
Abstract
The human cornea contains stem cells that can be induced to express markers consistent with multipotency in cell culture; however, there have been no studies demonstrating that human corneal keratocytes are multipotent. The objective of this study is to examine the potential of human fetal keratocytes (HFKs) to differentiate into neural crest-derived tissues when challenged in an embryonic environment. HFKs were injected bilaterally into the cranial mesenchyme adjacent to the neural tube and the periocular mesenchyme in chick embryos at embryonic days 1.5 and 3, respectively. The injected keratocytes were detected by immunofluorescence using the human cell-specific marker, HuNu. HuNu-positive keratocytes injected along the neural crest pathway were localized adjacent to HNK-1-positive migratory host neural crest cells and in the cardiac cushion mesenchyme. The HuNu-positive cells transformed into neural crest derivatives such as smooth muscle in cranial blood vessels, stromal keratocytes, and corneal endothelium. However, they failed to form neurons despite their presence in the condensing trigeminal ganglion. These results show that HFKs retain the ability to differentiate into some neural crest-derived tissues. Their ability to respond to embryonic cues and generate corneal endothelium and stromal keratocytes provides a basis for understanding the feasibility of creating specialized cells for possible use in regenerative medicine.
Introduction
T
Previous studies have isolated subpopulations of human corneal epithelial and stromal multipotent stem cells by their ability to efflux Hoechst 33342, a fluorescent dye that leads to easy fluorescence-activated cell sorting [3 –5]. A study by Du et al. identified a side population (SP) of stem cells in corneal stroma located in the peripheral region posterior to the Palisades of Vogt [4]. These cells were rare in number, and after four passages in primary human corneal culture, the SP cells comprised 1.3% of the total population. In vitro, the SP cells expressed ABCG2, an ATP binding cassette transporter, which has been identified as a specific marker for stem cells in a number of adult tissues [6 –8]. Du et al. (2005) were able to demonstrate that cultured human stromal SP cells were capable of expressing keratocyte, cartilage, and glial cell-specific makers in response to high levels of various growth factors in vitro. They concluded that the adult human corneal clonal SP cells retain the potential for differentiation.
While these studies indicate that the human cornea contains stem cells that can be induced to express markers consistent with multipotency in cell culture, there have been no studies indicating that human stromal keratocytes are multipotent. Our laboratory has reported the ability of quail keratocytes isolated from late embryos to retain neural crest progenitor properties by differentiating into stromal keratocytes, corneal endothelium, and other neural crest-derived tissues [9]. This suggests that stromal keratocytes retain the multipotency of their neural crest precursors.
The objective of this study is to examine the potential of human fetal keratocytes (HFKs) to differentiate into neural crest-derived structures when challenged in an embryonic environment. Stromal keratocytes isolated from human fetal corneas were injected along cranial neural crest migratory pathways and in the periocular mesenchyme in chick embryos. In the chick embryonic microenvironment, the injected keratocytes retained multipotentiality and transformed into neural crest derivatives such as smooth muscle in cranial blood vessels, stromal keratocytes, and corneal endothelium. However, they failed to form neurons despite their localization in the condensing trigeminal ganglion. These results show that HFKs retain the ability to differentiate into other tissues. Their ability to respond to embryonic cues and generate corneal endothelium and stromal keratocytes provides a basis for understanding the feasibility of creating specialized cells for possible use in regenerative medicine to treat corneal wounds and defects.
Materials and Methods
Isolation of stromal keratocytes and culture in vitro
Human donor corneas (approximately 1 month old and not suitable for human transplantation) obtained from Tissue Banks International (
Preparation of isolated keratocytes for microinjection
The primary cultures were confluent after incubation of 8 days in KSFM media. After one passage, cells were removed from plates and separated into single cells by incubation in balanced salt solution containing 0.25% trypsin/1 mM EDTA at room temperature for 1–5 min. The enzymatic reaction was stopped by addition of a soybean-trypsin inhibitor. Cells were transferred to centrifuge tubes and spun down at 2000 rpm. The cell pellet was washed three times with large volumes of Ringer's solution containing 0.1% BSA, resuspended in 6–10 μL of the same solution, and then kept on ice until injected.
