
Abstract
The nucleus pulposus (NP) plays a prominent role in both the onset and progression of intervertebral disc degeneration. While autologous repair strategies have demonstrated some success, their in vitro culture system is outdated and insufficient for maintaining optimally functioning cells through the required extensive passaging. Consequently, the final population of cells may be unsuitable for the overwhelming task of repairing tissue in vivo and could result in subpar clinical outcomes. Recent work has identified synovium-derived stem cells (SDSCs) as a potentially important new candidate. This population of precursors can promote matrix regeneration and additionally restore the balance of catabolic and anabolic metabolism of surrounding cells. Another promising application is their ability to produce an extracellular matrix in vitro that can be modified via decellularization to produce a tissue-specific substrate for efficient cell expansion, while retaining chondrogenic potential. When combined with hypoxia, soluble factors, and other environmental regulators, the resultant complex microenvironment will more closely resemble the in vivo niche, which further improves the cell capacity, even after extensive passaging. In this review, the adaptive mechanisms NP cells utilize in vivo are considered for insight into what factors are important for constructing a tissue-specific in vitro niche. Evidence for the use of SDSCs for NP regeneration is also discussed. Many aspects of NP behavior are still unknown, which could lead to future work yielding key information on producing sufficient numbers of a high-quality NP-specific population that is able to regenerate deteriorated NP in vivo.
Introduction
The nucleus pulposus (NP), found at the center of the IVD, is a tissue characterized by high proteoglycan (PG) and water content within a loose collagen network. It has a sparse, heterogeneous population of cells responsible for maintaining the matrix. The negatively charged extracellular matrix (ECM) retains between 66%–86% water [4], which permits distribution of and resistance to spinal loads, while sustaining the disc height. During development, the immature NP arises from the embryonic notochord and remains populated with notochordal cells (NCs) [5]. These large, highly vacuolated, and metabolically active cells produce a high PG content matrix in addition to providing regulatory cues for surrounding cells [6 –8]. With age and degeneration, the NP undergoes extensive changes in terms of matrix composition and cell population. Initial signs of degeneration present around the first decade of life along with a decline in NCs [9], although it is still unclear whether the decline is due to the continued differentiation of NCs into chondrocyte-like NPs or due to the programmed death (apoptosis) of the resident cells with invasion by cells from the cartilage endplate or annulus fibrosus (AF) [10 –12]. Degeneration of the NP results in a transition from the gelatinous healthy tissue to a more fibrous structure that is unable to provide complex mobility to the spine. The consequent altered transmission of spinal loads can ultimately lead to disc failure.
Conservative treatments of acute and/or chronic low back pain, such as bed rest, anti-inflammatory medications, and physical therapy, have proved ineffectual. The most popular surgical option is still discectomy followed by fusion. Despite the many reports of good to excellent outcomes with this method, long-term adverse biomechanical consequences to the adjacent vertebrae are still a concern [13].
Regenerative strategies over the past decades have advanced significantly, but a number of hurdles remain before a clinical treatment can be widely accepted. Autologous transplantation techniques are among the earliest tissue engineering strategies and recently have demonstrated promise for long-term articular cartilage repair in humans [14]. Due to a number of similarities between the cartilage and NP tissue, the autologous paradigm has been extended to disc repair. After promising results in canine models [15], the Euro Disc randomized trial found that, 2 years after surgery, patients maintained an increased pain reduction and a decreased loss of tissue hydration [16]. However, the effectiveness of this strategy may be limited due to the reliance on a population of cells with a deteriorated capacity as a result of the degenerative process. Procurement of cells is also difficult, in part, due to the high incidence of apoptosis [17], which reduces an already sparse cell population. This issue is further complicated by the extensive passaging procedures required to obtain sufficient numbers of cells for in vivo implantation. When passaged in monolayer, NP cells (NPCs) are known to proliferate slowly [18] and undergo increasing dedifferentiation [19 –21]. Cells from the degenerated NP also become increasingly senescent, which makes them minimally responsive to a growth factor or cytokine stimulation; the cells display an increased catabolic metabolism characterized by the increased production of matrix degrading enzymes [22 –25].
To address the need for a supplemental or alternative population, mesenchymal stem cells (MSCs) remain a topic of great interest. Recent literature highlights the use of MSCs in IVD and cartilage tissue engineering [26]. MSCs are multipotent stem cells that can be isolated from adult tissue (bone marrow, adipose, muscle, dermis, synovial membrane, synovial fluid, etc.). They possess the capacity to differentiate into cells with connective tissue lineages, including bone, fat, cartilage, ligament, and muscle. More specifically, MSCs are capable of differentiation to NPCs [27 –31]. Recently, a clinical study in patients diagnosed with lumbar disc degeneration with intact AF was performed using bone marrow-derived MSCs (BMSCs), which reported a higher water content at 12 months and improvement of pain and disability, but no improvement in the disc height [32]. Adipose-derived MSCs (ASCs) were also reported to be effective in promoting disc regeneration in a canine disc injury model [33]. Nevertheless, choosing an appropriate stem cell and preventing dedifferentiation and senescence during monolayer culture are big challenges for the ultimate success of cell-based NP regeneration.
