultured FTM- and TERM-PVCs (Fig. 2C). Importantly, flow cytometric analysis revealed that the frequency of Oct4A+and SSEA4+populations in FTM- and TERM-PVC cultures were maintained from passage 1 through passage 9, although there was a trend toward reduction of the Oct4A+population between passage 7 and 9 in FTM-PVC cultures (P>0.05, ANOVA) (Fig. 2D). In summary, while a population of SSEA4+cells is maintained in both FTM- and TERM-PVC cultures, only FTM-PVCs express the pluripotency factor the Oct4A protein in vivo and in culture.
FTM-PVCs have a greater proliferative and clonogenic capacity than TERM-PVCs
We next compared the proliferative potential of FTM-PVCs to TERM-PVCs. At passage 2, both FTM-PVCs and TERM-PVCs showed similar growth patterns, with a lag phase for the initial 48 h followed by a log phase from 72 to 144 h. However, the cell count was significantly higher in FTM-PVCs (P<0.05) at each time point from 96 to 144 h when compared to TERM-PVCs (Fig. 3A). The proliferation rates in both cell types were consistent until passage 6, and then significantly decreased at passage 8 (P<0.05, ANOVA). However, the proliferative capacity of FTM-PVCs was significantly higher compared with TERM-PVCs at all passages analyzed (P<0.05) (Fig. 3B). The extrapolated analysis for the accumulated cell number revealed that>10,000-fold more FTM-PVCs could be obtained in comparison to TERM-PVCs after 10 passages (P<0.01) (Fig. 3C). Tested lines displayed a normal chromosomal karyotype at passage 3 and at passage 9 (Fig. 3D). We also found that at least three times more FTM-PVCs were able to generate clones when compared to TERM-PVCs (20.5±3.0 vs. 6.25±5.2; P<0.05) (Fig. 3E, F). Thus, FTM-PVCs have a greater proliferative potential and clonogenic capacity than TERM-PVCs.

FTM-PVCs proliferate faster than TERM-PVCs.
FTM-PVCs have greater mesenchymal differentiation potential than TERM-PVCs
The abilities of FTM- and TERM-PVCs to differentiate into adipocytes, chondrocytes, and osteocytes were compared in vitro. Quantitation of the total relative amount of lipid vacuoles using Oil Red O staining and spectrophotometric analysis [17] revealed that FTM-PVCs generated significantly more lipid vacuoles than TERM-PVCs when cultured under adipogenic conditions (P<0.05) (Fig. 4A, B). Similarly, the percentage of total cell area positive for Oil Red O staining [27] rose to ∼6% in FTM versus ∼4% in TERM adipogenic induction (P<0.05) (Fig. 4C). Quantification of adipogenic transcripts peroxisome-proliferator-activated receptor gamma (PPARG) and lipoprotein lipase (LPL) levels in undifferentiated and differentiated PVCs showed increased PPARG expression upon adipogenic differentiation (Fig. 4D). However, FTM-PVCs expressed significantly higher levels of PPARG when compared to TERM counterparts in undifferentiated conditions as well as upon differentiation (Supplementary Table S2; P<0.05). LPL was not detected. FTM-PVCs also produced a significantly higher number of chondrogenic pellets when compared to TERM-PVCs (P<0.05) when cultured as micromass in chondrogenic conditions (Fig. 4E, F). The chondrogenesis-associated transcripts SOX9 and aggrecan (ACAN) are expressed at high levels in undifferentiated FTM- and TERM-PVCs (Supplementary Table S3) and, with the exception of ACAN upregulation by one of three FTM-PVC lines, were not significantly upregulated. Collagen type II alpha1 (COL2A1) expression was upregulated in both populations of PVCs upon chondrogenic differentiation (Fig. 4G). Alizarin S staining and spectrophotometry was used to quantify calcium deposits, a measure of osteocyte activity, in PVCs cultured under osteogenic culture conditions. The osteo-genic differentiation was robust and there was no significant difference in the quantity of calcium deposits between FTM-PVCs and TERM-PVCs at the end of differentiation (Fig. 4H, I). The osteogenic transcripts RUNX2 and BGLAP (Osteocalcin) transcripts are highly expressed in undifferentiated PVCs (Supplementary Table S4 and Table 1), and were upregulated during osteogenic differentiation in both FTM- and TERM-PVCs (Fig. 4J). Undifferentiated PVCs also express SPP1 (osteopontin) (Supplementary Table S4) and two of the three FTM lines upregulated SPP1 expression during osteogenic differentiation, while none of the TERM-PVCs did. FTM-PVCs showed a significantly higher increase in RUNX2 expression when compared to TERM-PVCs at the half-way point of differentiation (day 11) (P<0.005), but a comparable increase was observed at the end of differentiation (Fig. 4J). In summary, FTM-PVCs on average are more efficient at generating adipocytes, chondrocytes, and in some respects, osteocytes when compared to TERM-PVCs.