Microinjection of HFKs into chick embryos
Fertilized White Leghorn chick (Gallus gallus domesticus) eggs were obtained from commercial sources. Chick eggs were incubated at 38°C for 30 h to obtain 5- to 7-somite (HH stages 8–9) [13]. Embryos were prepared as previously described [14]. Briefly, chick eggs were windowed and the vitelline membrane removed from the region of injection using pulled glass needles. HFKs were injected under the ectoderm alongside both sides of the dorsal neural tube extending from the region of the caudal diencephalon to the hindbrain (Fig. 1). In some experiments HFKs were injected into the periocular region of E3 embryos. Injected embryos were re-incubated for 0–15 days, n=10, for each time point. At the time of injection, 2 μL of cells was resuspended into 1 mL of KSFM media and distributed evenly into 6-chamber slides. The cells were allowed to attach at 37°C under 5% CO2 for 4–6 h.

Human fetal keratocytes (HFKs) in culture and after injection along migratory pathways of chick neural crest cells.
Immunohistochemical analysis and imaging
Cell cultures were briefly rinsed with Ringer's solution then fixed in 4% paraformaldehyde (PFA) for 2 h at room temperature. Experimental chick embryos were collected in Ringer's solution, fixed over night in 4% PFA, and then embedded in gelatin and cryosectioned at 8–10 μm. Fixed cell cultures and cryosections were prepared for immunostaining using standard protocols. Briefly, samples were rinsed in phosphate buffered solution containing 0.1% triton-X (PBT) and blocked with antibody buffer (PBT containing 0.2% BSA and 5% heat-inactivated goat serum). To label corneal stromal cells, mouse chondroitin sulfate (IgM; Sigma) was used diluted 1:200 and rabbit CD34 (IgG; Santa Cruz Biotechnology) was used diluted 1:100. Human nuclear antibody (HuNu; Millipore) was used diluted 1:500 (IgG1) to label all human cells. HNK-1 (IgM) was used diluted 1:50 to label migratory neural crest cells. Mouse anti-MF20 (IgG2b, DHSB) against myosin heavy chain was used diluted 1:3 to label cardiac muscle, and smooth muscle actin-SMA (IgG2a; Sigma) was used diluted 1:1000 to label the smooth muscle lining of blood vessels. Mouse HuC/D (IgG2b; Invitrogen) was used diluted 1:500 to label neuronal cells, and rabbit anti-Neurofilament 200 (IgG; Sigma) was used diluted 1:100 to label axons. Rat anti-N-cadherin (IgG; DHSB) was used diluted 1:3 to label the corneal endothelial cell layer.
Following overnight (1 h at room temperature for cells) incubation in antibody solution, sections and cell cultures were extensively rinsed in PBT, blocked, and then incubated in secondary antibodies. The following secondary antibodies (Molecular Probes) were used at 1:200; Alexa 594 goat anti-mouse IgG1, Alexa 488 goat anti-mouse IgG1, Alexa 350 goat anti-mouse IgM, Alexa 488 goat anti-rat IgG, Alexa 488 goat anti-rabbit IgG, Alexa 488 goat anti-mouse IgG2a, and Alexa 350 goat anti-mouse IgG2b. Some sections were counterstained with DAPI to show all nuclei. Slides were cover-slipped with Perma Flour (Immunon) and fluorescent images were captured using a Zeiss Axiocam mounted on a Zeiss Axioskop 2 microscope (Zeiss). Images were assembled using Adobe Photoshop 7.0 (Adobe Systems, Inc.).
Results
Isolation and transplantation of HFKs
To determine the differentiation state of the HFKs at the time of injection, some cells were allowed to attach in vitro then examined for morphology and expression of keratocyte markers. Live attached cells exhibited a flat dendritic morphology with several long interconnecting cellular processes (Fig. 1A) that is typical for keratocytes [15]. Immunocytochemistry revealed that the cells stained positive for the transmembrane protein, CD34. CD34 is expressed in stromal keratocytes but not in corneal epithelium, endothelium, or human corneal stromal stem cells in vitro [16 –18]. CD34 expression is lost in keratocytes that have become myofibroblastic or fibroblastic in culture [16,19]. In addition, the cultured cells stained positively for chondroitin sulfate (Fig. 1C), which has been previously shown to label keratocytes [20]. All cells were positive for both CD34 and chondroitin sulfate (Fig. 1D). Furthermore, we found that all cells stained positive for stromal keratocyte-specific 3G5-defined ganglioside, but did not stain for N-cadherin, an endothelial marker (data not shown). Thus, at the time of injection into chick embryos, cultured HFKs expressed markers that are characteristic of keratocyte phenotype.