In vitro culture may be a valuable opportunity to boost the potential regenerative capacity of NPCs or adult stem cells. With the correct conditions, pretreatment can yield a cell population with a greater in vivo success [34 –36]; this finding is of particular importance in the IVD due to the low-nutrient, hypoxic environment in which cells must ultimately function. Research applying the decellularized extracellular matrix (DECM) in tissue engineering indicates that it is a promising strategy to support proliferation and differentiation of recellularized cells due to their encrypted intrinsic cues to regulate cell functions and guide new tissue regeneration [37]. Compared to the DECM that is derived from mature tissue which can provide a lineage-specific differentiation signal, our recent studies demonstrated that the decellularized stem cell matrix (DSCM) can provide an in vitro niche-like microenvironment rich in stemness in which adult stem cells and primary cells, such as NP cells and chondrocytes, are able to greatly expand, while their redifferentiation capacity is retained. In this review, we will focus on key mechanisms utilized by NPCs throughout life for survival in the disc microenvironment. Stem cells derived from the synovium (SDSCs) are discussed as a potential tissue-specific stem cell for NP regeneration; their DSCM provides a key component of an in vitro microenvironment for NPC rejuvenation (key points summarized in Fig. 1 and Table 1).
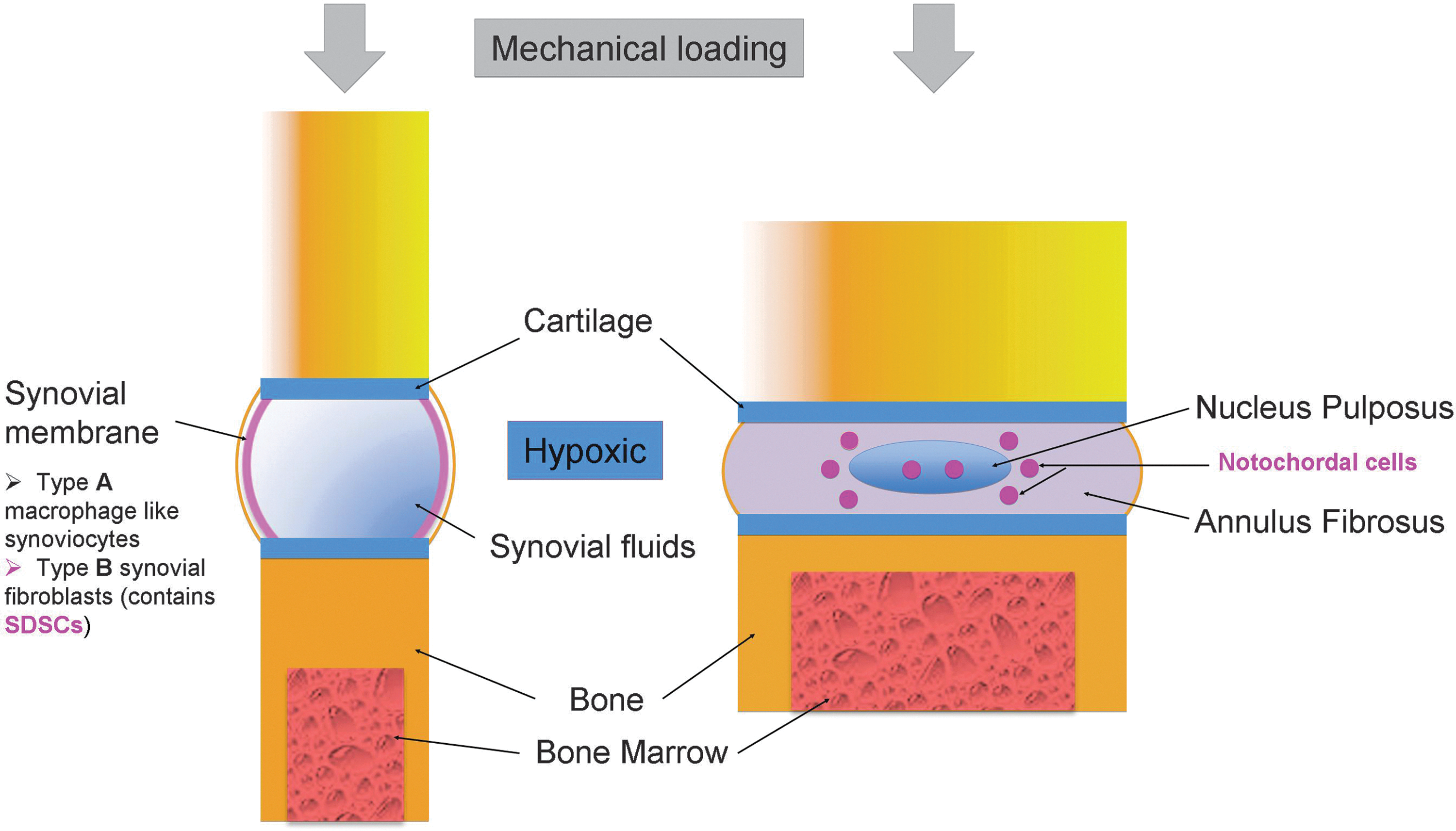
Resemblance between synovial joint and intervertebral disc. Color images available online at
ADAMTS, a disintegrin and metalloproteinase with thrombospondin motifs; DSCM, decellularized stem cell matrix; ECM, extracellular matrix; GAG, glycosaminoglycan; HAS2, hyaluronan syntheses 2; IVD, intervertebral disc; MMP, matrix metalloproteinases; NC, notochordal cells; NCCM, notochordal cell-conditioned medium; NP, nucleus pulposus; PG, proteoglycan; SDSC, synovium-derived stem cell; TGF-β, transforming growth factor-β; UDPGD, uridine diphosphoglucose dehydrogenase.