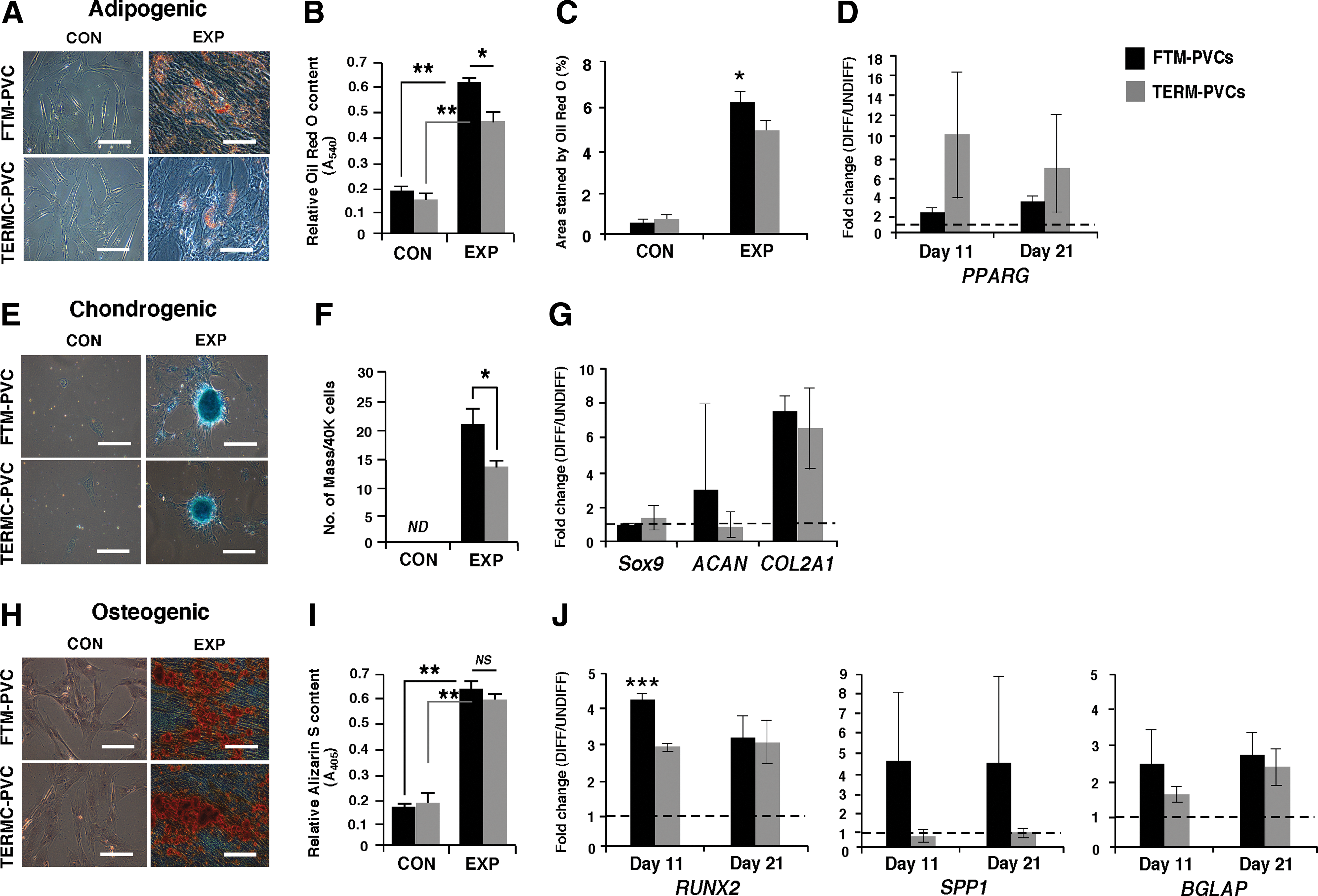
FTM-PVCs and TERM-PVCs can differentiate into three of the main mesenchymal lineage cell types.