To elucidate their developmental potential, HFKs were injected bilaterally into the cranial mesenchyme adjacent to the neural tube of stage 9 (E1.5) chick embryos. In embryos fixed immediately after injection (0 h, n=4), aggregates of cells positively immunostained with the human specific nuclear antibody (HuNu) were detected on either side of the neural tube spanning from the rostral mesencephalon to the rhombomere 2 (Fig. 1E). Injected cells were in the region of the endogenous migratory neural crest cells, labeled by the HNK-1 antibody, but were not themselves positive for the neural crest marker (Fig. 1F, G).
Injected HFKs co-localize with migratory neural crest cells
In the cranial region of the developing chick embryo, neural crest cells exit the dorsal neural tube and migrate extensively along defined routes [21,22]. To determine whether the grafted human keratocytes follow the path of migratory neural crest cells, we examined the embryos at approximately 20 h following injection. Injected embryos collected at this time (n=7) showed bilateral dispersions of HuNu-positive cells from the injection sites adjacent to the neural tube (Fig. 1E) into the surrounding cranial region (Fig. 2A). Cross section through the diencephalic region revealed that HuNu-positive cells co-localized with HNK-1-positive cells that form the dorsal optic stream of migratory neural crest cells (Fig. 2B, arrowheads), but were absent from the ventral stream (Fig. 2B, arrows). At the midbrain level, HuNu-positive cells co-localized with the late-migrating neural crest cells (Fig. 2C, arrowheads) that subsequently aggregate to form the trigeminal ganglion. However, HuNu-positive cells were interspersed with the cranial mesenchyme (Fig. 2C, asterisks) and did not appear to co-localize with, but they were located adjacent to, the HNK-1-positive cells in the lateral regions. A similar pattern was observed in the hindbrain region, although HuNu-positive cells were intermixed with the cardiac mesenchyme (Fig. 2D). Our results show that HFKs injected adjacent to the neural tube either co-localize with the endogenous migratory neural crest cells or disperse in the cranial mesenchyme. Approximately 6×103 cells were introduced per embryo.
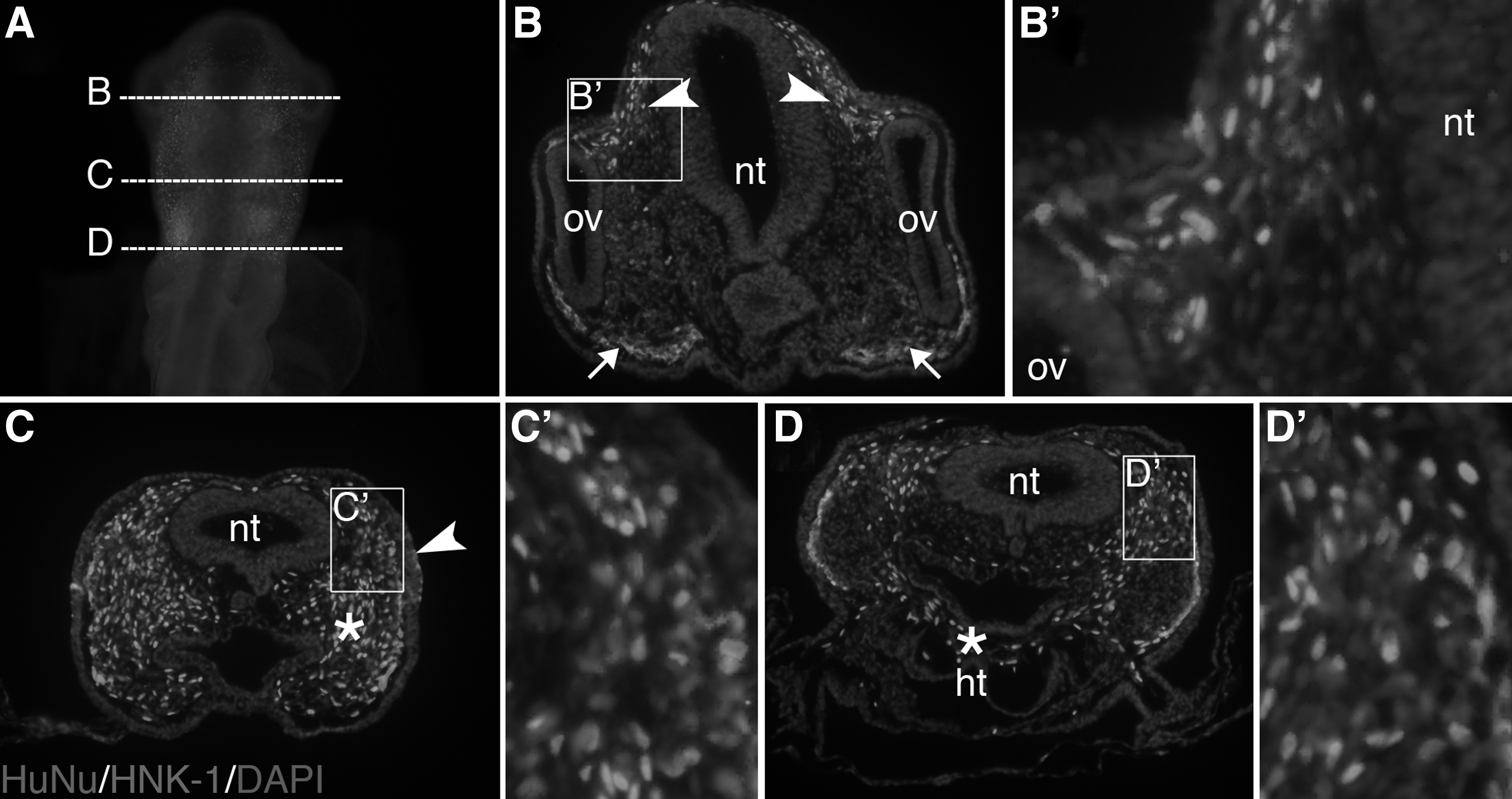
Injected keratocytes co-localize with migratory neural crest cells.
HFKs contribute to cranial blood vessels and heart