Disc Microenvironment
NCs and NPCs
During development, the mesoderm gives rise to the rod-shaped notochord, which in turn forms a juvenile NP [38]. The NC population of the disc gradually diminishes during the first years of life, until the majority of cells become mature, chondrocyte-like NPCs [5,39]. A low tolerance to environmental factors suggests that mature cells may arise due to the altered environment in the aging disc. As evidenced by greater rates of oxygen and glucose consumption along with more lactic acid production, NCs are more metabolically active than NPCs, which may explain their compromised ability to remain viable in the face of nutrient deprivation [40]. Additionally, mechanical loading in discs induces a large decrease in NC density compared to NPCs [41]. Therefore, NCs may transition to NPCs to retain viability in the face of increasingly harsh disc conditions.
Despite the increased hostility of the disc environment with age, some NPCs continue to express notochordal markers throughout life [42] and may play an important role for the cells. Coculture of NCs with NPCs results in a higher PG synthesis, a phenomenon thought to be a result of secreted soluble factors as opposed to cell–cell interactions [43]. This finding is supported by the recent identification of the connective tissue growth factor precursor in a NC-conditioned medium (NCCM), which induces an increase in ACAN and VCAN gene expression [44]. The increase in ACAN gene expression in response to the NCCM also extends to PG production by NPCs in a dose-dependent fashion and proliferation in a dose-independent fashion [7]. These results support the regulatory role of NCs through soluble factors. When the impact of varying ratios of NCs to NPCs was evaluated, the greatest increase in glycosaminoglycan (GAG) production occurred at a 50–50 ratio [45]. This information is particularly important given the widespread use of the stem cell–NPC coculture, as it provides an indication of the proportion of each cell type needed. While the precise mechanisms of an interaction are still in need of further investigation, it seems reasonable to conclude that, during the first years of life, NCs are present in sufficient numbers to continuously regulate the NPC behavior. Degeneration may, in part, represent a decline in NC regulation of the surrounding cells. Their loss is often implicated as the trigger for degeneration [9], but it might be beneficial to specify that NCs transition into a more mature population that is better able to survive under the changing disc conditions that occur with age. Although some NC-like cells remain, they are not present in sufficient numbers to provide ample regulatory cues for the more mature cells.
Metabolic adaptations of NPCs
The NP is avascular [46] and relies on diffusion over long distances for nutrient transport [47]. The most utilized vasculature originates in the vertebral bodies and extends across the superficial regions of the endplates [48]. Additionally, sparsely distributed vascular beds extend to the dorsal and ventral surfaces of the AF. Even after a major disc disruption, these vessels never penetrate the NP [49,50]. The unsurprising consequence is that oxygen concentrations are lowest and lactate concentrations are highest in the NP [51], further resulting in the accumulation of anaerobic metabolism byproducts that produce a low pH. The constantly variable loads on a negatively charged ECM create high osmolarity [52] that influences gene expression [53]. Therefore, the NP can be considered to exist in a hypoxic niche where cells express a unique phenotype that differentiates them from those of surrounding tissues [54].
The effects of hypoxia are regulated by the hypoxia inducible factor (HIF) group of transcription factors, whose family of HIF target genes regulates key cellular functions, including energy metabolism, angiogenesis, cell survival, apoptosis, matrix synthesis, proliferation, self-renewal, and differentiation [55]. HIF-1 is a heterodimer consisting of an α and a β subunit, with the former degrading quickly under normoxic conditions [56]. The combined action of HIF-1 and HIF-2 is considered to be dependent on the cell type [57]. A central function of HIF-1α is promoting transcription of glycolytic enzymes essential for cells to switch from aerobic to anaerobic metabolism [58], in part, by inhibiting adenosine-5′-triphosphate (ATP) production through oxidative phosphorylation [59]. One unique trait of NPCs is the continued expression of HIF-1α under normoxic conditions, whereas, in other cell types, this isoform is quickly degraded [60]. Known to be involved in controlling mineralization in various tissues, expression of ANK, a pyrophosphate transporter, is negatively regulated by both HIF-1 and HIF-2 in the NP and is thought to prevent mineralization [61]. A decrease in ACAN expression is also observed upon HIF-1 inhibition, suggesting that the unique oxygen-independent stabilization of HIF-1α is an important metabolic adaptation for both glycolysis and ACAN expression [62]. This finding further supports the recent report that NPC metabolism is not reduced at oxygen concentrations as low as 1% and that the hypoxic environment positively impacts aggrecan production, while promoting a healthy gelatinous matrix as opposed to a fibrous one [63].