Ct values were normalized to the housekeeping genes on the array (GAPDH, RPL13A, and HPRT1).
PVCs, perivascular cells; FTM, first trimester; SD, standard deviation.
FTM- and TERM-PVCs display similar cardiomyogenic potential
Undifferentiated FTM- and TERM-PVCs were cocultured with neonatal rat cardiomyocytes for 14 days to induce their differentiation into cardiomyocytes. With coculture, we observed a gradual loss of the typical elongated, spindle-like morphologies of PVCs and a corresponding increase in larger cells with cardiomyocyte-like morphological features (Fig. 5A). Undifferentiated and cocultured FTM- and TERM-PVCs were immunostained for the cardiomyocyte-specific markers: α-SA and cTnT on day 14 to assess cardiomyogenic induction efficiency. The basal levels of these markers in undifferentiated TERM-PVCs were significantly higher than in undifferentiated FTM-PVCs (P<0.05), but comparable proportions of α-SA- or cTnT-positive cells were observed in each PVC population after cardiomyogenic induction (Fig. 5B), representing an approximately threefold and eightfold increase in αSA and cTnT expression in FTM-PVCs, respectively, versus twofold and twofold corresponding increases in TERM-PVCs (P<0.05; P<0.001). No beating cells were observed. This data suggest that FTM- and TERM-PVCs have a significant cardiomyogenic potential, and that FTM-PVCs show more plasticity toward this lineage.
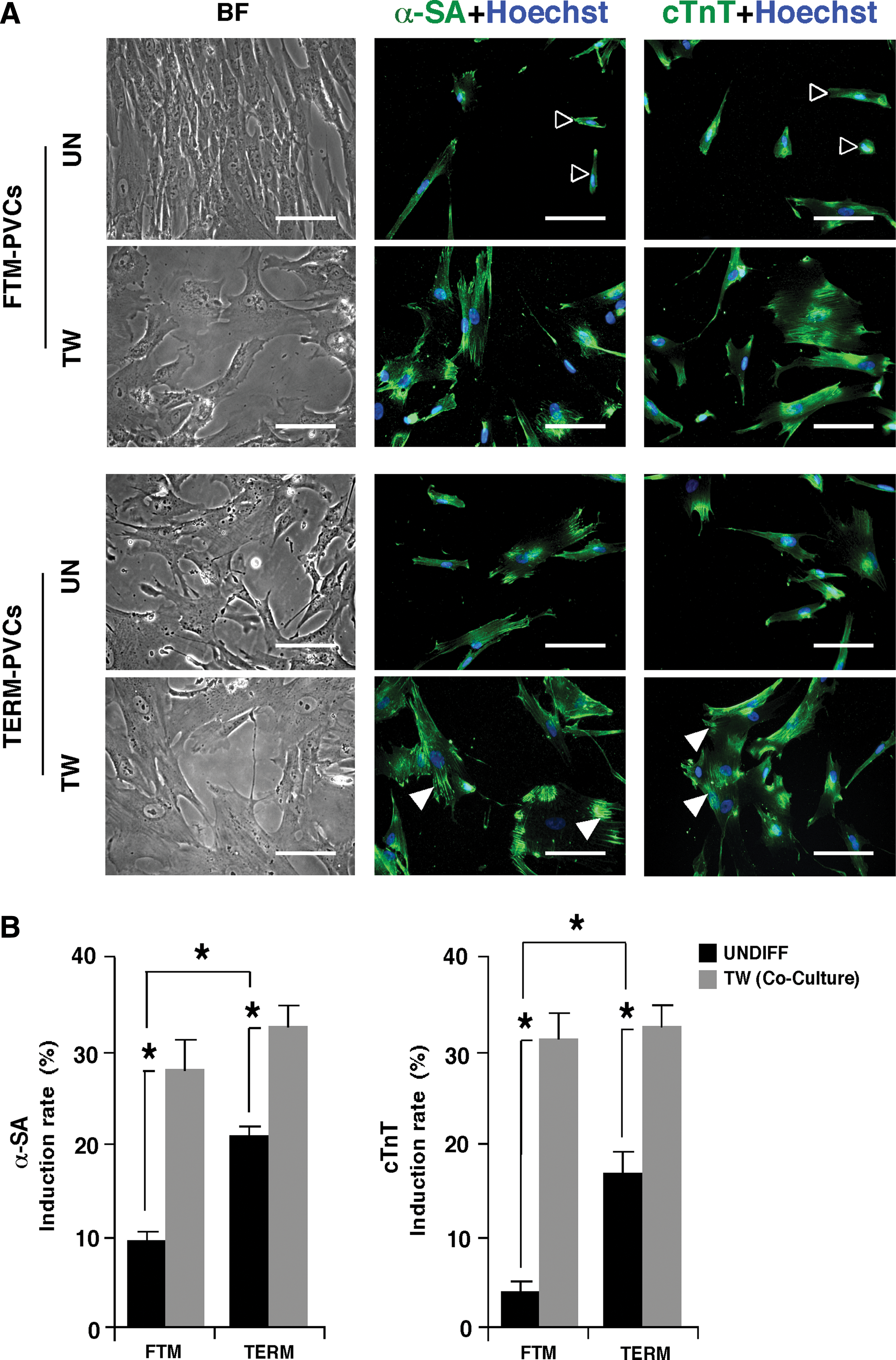
The cardiomyogenic potential of FTM- and TERM-PVCs.