To determine whether grafted HFKs can acquire different fates due to signaling from the embryonic environment, we analyzed their expression of specific differentiation markers at later stages of development. Transverse sections through E5 injected embryos contained HuNu-positive cells lining the cranial blood vessels (Fig. 3A). As cranial blood vessels and pericytes of ocular vasculature are derived from the neural crest [21,23 –25], the HuNu-positive cells appeared to migrate to an appropriate neural crest location. They also expressed the tissue-specific marker, alpha-smooth muscle actin (α-SMA), consistent with that of the surrounding endogenous pericytes (Fig. 3B, arrows). In contrast, those HuNu-positive cells that localized in the mesenchyme adjacent to the neural tube did not stain for α-SMA (Fig. 3C), suggesting that expression in the blood vessels was specific. Similar to E4, HuNu-positive cells were found in the cranial blood vessel linings at E7 and demonstrated α-SMA expression (Fig. 3D, E).

Injected HFKs contribute to cranial blood vessels and cardiac cushion mesenchyme. Section through E5 head demonstrating HuNu-positive cells in the blood vessels
Another derivative of cranial neural crest cells is the cardiac cushion mesenchyme and the cardiac outflow tracts [26,27]. Similar to the endogenous neural crest cells, injected HFKs migrated through the pharyngeal arches and into the primitive heart region where they localized within the cardiac cushion mesenchyme between the endocardium and myocardium (Fig. 3F–H). This mesenchyme is surrounded by the early developing great vessels, which express α-SMA, and the myocardium wall, which is MF20 (sarcomeric myosin)-positive. Unlike the HuNu-positive cells in the cranial blood vessel lining, which were positive for α-SMA, the injected cells that migrated into the cardiac cushion mesenchyme did not express either α-SMA or MF20. Endogenous neural crest cells that localize in the cardiac cushion mesenchyme do not express α-SMA or MF20 [28] either; however, we were unable to specifically identify the differentiation state of these cells due to the lack of good markers for cardiac cushion cells. Nevertheless, it appears that the injected HFKs migrated into the cardiac cushion mesenchyme as did the endogenous neural crest cells. It is therefore possible that they respond to environmental cues in this region and contribute to cardiac development.