Additionally, HIF-2 was also found to regulate the p300-binding protein cited2, which regulates both HIF-1- and HIF-2-dependent vascular endothelial growth factor (VEGF) expression in NPCs [64]. Another key consequence of the low-oxygen environment in NPCs is promoting the expression of proteins responsible for Notch signaling, an important regulator of cell proliferation [65]. Hypoxic activation of the phosphatidylinositol 3-kinase/Akt (PI3K/Akt) and the mitogen-activated protein kinase/extracellular signal-regulated kinase (MAPK/ERK) pathways is thought to underlie the downregulation of apoptosis in healthy NPCs in a hypoxic, serum-deprived environment [66]. Hypoxia also induces Notch signaling related to maintaining an undifferentiated state and cell fate determination [67,68].
Severe osmotic fluctuations impact the phenotype of resident cells. One of the most readily visible characteristics of NCs is their large vacuole, which plays an important role for regulating cell volume and tonicity [69]. The transmembrane channel protein Aquaporin 1 (AQP-1), which is permeable to water and O2, is expressed in a number of tissues and intact discs. It prevents rapid volume deformation under osmotic stress and facilitates O2 diffusion across the plasma membrane [70,71]. Expression of AQP-1 has also been shown to play a role in regulating chondrocyte volume as well as synovial homeostasis [72,73]. Hypoosmotic conditions downregulate AQP-1 to avoid rapid swelling deformation, although low oxygen upregulates AQP-1 to aid in oxygen use [74]. Hyperosmotic conditions activate the G1 and G2 cell cycle checkpoints, resulting in decreased proliferation for NPCs [75]. The negative effect on proliferation is maintained even in the presence of the platelet-derived growth factor (PDGF) or insulin-like growth factor-I (IGF-I) as evidenced by the reduced novel DNA synthesis and reduced ERK and Akt activation [76]. These results indicate that hyperosmotic conditions during culture are extremely undesirable, while a lower environmental osmotic pressure may be more favorable for in vitro culture where proliferation is the primary goal.
Integrin-mediated interactions between NPCs and ECM
Both in vitro and in vivo, the ECM to which cells are attached is a key microenvironmental factor that helps determine many aspects of cell behavior through both physical and biochemical interactions. It is a dynamic structure that remains active throughout life, facilitating the transmission of physiological signals through its mechanical structure and chemical composition [77]. A primary method of cell–ECM communications occurs via integrins, a family of transmembrane proteins operating as cell surface receptors that connect the cytoskeleton to the surrounding ECM [78]. Functioning as heterodimers, integrins are composed of an α and a β subunit. Each unique subunit combination determines binding to specific ECM components [79]. Importantly, integrin expression is dependent on the cell type and can be used to help identify downstream cellular responses from cell–matrix or cell–cell interactions.
Work by Nettles et al. quantitatively identified specific integrin subunits in NP and AF tissues using immunostaining. They found that, although the NP and AF both expressed integrin subunits associated with collagen, fibronectin, and laminin binding, the immature porcine NP contains a higher expression of the α6 and β4 subunits [80]. The α6 subunit, which occurs in both the α6a and α6b forms, also functions in chondrogenic differentiation. Specifically, the α6b subunit supports differentiation, while α6a is involved in stabilizing the chondrogenic phenotype [81]. Using flow cytometry to quantify integrin expression levels upon isolation and subsequently after monolayer culture, it was found that expression of the integrin subunit α6 was significantly higher in NPCs, while expression of the integrin subunit α5 was higher in AF cells [82]. It was also found that type II collagen adhesion occurred through the α1β1 receptor, but type I collagen adhesion was only partially mediated through this receptor. This finding suggests that type I collagen attachment is mediated by integrin receptors as well. In addition, fibronectin attachment was found to occur through the α5β1 integrin [82].
In combination with integrin subunits β1 or β4, the α6 subunit is a primary mediator of cellular adhesion to the ECM protein laminin, a heterotrimeric protein composed of individual α, β, and γ chains. Currently, at least 15 isoforms of laminin have been identified; these are expressed differently in various tissues [83,84]. In NPCs, a higher expression of the laminin α5 chain, laminin receptor integrin subunits α3, α6, and β4 along with CD239 and CD151 were found compared to cells from the AF [85]. Gilchrist et al. also examined the cell-matrix interactions and found a faster cell spreading and a higher resistance to detachment on laminin isoforms LM-511 and LM-332, in contrast to LM-111, fibronectin, and collagen II [86]. Substrates coated with laminin softer than 720 Pa were also found to promote in vitro PG production [87]. Unsurprisingly, the notochord from which the NP is believed to derive is a laminin-rich structure [88].
Taken together, these works provide insight into which specific receptors mediate NPC individual ECM components. One future application of these results might be to investigate their involvement in cell–DSCM interactions (discussed below), since the components in the above work account for a majority of DSCM composition.