FTM-PVCs transdifferentiate into hepatocyte-like cells more efficiently than TERM-PVCs
The ability of some populations of MSCs to differentiate along nonmesenchymal lineages has previously been reported, but remains controversial [12]. We compared the ability of FTM-PVCs and TERM-PVCs to differentiate toward the hepatic lineage using a two-step culture model. Early in the initiation step, the cell size of PVCs increased noticeably and the cells started to lose the typical fibroblast-like morphology observed in undifferentiated control cells. Round-shaped cells with a polygonal structure were gradually observed in the maturation step. The majority of FTM-PVC lines underwent this hepatocyte-like morphological change faster than TERM lines (Fig. 6A). With the exception of AFP, hepatic transcripts, including ALB, CYP3A4, and HNF1A are expressed at very low levels in undifferentiated PVCs and AFP and ALB transcripts levels are higher in TERM-PVCs when compared to FTM-PVCs (P<0.05; Supplementary Table S5). AFP, ALB, and HNF1A transcripts were significantly upregulated in FTM- and TERM-PVCs at the end of hepatic induction (Fig. 6B). FTM-PVCs showed a significantly greater fold change in ALB expression during the first step of differentiation, and an overall trend for greater upregulation of hepatic transcripts in two of three FTM-PVC lines when compared to TERM-PVCs. We analyzed the hepatic phenotype and function of the differentiated cells at day 32 by performing immunocytochemistry for the hepatic marker AFP (Fig. 6C). Quantification of the percentage of AFP-positive cells that also had a polygonal morphology at day 0 and 32 showed that an increased proportion of the FTM-PVCs had transdifferentiated into hepatocyte-like cells when compared to TERM-PVCs (50.3%±2.4% vs. 23.6%±1.8%) (P<0.01) (Fig. 6D). Flow cytometric analysis of cellular AFP and albumin protein levels showed an increase between day 0 and 32 in FTM-PVCs cultured under hepatic conditions (Supplementary Fig. S4). The majority of induced FTM-PVCs showed Dil-ac-LDL uptake, whereas undifferentiated cells did not (Supplementary Fig. S4). Thus, PVCs have the potential to transdifferentiate to hepatocyte-like cells in vitro, and FTM-PVCs do so more efficiently than TERM-PVCs.