Injected HFKs localize in the trigeminal ganglion but they do not form neurons
Neural crest cells contribute to sensory and autonomic ganglia of the peripheral nervous system [29 –31]. We examined the ability of injected HFKs to contribute to the trigeminal ganglia. Since HuNu-positive cells localize with the presumptive trigeminal ganglion mesenchyme (Fig. 2C), we examined their ability to differentiate into sensory neurons. HuNu-positive cells were located in the proximal region of the trigeminal ganglion (Fig. 4A–D), where endogenous neural crest cells normally form sensory neurons and glia [31]. However, the HuNu-positive cells did not stain for the neuron-specific markers HuC/D and neurofilament at E5 and E8 (Fig. 4B, D). Our results indicate that despite the co-localization of injected HFKs with neural crest cells during gangliogenesis, they do not respond to the neurogenic environmental cues to form trigeminal sensory neurons.
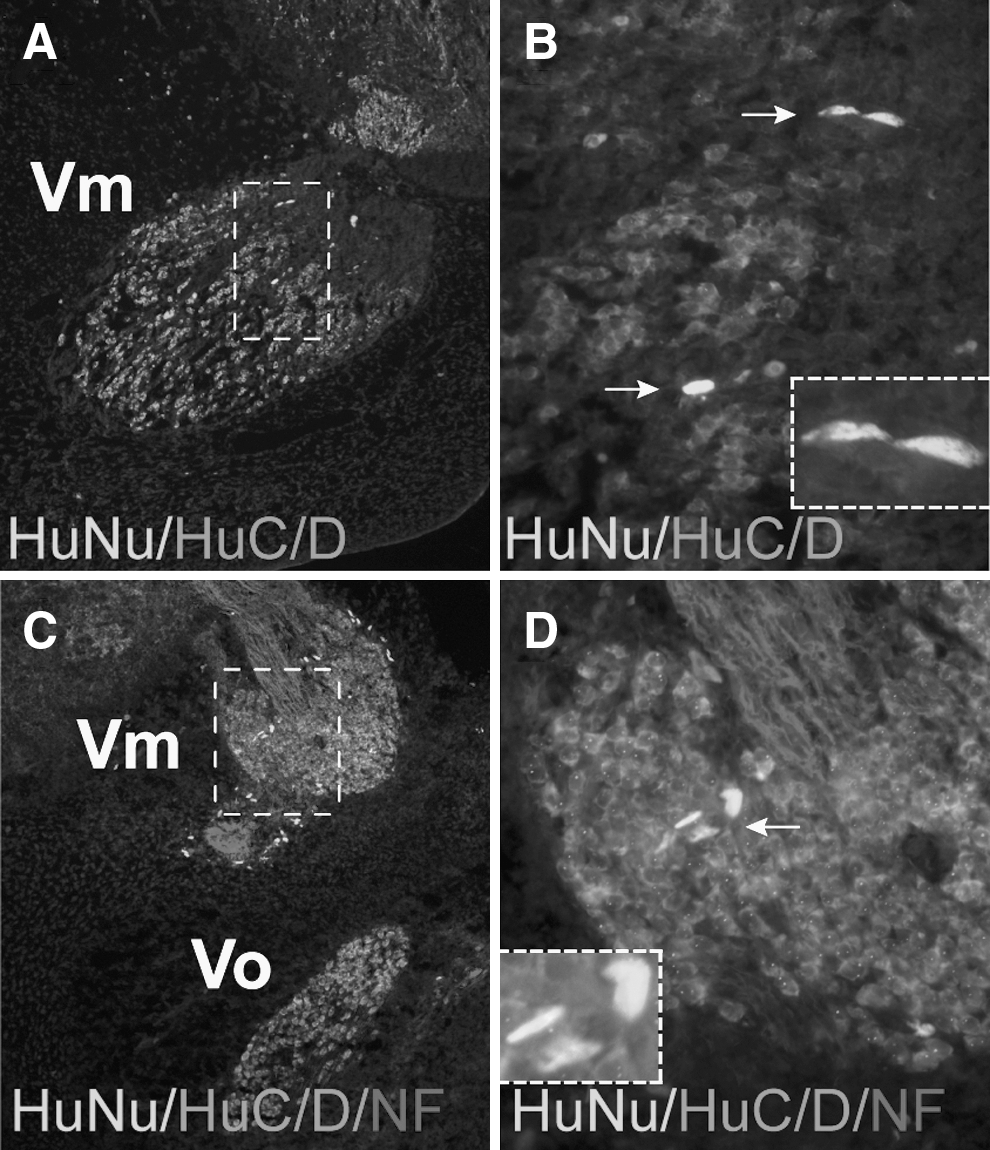
Injected HFKs localize within the developing chick trigeminal ganglion. E5
HFKs contribute to corneal development in the chick embryo
Endogenous neural crest cells that form the cornea pause for about 1 day in the periocular region at E3 before migration into the eye [32]. In chick, neural crest migration occurs in two waves, beginning at approximately E4 to form an endothelial monolayer of cells, and is followed by a second wave that invades the primary stroma and differentiates into keratocytes by E6. To determine whether injected HFKs contribute to the chick cornea, the eye region of host embryos was examined at developmental stages corresponding to the times of the neural crest invasion, from E3 through E8. While HuNu-positive keratocytes could be detected in the migratory pathway leading to the periocular region by E3, only very few cells were found in the periocular mesenchyme