Mechanical loading on NPCs
The IVD provides mobility to the spine and allows complex motion. However, mechanical loading of the IVD also contributes to disc degeneration. The IVD experiences various kinds of mechanical stimuli during loading, including volumetric changes, fluid flows, pressures, and deformations in tension, compression, or shearing configurations. The healthy IVD transfers load via hydrostatic pressurization of the hydrated NP. The NP contributes by dissipating energy under large physiological deformations [89]. Reductions in hydrostatic pressurization were observed in the aging and degenerated disc. Changes in tissue properties of the disc, including dehydration and reorganization of the NP and stiffening of the AF, markedly alter the mechanics of load transfer in the spine.
Biochemical changes occurring in early degeneration, including a reduced GAG content in the NP, may alter the disc mechanical behavior and contribute to the progression of degeneration [90]. Other than hydrostatic pressure, the NP also experiences high interstitial fluid and osmotic pressure during loading that arises from a high fluid content and PG-associated negative charges [91,92]. NPCs are also sensitive to fluid shear stress [93]. The magnitude- and duration-dependent changes in gene expression resulting from compression were identified in the NPC. Mechanical loading also influenced PRTN and ACAN expression by human IVD cells [94].
Mechanical loading affects matrix biosynthesis of IVD cells; the pathways are still unknown, although recent studies suggest that cellular matrix biosynthesis is an energy demanding process. The investigation of the effects of static and dynamic compressive loading on energy metabolism suggested that IVD cell energy production is affected in a cell type- and compression type-dependent manner; the total ATP content in the NP increased when 1 Hz dynamic loading was applied [95]. A trend of decreasing metabolic activity in the NP with increasing torsion magnitude was observed and the apoptotic activity in the NP significantly increased with 5° torsion [96]. Increasing magnitudes showed more anticatabolic gene expression changes, whereas increasing duration resulted in increases in procatabolic gene expression [97]. Strong differences in the disc response to cyclic torsion and compression are apparent with torsion increasing elastin expression and compression resulting in a more substantial increase in disc metabolism in the NP [98].
Construction of an In Vitro Microenvironment for NP Regeneration
Aberrant behavior of the cell population is a major component of the degenerative process, which gives various tissue engineering strategies a unique, specific target for intervention through stem cells. However, adult stem cells are not all created equal with respect to their capacity to generate different types of tissues. Even under chondrogenic induction, BMSCs tend to proceed to endochondral bone formation [99], while ASCs suffer from deficient chondrogenesis [100]. Consequently, the choice of an adult stem cell population will have a direct influence on clinical results. Given the precursor nature of NCs and their continued regulatory role of NPCs in the disc, it would be beneficial to find a population to serve as a functional replacement. Articular cartilage in synovial joints bears the most resemblance to the NP (Fig. 1), but is not a viable option due to a lack of precursors, high susceptibility to degeneration, and various other pathologies. In contrast, synovial lining has a highly robust adult stem cell population that exhibits a great capacity for both cartilage and NP repair and may be suitable for NP regeneration (Table 1).
Shared properties of SDSCs and NCs
Although currently classified as a symphysis, there is a debate as to whether the IVD should instead be classified as a diarthrosis. Arguments against reclassification to a diarthrodial joint were by Gringon and Roland who firmly concluded that the 2 joints are not categorically similar [101]. However, due to a number of morphological and functional similarities along with the new research, Shapiro et al. recently replied with an interesting (and uniquely titled) review of the new evidence for reclassification [102].
As discussed by Shapiro et al. [102], the majority of anatomical structures in the IVD and synovial joint can be matched relatively well in terms of structural similarities, with the exception of the NP. Although the synovial joint cavities are filled with synovial fluid, the fluid may serve some similar roles as the NP with respect to lubricating the adjacent cartilage. Lubricin was first isolated from the synovial fluid secreted by synovial cells [103] and primarily functions as a boundary lubricant for cartilage at the superficial zone [104]. Within the disc, lubricin is similarly found at the matrix of the superficial zone and additionally occurs throughout the disc [105], with a significant occurrence in the NP [106].
A significant amount of the large aggregating PG aggrecan is the most characteristic feature of the NP and has important functions in regulating cell behavior. The G1 domain of individual aggrecan molecules interacts with the link protein and hyaluronan to form stable ternary complexes. The final PG macromolecule aggregate contains many aggrecan molecules [107,108], with hyaluronan being essential for the formation of these large aggregates [109]. Both the link protein and hyaluronan can help regulate disc cell metabolism. The in vivo administration of the link protein in rabbit discs was found to increase ACAN gene expression, while decreasing PRTN gene expression [110]. Synoviocytes express link proteins [111] that may aid in the aggregation of fragmented PG in the disc and additionally produce their own GAGs [112].