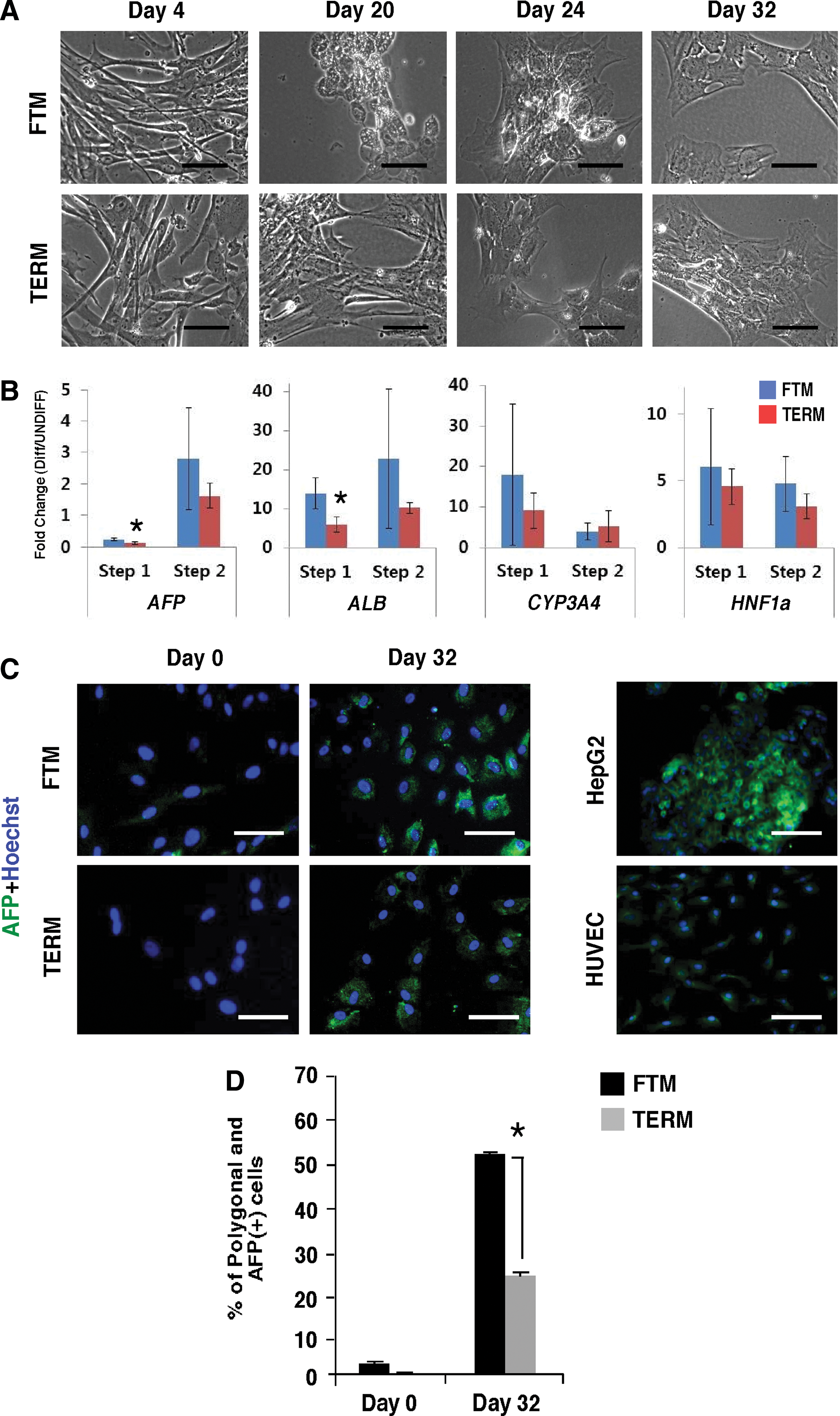
The hepatic differentiation potential of FTM- and TERM-PVCs.
FTM-PVCs have greater neural potential than TERM-PVCs
To assess whether PVCs have any proclivity toward the neural ectodermal lineage, we performed QPCR analysis and immunocytochemistry for neural markers on undifferentiated FTM- and TERM-PVCs. QPCR analysis showed that FTM- and TERM-PVCs express high levels of NESTIN, neural cell adhesion molecule (NCAM), and βIII-tubulin (TUBB3), but that glial fibrillary acidic protein (GFAP) expression is outside considerable detection limits (Supplementary Table S6). We observed significantly increased levels of NESTIN and TUBB3 transcripts in FTM-PVCs when compared to TERM counterparts (P<0.05) (Supplementary Table S6). By immunocytochemistry, the occasional cells showed weak, but detectable expression of the neuronal marker neurofilament M, but the neuronal marker TUBB3 or the astrocyte marker GFAP were not expressed in undifferentiated FTM- or TERM-PVCs. Both FTM- and TERM-PVC cultures showed high expression of the Nestin protein, a neural progenitor-associated marker also expressed in pericytes (Fig. 7A). We then performed a modified neurosphere assay and found that FTM-PVCs formed spheres ∼13 times more efficiently than TERM-PVCs (P<0.05, Fig. 7B, C). We were able to pool spheres to compare neural gene expression in both FTM- and TERM-PVC-derived spheres, and found an upregulation of NESTIN and NCAM, but not TUBB3 gene expression upon sphere formation in FTM-PVCs; NCAM upregulation was significantly greater when compared to TERM-PVCs (P<0.001). TERM-PVC-derived spheres displayed upregulated NESTIN, but not NCAM or TUBB3 expression (Fig. 7D). Upon further differentiation on Matrigel, cells migrating out of FTM-PVC-derived spheres exhibited strong expression of Nestin and TUBB3, with some cells showing neuronal-like morphology. TERM-PVCs seldom formed spheres, and the few spheres that were isolated rarely underwent differentiation or expressed TUBB3 (Fig. 7E). Taken together, these findings suggest that FTM-PVCs have increased neuronal potential when compared to TERM-PVCs.