poised to form the cornea. By E5, no HuNu-positive cells contributed to forming corneal endothelium, but were instead found in the region of the perilimbal sclera (data not shown). From this observation, it is possible that the majority of the HuNu-positive cells did not co-localize with the neural crest of the presumptive cornea during eye development. Another possibility is that the HuNu-positive cells proliferated at a slower rate than the endogenous periocular mesenchyme.
To overcome this shortfall and to challenge HFKs in the embryonic environment of the forming cornea, cells were injected directly into the space between the lens vesicle and the overlaying presumptive cornea epithelium at E3 (Fig. 5A, arrow). By E5, HuNu-positive cells behaved like the endogenous neural crest cells and organized in a monolayer between the lens and ectoderm, characteristic of the corneal endothelium (Fig. 5E, arrows). In addition, these cells expressed both the stromal marker chondroitin sulfate and the endothelial marker N-cadherin (Fig. 5F, arrowheads), although the latter at a lower level than endogenous endothelial expression (Fig. 5F, asterisk). By stage E8, all three layers of the cornea are formed (Fig. 5G). Corneal stromal cells have changed from mesenchymal neural crest cells to exhibiting the thin and elongated morphology characteristic of keratocytes [33]. Corneal endothelial cells express N-cadherin, which distinguishes them from the other corneal layers [34]. In sections of E8 embryos, HuNu-positive cells were present in both the cornea stroma and endothelium. HuNu-positive cells in the corneal stroma were positive for chondroitin sulfate while those in the corneal endothelium remained N-cadherin-positive, suggesting that HFKs can respond to environmental cues within the forming eye and differentiate into appropriate cell types of the cornea.