The soluble factors secreted by NCs that increase NPC production of PGs [43] and increase proliferation [7] may occur, in part, by hyaluronan-related mechanisms. In articular chondrocytes, the addition of hyaluronan enhances PG synthesis despite the presence of interleukin-1 (IL-1) [113]. Similarly, NCs promote PG production, in part, by regulating hyaluronan levels; the NCCM induces an upregulation of hyaluronan syntheses 2 (HAS2) in NPCs [44]. Hyaluronan is synthesized through the membrane-bound HAS enzyme [114] and is critically important for both articular chondrocyte and synovial cell synthesis of hyaluronan. In the disc, it is crucial for normal development as evidenced by the malformation of IVDs in HAS2 knockout mice [115]. The synovium similarly expresses HAS in healthy subjects, but expression is decreased in arthritic patients [116]; this phenomenon may be comparable to the change in degenerated discs. A key receptor mediating cell interactions with hyaluronan is CD44, which is expressed in both the NP in vivo [117] and isolated NPCs in culture [118]. During development, CD44 colocalized with hyaluronan and PGs can be identified in rat NCs only upon disc formation [119]. Expression of CD44 is also found in 80%–90% of SDSCs [120,121]. The extensive role of CD44 and hyaluronan in the formation of large aggregating PGs characteristic of a healthy NP may make SDSCs a good choice.
The enzyme uridine diphosphoglucose dehydrogenase (UDPGD) is exclusively responsible for the conversion of uridine diphosphoglucose (UDP) to UDP-glucuronic acid [122]. In turn, UDP-glucuronic acid is essential for the synthesis of a number of GAGs, including hyaluronan, chondroitin sulfate, and heparin sulfate [123]. The protein core in aggrecan contains a domain for binding hyaluronan that in turn contains as many as 100 chondroitin sulfate chains and 60–80 keratan sulfate chains [124,125]. Therefore, it is reasonable to suggest that UDPGD is essential for proper tissue development and function. In contrast to numerous studies on the role of UDPGD in cartilage and synovium, to our knowledge, only one such study exists for IVD. Butler and Pousty found a large percentage of NCs in 5-month-old pigs contained UDPGD [126]. Type B synovial cells similarly demonstrate a high UDPGD activity [127,128], which may reflect their high hyaluronan production. In immobilized joints, a decreased hyaluronan content in the synovial fluid is associated with a decreased UDPGD activity in synovial cells [129]. The use of SDSCs with a high UDPGD activity may therefore aid hyaluronan restoration in degenerated discs.
Hypoxia and HIF proteins regulate expression of β-1,3,-glucuronyltransferase 1 (GlcAT-1), a key enzyme in GAG synthesis in NPCs [130]. The bone morphogenetic protein 2 (BMP-2) and transforming growth factor (TGF)-β stimulate expression of GlcAT-1 in NPCs through a signaling network comprising MAPK, AP1, TonEBP, and Sp1 [131]. GlcAT-1 promoted the synthesis of chondroitin sulfate GAG chains of the cartilage matrix. GlcAT-1 delivered by nonviral gene transfer was able to enhance PG synthesis and overcome cartilage PG depletion produced by IL-1β [132].
Advantages of SDSCs in NP regeneration
Synovial membrane contains 2 kinds of synovial cells: type A macrophage-like synoviocytes and type B fibroblast-like synoviocytes. The latter contains a population of adult stem cells, referred to as SDSCs [133]; these cells display a high chondrogenic potential (for a review of SDSCs as a tissue-specific stem cell for cartilage engineering and regeneration, see Jones and Pei [134]). Sufficient numbers of synovial cells can be obtained through a minimally invasive punch biopsy [121] and the regenerative capacity of the synovium has been demonstrated after analogous procedures [135,136], making isolation a low-risk process. Theoretically, the NP can be considered an immature type of cartilage, and the high chondrogenic potential of SDSCs combined with a lower hypertrophic potential make this population a good candidate for NP regeneration. Another interesting trait of SDSCs is that they may be able to mimic the regulatory and precursor roles typical of NCs. Not only do they support matrix production, but they also support the restoration of balance between anabolic and catabolic metabolism in the degenerated disc environment.
Our study first established the feasibility of using SDSCs in coculture with NPCs [137]. Compared to NPCs alone, there were high levels of collagen II and aggrecan with minimal levels of collagen I and no collagen X detectable in the coculture of SDSCs and NPCs [137]. This study indicated that SDSCs can beneficially regulate NPC behavior in a fashion reminiscent of NCs by stimulating the production of appropriate ECM. When SDSCs were implanted in vivo in rabbits, Miyamoto et al. also found that these cells could help restore homeostasis in disc cell metabolism through downregulating IL and matrix metalloproteinases (MMP)-related genes, while upregulating collagen- and PG-related genes [138]. These actions mimic the regulatory behavior of NCs in that when NPCs are treated with the NCCM, downregulation of IL-1β as well as MMP and a disintegrin and metalloproteinase with thrombospondin motif (ADAMTS) genes occurs [139].
Proinflammatory cytokines, such as IL-1β, and matrix-degrading enzymes, including MMPs and ADAMTS, are extensively involved in degeneration. They must be addressed for successful, lasting tissue regeneration. Compared to nondegenerated discs, degenerated discs demonstrate an upregulation of matrix-degrading enzymes, including MMPs 1, 3, and 13 as well as ADAMTS-4. While there is an accompanying increase in some of their inhibitors [140], there is a net increase in matrix-degrading enzymes over their inhibitors [141]. In addition to the increased gene expression of various matrix-degrading molecules, there is a decrease in key matrix genes, such as ACAN, COL2A1, and SOX6 [142]. Additionally, the trend of both mRNA and protein expression of key matrix components, such as aggrecan, types I and II collagen, and Sox9 increasingly decrease with the progression of degeneration, while mRNA levels of IL-1β and the tumor necrosis factor (TNF) alpha increase [143]. This increasingly catabolic state was found to be associated with a senescent phenotype [144].