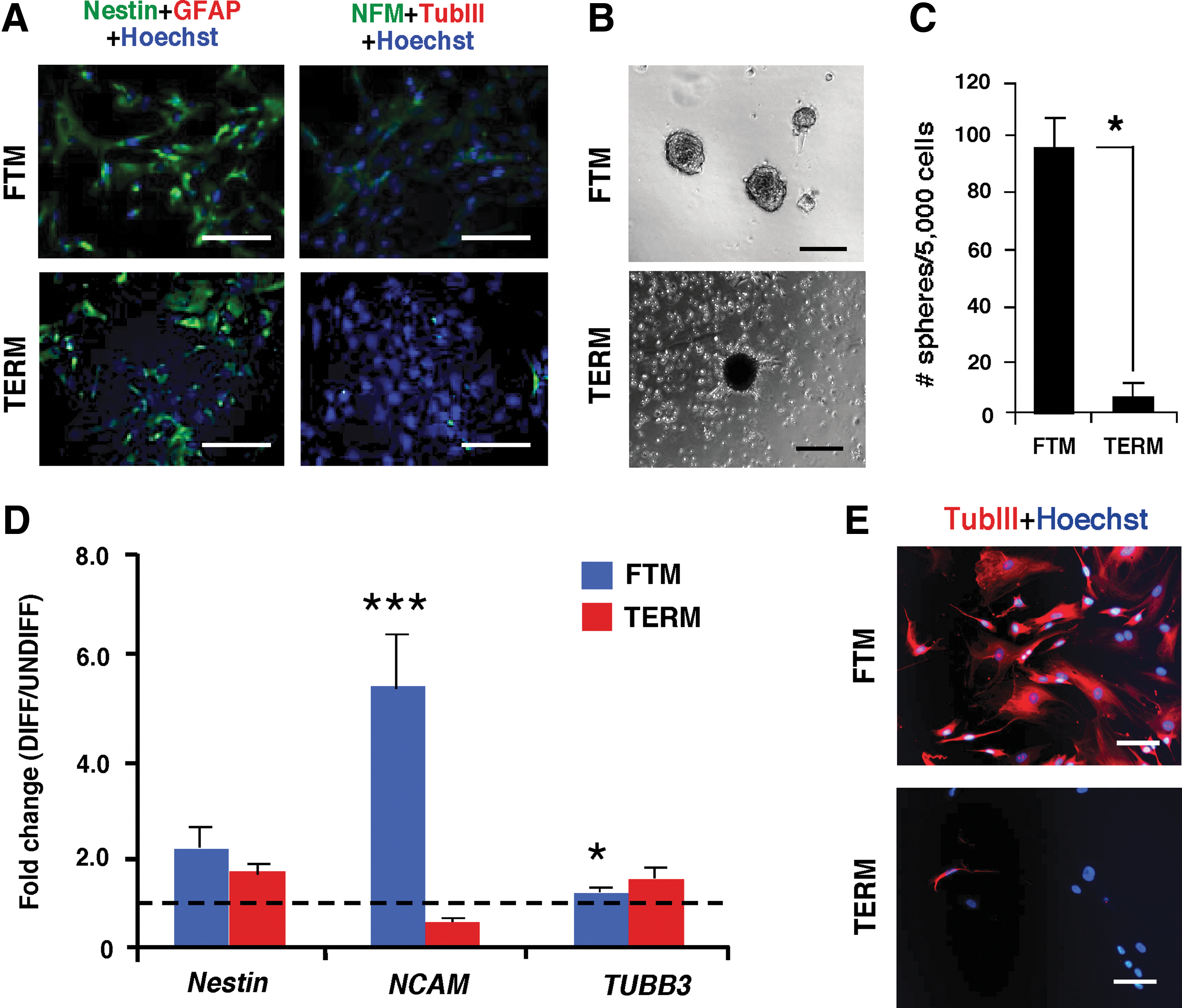
The neural potential of FTM-PVCs.