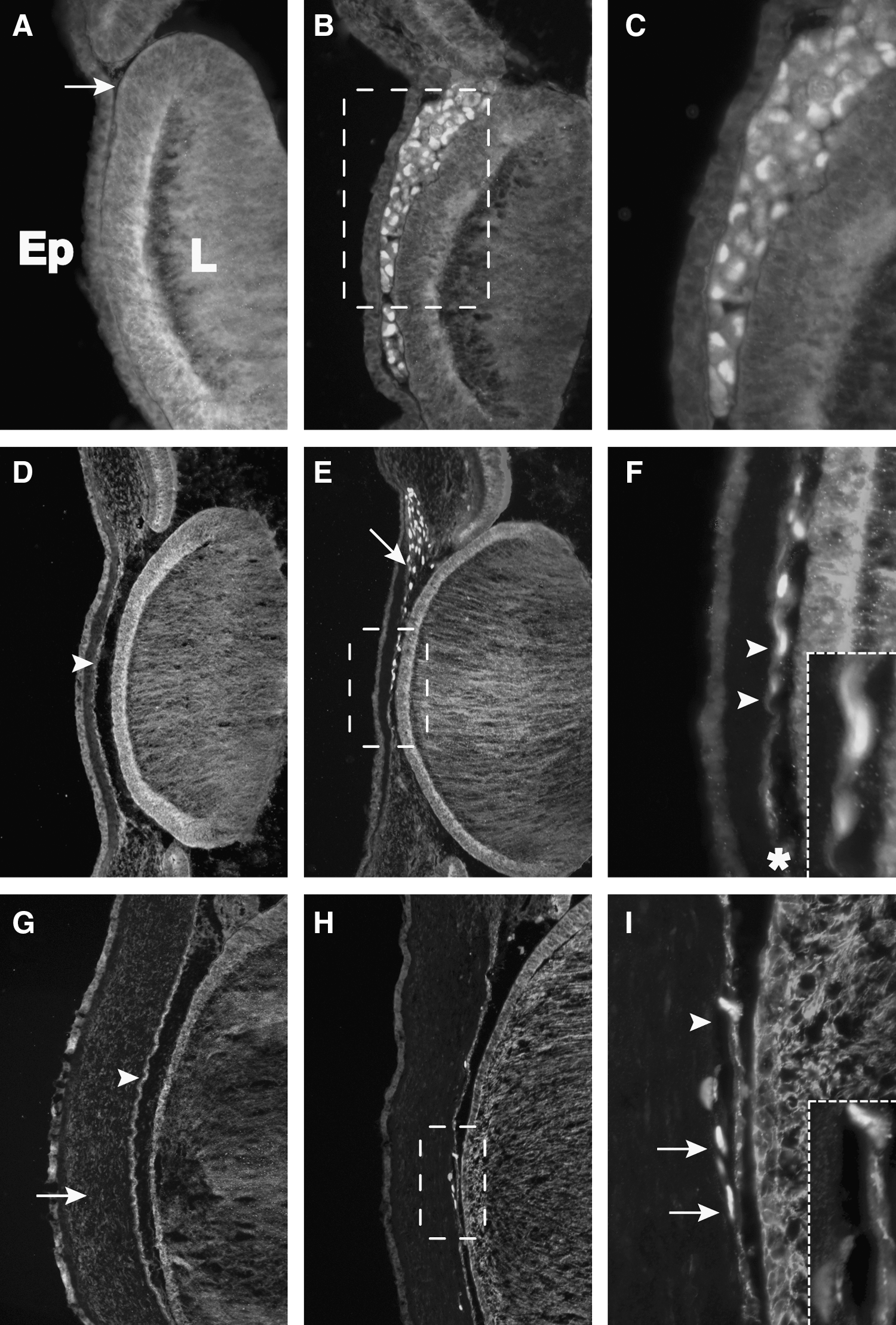
HFKs injected adjacent to the periocular mesenchyme form early corneal endothelium and stromal keratocytes. HFKs were injected between the lens rudiment and presumptive corneal epithelium at E3, shown by the arrow in the contralateral control eye
Discussion
Stromal keratocytes comprise the largest proportion of the cornea and secrete extracellular matrix proteins that are crucial to its transparency. Despite the important role of these cells in vision, most studies are focused on the ability of human keratocytes to maintain their phenotype in vitro. Thus, little is known about their regenerative potential. In this study, we show that when grafted into the embryonic environment, human stromal keratocytes respond to signals available to endogenous neural crest cell precursors and can differentiate into some neural crest-derived tissues, including cranial blood vessels, corneal endothelium, and keratocytes.