The accumulation of fragmented PGs signifies a degrading disc matrix [145]. Fibronectin, which provides cues to cells via integrins and aids in ECM organization, becomes elevated in degenerated discs and also in fragments [146]. The smaller size of PG fragments allows them to slowly escape the disc over time, resulting in decreased disc water content. Alternatively, the injection of fibronectin fragments in healthy joints induces a similar degenerative process to traditional disc degeneration [147]. One potential tool in addressing the negative changes associated with the accumulation of fragmented matrix components is through hyaluronan. In cartilage explants, it can mitigate the effects of the fibronectin fragment-induced damage, in part, by enhancing PG synthesis [148]. It additionally acts as a suppressor of MMPs and ADAMTS (for a review of the anti-inflammatory actions of hyaluronan, see Masuko et al. [149]). Because UDPGD is necessary for the production of hyaluronan, the high activity of UDPGD in SDSCs may suggest one mechanism through which SDSCs help normalize damaged discs. When implanted in vivo, SDSCs appear to restore tissue homeostasis through the downregulation of IL-, TNF-, and MMP-related genes as well as upregulation of collagen- and PG-related genes [138]. These effects are in agreement with the known actions of hyaluronan and may, in part, be the result of a rise in hyaluronan production. Cell-based regenerative strategies are contingent on the use of a healthy, robust population of cells in sufficient numbers to overcome the negative changes in the disc environment. However, as discussed in the review by Li and Pei [24], stem cells are susceptible to senescent changes. To overcome this major hurdle, our group has demonstrated the ability of SDSCs to produce the DSCM that can revitalize cells during expansion.
A unique niche from DSCM deposited by SDSCs
For NP regenerative techniques, the current evidence indicates that SDSCs represent an important supplemental source of cells due to their ability to restore a more normal disc microenvironment, in part, increasing anabolic metabolism and decreasing catabolic metabolism. These actions may make them a good replacement for the diminished NC population. However, ex vivo expansion offers a chance to modulate cellular behavior and must be addressed for optimal outcomes. Intriguingly, our work has shown that SDSCs are capable of producing a unique ECM that can be decellularized to produce the DSCM as part of a tissue-specific microenvironment. Utilizing the DSCM during both SDSC and NP expansion greatly enhances their properties during in vitro tests compared to untreated cells. This result suggests that in vivo outcomes may similarly be improved upon, although it is yet to be confirmed.
The in vitro culture substrate allows for significant manipulation of cellular characteristics that enhance function. Plastic substrates, the current culture standard, impose negative changes on cells. Our work first identified the DSCM deposited by SDSCs as a promising tissue-specific substrate that dramatically improves proliferation and additionally enhances chondrogenic potential [150] (reviewed by Pei et al. [151]). The specificity for chondrogenic potential led us to investigate this type of matrix on chondrocyte expansion. We found that the DSCM could delay replicatively senescent articular chondrocyte dedifferentiation, while enhancing redifferentiation [152]. This finding is crucially important for cartilage tissue engineering and autologous techniques because of the extensive in vitro expansion required and the decline of the chondrocyte population with disease progression; these changes occur similarly in the NP. Chondrocytes were also capable of producing their own matrix that supported proliferation and enhanced chondrogenic potential [153]. Accelerated cellular senescence is prevalent in the NP during degeneration. In addition to increased catabolic metabolism, it results in decreased replicative potential [144]. Consequently, we investigated the effect of the DSCM on NPCs and found similar positive results. When expanded on plastic flasks, NPCs lost their redifferentiation capacity after 4 passages, while those expanded on the DSCM maintained a high proliferation rate as measured by cell numbers and high redifferentiation capacity for up to 6 passages as suggested by a higher ACAN, COL2A1, and SOX9, and a lower COL1A1 expression [154]. More importantly, DSCM-treated NPCs at passage 6 exhibited 10.8-fold higher COL2A1 and 10-fold higher ACAN mRNA levels compared with those expanded on plastic flasks, indicating that the DSCM expansion could retain NPC redifferentiation capacity without changing the ratio of ACAN to COL2A1 [154].
The regulation of cell behavior by the DSCM occurs, in part, through integrin-mediated interactions. When BMSCs were expanded on the DSCM, they displayed an increased rate of proliferation and strong chondrogenic potential compared to those cultured on plastic flasks. Assessments of integrin expression on DSCM-expanded BMSCs found upregulation of integrins α2 and β5 as well as sustained ERK1/2 and cyclin D1 activation [99]. When integrin α2 is inhibited, both focal adhesion kinase activation and ERK1/2 phosphorylation are suppressed [155], suggesting an underlying mechanism for DSCM regulation of cell behavior. In the future, comprehensive evaluation of integrin expression after DSCM expansion will likely yield important information regarding the specific mechanisms through which cell behavior is regulated by the DSCM.