FTM- and TERM-PVCs differ in gene expression
To ask if molecular differences pointing to proliferation or lineage potential biases exist between cultured FTM- and TERM-PVCs, we compared the transcriptional profiles of PVCs using commercially available quantitative PCR-based arrays. One hundred sixty two genes related to cell lineage, stemness, and cell cycle regulation were profiled. Six transcripts were found to be significantly upregulated in FTM-PVCs when compared to TERM-PVCs, including FGF5, PPARG, SMTN, TUBB3, DNMT3B, and PODXL; DCN was found to be significantly downregulated (P<0.05) (Table 1). Differences in an additional set of 18 transcripts encoding growth factors and cytokines, cyclins and developmental transcription factors were found to be differentially expressed between FTM- and TERM-PVCs, but failed to reach significance (P>0.05) (Table 1). Another set of sixty genes were found to be expressed in both FTM-PVCs and TERM-PVCs, at equal levels and include the marker CD44, transcriptional factor RUNX1 as previously identified in umbilical cord-derived PVCs [25], members of the tyrosine kinase receptor and fibroblast growth factor families, Notch, Wnt, and BMP signaling pathways (see Table 1 and Supplementary Table S7 for full list). We were particularly interested in evaluating the expression levels of genes on the array that are associated with pluripotency such as SOX2, NANOG, and OCT4/POU5F1, as previous groups have reported expression of these transcripts in fetal sources of MSCs [28]. While OCT4/POU5F1 transcripts were detected at low levels in both FTM-PVC and TERM-PVC cultures with some FTM lines expressing higher levels, SOX2 and NANOG transcripts were detected at levels that we consider outside considerable expression levels (Ct>33) in either PVC populations (Supplementary Table S7); this is in accordance with our protein expression data and previous reports [25]. One caveat to this experiment is that high Ct values can reflect low to nondetectable transcript expression in PVCs, or low expression by subpopulations of the MSC lineage. Because of its role in endodermal differentiation, we validated the protein expression of SOX17, which appears to be expressed at low levels, but is upregulated in FTM-PVCs (average Ct 31) when compared to TERM-PVCs (average Ct 34). Using flow cytometric analysis, we found that 9.03%±0.48% of FTM-PVCs expressed detectable levels of the Sox17 protein, while TERM-PVCs did not (0.03±0.005), P<0.001 (Supplementary Fig. S5). Thus, a subpopulation of FTM-, but not TERM-PVCs expresses Sox17.
Discussion
It has recently been reported that young sources of adult stem cell populations show greater proliferative potential and plasticity than older counterparts [29,30] and, for this reason, may have more potential for cell therapy. This study is the first to characterize a population of PVCs derived from the FTM umbilicus. We directly compared the phenotypic, growth and differentiation properties of these cells to ontogenetically related TERM-PVCs [24]. Our data support five major conclusions. First, FTM-HUCs are a richer source of pericyte-like cells. Second, conventional MSC culture conditions support FTM-PVC and TERM-PVC populations, where the majority of cells display immunophenotypes corresponding to a mesenchymal lineage with pericyte-like characteristics, but do not support the potentially interesting CD146high population or small pluripotent factor-expressing population found in freshly isolated PVCs. The impact of this, if any, on functional properties of PVCs is not yet understood. Third, our molecular investigations indicate that while FTM-PVCs and TERM-PVCs share similar gene expression signatures, they differ in key genes that are known to regulate pluripotency, cell fate, and differentiation in stem/progenitor cells. Namely, the majority of the FTM-PVCs express the pluripotent factor Oct4A and ∼10% express Sox17, whereas TERM-PVCs do not. Fourth, FTM-PVCs show greater proliferative and clonogenic potential than TERM-PVCs, with a greater than 1.3-fold faster average PDTs at all passages analyzed, and greater than threefold ability to generate clones when plated as single cells. Finally, FTM-PVCs show ∼30% greater in vitro differentiation toward selective mesenchymal lineages, including adipogenic and chondrogenic lineages, and toward non-mesenchymal lineages such as the neuronal and hepatic lineages when compared to TERM-PVCs. Osteogenic differentiation and cardiomyogenic-like differentiation were equally robust between the FTM- and TERM-PVCs but, overall, FTM-PVCs show more plasticity with regard to fate acquisition when compared to TERM-PVCs. Thus, a natural restriction in proliferative capacity and multipotentiality is imposed on PVCs as gestation progresses.