In our most recent investigation, we attempted to manipulate matrix properties by permitting SDSCs to secrete the DSCM in hypoxic conditions and, furthermore, to determine whether NPCs alone or NPCs with SDSCs could produce an analogous matrix [156]. We found that matrix deposited by either SDSCs alone, NPCs alone, or a coculture of SDSCs and NPCs could provide a robust cell expansion system for SDSC proliferation. However, expansion on the SDSC-derived DSCM provided the highest mitotic effect. The hypoxia-prepared DSCM also favored NPC viability and GAG production as well as COL2A1 mRNA expression and decreased NPC hypertrophy, as evidenced by lower mRNA levels of COL10A1 and ALP. These results also present the intriguing possibility that DSCM properties can be further customized by treating cells (during ECM production) with exogenous factors.
The harsh environment in degenerated tissue, including inflammatory cytokines, MMPs, nitric oxide, serum deprivation, or mechanical forces, might cause implanted cell necrosis or apoptosis [141 –143]. Oxidative stress has been demonstrated to play a role in disc degeneration and discogenic back pain development by an increased synthesis of proinflammatory cytokines [157]. Our recent reports suggested that DSCM-expanded SDSCs could acquire an enhanced capacity to resist reactive oxygen species-mediated apoptosis [158]; the implantation with DSCM-expanded SDSCs exhibited hyaline cartilage resurfacing, indicating that DSCM pretreatment could not only enhance chondrogenic capacity of SDSCs, but could also probably promote the ability to resist inflammation [159].
Development of a comprehensive microenvironment for in vitro NPC expansion
Two of the primary hurdles for cell-based disc regeneration are limitations in the cell population and inadequate ex vivo expansion procedures. The native cell population increasingly declines, while the disc microenvironment becomes increasingly hostile. Available evidence suggests that the first hurdle may be overcome through the use of SDSCs for their ability to restore homeostasis in the disc, while supporting ECM reconstruction. The second major hurdle may also be addressed indirectly with SDSCs; the DSCM secreted by SDSCs provides a highly promising substrate for NP regeneration. To our knowledge, the SDSC-produced DSCM is the most beneficial substrate for not only NPCs, but for other SDSCs as well. Additionally, environmental hypoxia can support in vitro expansion and modulate DSCM properties.
Growth factor supplementation during culture expansion can support cell proliferation and help retain differentiation potential. Containing a connective tissue growth factor precursor [44], the medium conditioned by NCs stimulates DNA synthesis through the MEK/ERK and PI3k/Akt pathways [160]. The positive effects on human IVD cell proliferation appear through similar pathways after treatment with the PDGF, basic fibroblast growth factor (FGF-2), or IGF-I [160,161]. Another study suggested that the FGF-2 was found to help NPCs maintain a differentiated phenotype through multiple passages [162]. Although not comprehensive, other reviews have discussed the topic in detail and evidence is presented to demonstrate the importance of certain exogenous factors as part of an in vitro microenvironment. Interestingly, hypoxia has been shown to trigger the PI3K/Akt and MAPK/ERK pathways as well [66,163].
The combination of DSCM, hypoxia, and biochemical factors provides an in vitro microenvironment that more closely mimics the in vivo niches in which stem cells reside. The added degrees of control are not redundant and more strongly benefit cells together rather than individually. A study examining the combination of hypoxia, FGF-2, and DSCM produced the highest cell numbers, downregulated key hypertrophic markers, and promoted a strong chondrogenic response upon TGF-β induction. Although FGF-2 supplementation alone increased the expression of hypertrophic marker genes, the combination with hypoxia or DSCM successfully decreased the expression of these genes [164]. These data indicate that the added degrees of control can be used to promote a number of desired cell behaviors, while minimizing undesirable ones through pretreatment in a more biologically relevant microenvironment. While such a comprehensive system represents significant progress from older expansion systems in terms of microenvironmental control of cell behavior and prevention of senescence, the complexity pales in comparison to the degree of control occurring in vivo. Additionally, a number of factors have yet to be investigated. In the future, improvement of autologous techniques will undoubtedly be largely contingent on understanding the various interactions between factors and mechanisms through which cells sense and interact with their environment. This information will facilitate the construction of complex ex vivo microenvironments able to yield cell populations suited to addressing unique patient needs.
Summary
In this review, we have focused on discussing the potential of applying the stem cell matrix deposited by SDSCs to expand NPCs for disc engineering and clinical applications. There are still challenges to success, such as the composition of the DSCM deposited by SDSCs and whether the DSCM deposited by SDSCs is better than that deposited by other stem cells. Any progress on the above unsolved issues will further our ultimate aim of constructing an in vitro microenvironment for healthy and functional NP regeneration. SDSCs are tissue-specific stem cells and currently available research suggests that they may mimic the regulatory role of NCs for NP regeneration. If successful, SDSCs could greatly supplement current autologous techniques by providing both greater cell numbers from a smaller initial population and reversing senescence.
Footnotes
Acknowledgments
The authors thank Suzanne Smith for her help in editing the manuscript. This project was partially supported by research grants from West Virginia University Senate Research grant award (R-12-010), the AO Foundation (S-12-19P), the Musculoskeletal Transplant Foundation, and NIH R03 (no. 5 R03 DE021433-02).
Author Disclosure Statement
No competing financial interests exist.