The primitive nature of perinatal sources of MSCs such as the umbilical cord, placenta, and amniotic sac is a relatively newly explored concept and alludes to the reported expression of pluripotency markers in these cell types, as well as to their increased proliferative and differentiation capacities when compared to adult sources of MSCs such as the BM [28,31]. The expression of pluripotency markers such as Oct4A, SSEA4, Sox2, and Nanog have been reported (mainly at the transcriptional level) and varies in MSCs derived from a variety of tissues, but their function in the regulation of MSC proliferative and differentiation capacities remains unclear [28,32 –35]. Of course, an important caveat to any comparative MSC study, including this one, is the inherent tissue structure and size differences between MSC sources that preclude the use of identical isolation protocols and, which may, in turn, have important effects on adherence, growth kinetics, and multipotential characteristics [11]. Nonetheless, the molecular studies presented in this manuscript and others support several mechanistic explanations for the increased proliferative capacity as well as the increased plasticity of cultured FTM-PVCs in comparison to their mature counterparts, the major one being a dynamic expression of stem cell-associated markers such as Oct4A during umbilical cord tissue development. Oct4A plays an essential role in maintaining self-renewal and pluripotency of ESCs and cord blood-derived MSCs [36,37], promotes cell proliferation and colony formation in MSC culture [33] and may do this by targeting genes that regulate cell proliferation and plasticity [38]. We and others have shown that, while OCT4 transcripts are detected in both FTM and TERM-PVC cultures, the Oct4A protein is only expressed in early PVCs, decreases by the second trimester (data not shown), and is not detectable in TERM-PVCs [24,25]. Similarly, Oct4A expression tends to decrease after passage 7 in cultured FTM-PVCs. Interestingly, there appears to be a correlation between OCT4 levels and proliferation rates as well as plasticity in FTM-PVCs, where we observed increased OCT4/Oct4A levels in the FTM-PVC lines showing the highest proliferation rates and best differentiation capacities toward all lineages. On this note, the inter-line variability in lineage differentiation efficiencies and gene expression levels was clearly observed between FTM-PVCs and cannot be solely attributed to gender- or gestational age-related factors (data not shown). The Oct4A-expression level may thus be a useful marker to select or enrich for the best FTM lines to use in in vivo regenerative experiments. The gene expression differences between FTM- and TERM-PVCs could explain their differences in cell cycle progression, cell fate and lineage differentiation potential, but may also reflect intrinsic differences in the hierarchy and heterogeneous MSC progenitor populations found within each culture. While this model remains to be demonstrated in appropriate clonal experiments, the heterogeneity of cultured PVCs may present particular advantages for cellular therapy applications requiring the regeneration of the many cell types present in an organ or tissue.
The mechanisms regulating ontogenic changes in PVCs during gestation (for PVCs derived from the umbilical cord, placenta, or amnion) and human development (for other sources of PVCs) remain largely unexplored, but are likely to involve epigenetic mechanisms such as DNA methylation and chromatin modification. Such mechanisms may regulate changes in the levels of commitment in perivascular progenitor cells during pregnancy, and be influenced by the maternal/fetal circulation and hormones or, in an in vitro context, by the culture environment. We are currently investigating alternative isolation and culture conditions to better preserve or enrich for the pericytic and multipotential characteristics of PVCs and optimize their growth and secretory properties for their future use in clinical applications. Our future studies will also focus on evaluating these cells in in vivo models of pathology and injury to determine if they can directly and/or indirectly contribute to repair.
Footnotes
Acknowledgments
We would like to gratefully acknowledge all of the CReATe Fertility Centre research team members for their support, in particular, Stephanie Grover, Katya Park, and Tanya Barretto. We thank the Morgentaler clinic staff as well as Dr. John Davies and Elaine Cheng of Tissue Regeneration Therapeutics (TRT) for their assistance in acquiring the umbilical cord tissue and PVCs, as well as the CReATe Cord Bank staff for their technical assistance. We also thank the Ren-Ke Li lab (University Health Network, Toronto) and Freda Miller lab (Hospital for Sick Children, Toronto) for their assistance with the rat cardiomyocyte cultures. Some of the data presented in this manuscript were previously published in abstracts by the International Society for Stem Cell Research (ISSCR) and the Canadian Fertility and Andrology Society (CFAS).
Author Disclosure Statement
Dr. Clifford Librach has filed patent applications in Canada, U.S., Mexico, and Australia for this invention entitled “Method of Isolation and Use of Cells Derived from First Trimester Umbilical Cord Tissue.” The applications are currently pending and awaiting examination. Dr. Clifford Librach is the owner and director of the CReATe Cord Blood Bank and Peristem™ program.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.