Previous studies have demonstrated that cells other than keratocytes reside in the mammalian corneal stroma, including corneal stromal stem cells [4,35,36]. These multipotent mesenchymal stromal cells are present in the peripheral and limbal cornea and have the ability to differentiate into tissues that are distinct from the ocular mesenchyme such as adipocytes, osteocytes, and chondrocytes [37 –39]. A recent study demonstrated that human cornea-derived precursor cells can be induced to form functional endothelial cells by retinoic acid and wnt/β-catenin signaling in vitro [40]. In addition, corneal stromal stem cells have been observed to adopt keratocytic function and characteristics when injected into mouse corneal stroma [41]. The HFKs characterized in our study likely represent a distinct population from these stem cell populations. They were isolated from the central cornea, excluding the peripheral and limbal regions where stem cells have been proposed to reside [4,35]. At the time of injection into the chick embryos, HFKs were positive for CD34 and chondroitin sulfate markers, which are expressed by keratocytes, but not corneal fibroblasts, myofibroblasts, or human corneal stromal stem cells in vitro [16 –18]. The combined expression of these makers and their elongated and dendritic morphologies suggest that HFKs were keratocytic at the time of injection into the embryos.
Following injection into the cranial region of the chick embryo, the HuNu-positive cells did not appear to strictly follow the pathways of the endogenous neural crest cells but rather dispersed randomly within the cranial mesenchyme [42,43]. This suggests that the injected keratocytes did not respond to the same migratory cues, such as semaphorins 3A and 3F, multiple ephrins, integrins, annexin A6, and BMPs, that guide neural crest cells to various locations [44 –50]. Nonetheless, some HuNu-positive cells co-localized with the neural crest cells as they ceased to migrate in the regions of the condensing cranial ganglia, in the ventral region of the forming cranial blood vessels and heart, and in the periocular region of the forming eye. Only the HuNu-positive cells that resided along the cranial blood vessels were SMA-positive, suggesting that they responded to pro-angiogenic growth factors such as TGFß, PDGF, and VEGF in the vascular environment associated with the formation of myofibroblasts [51 –54]. Despite their localization in the trigeminal ganglion, the HFKs did not form neurons. This result confirms our previous observation that quail embryonic keratocytes do not differentiate into neurons [9]. Furthermore, HuNu-positive cells only contributed to the corneal endothelium and stroma when they were directly injected into the periocular region. Given that the HFKs injected alongside the neural tube were localized in the periocular region, it is possible that the HFKs proliferated more slowly than the endogenous chick cells and became diluted during cornea development. Within the ocular region, HFKs differentiated into N-cadherin-positive corneal endothelial cells and also contributed to the stroma.
Due to the inability of human corneal keratocytes to maintain their characteristic phenotype in culture, several studies have identified potential sources for keratocyte differentiation. Human bone marrow mesenchymal stem cells and adipose-derived stem cells have been shown to differentiate into keratocyte-like cells in vitro [55,56]. Transplanted directly into the cornea stroma of mice, these bone marrow and umbilical stem cells assumed typical keratocytic phenotypes [56,57]. Our study demonstrates for the first time that the stromal keratocytes retain sufficient multipotentiality to form several derivatives in addition to keratocytes. They can display characteristics of corneal endothelial cells, vascular smooth muscle cells, and perhaps the endocardial cushion mesenchyme. Interestingly, it has recently been shown that human keratocytes can be reprogrammed into induced pluripotent stem cells, which, when injected with a hydrogel, may aid in corneal wound healing [58].
We utilized keratocytes isolated from 1-month-old corneas in this study. Upon migration into the region of the presumptive cornea, neural crest cells down-regulate HNK-1 [59], transform from mesenchymal to fibroblastic morphology [33], and initiate the expression of molecular markers such as collagens, keratan sulfate, and chondroitin sulfate [60 –63]. During subsequent development, the newly formed keratocytes synthesize the extracellular matrix and maintain the transparency of the corneal stroma. At birth, the human corneas are fully formed and transparent, and the isolated HFKs exhibited keratocyte characteristics that are different from the mesenchymal neural crest progenitors. However, it is possible that tissue age may play an important role in the multipotentiality of human keratocytes. After birth, the infant cornea continues to develop by changing the shape of its curvature, increasing in size, and inverting its horizontal and vertical dimensions [64,65]. Therefore, fetal keratocytes may possess the potential to proliferate, unlike the quiescent adult keratocytes. Keratocyte density is reduced with age as demonstrated by in vivo confocal microscopy, and this reduction appears to be associated with a decreased density of mitochondria in the remaining keratocytes as well as changes in the collagen fibrils seen with age [66 –69]. Because of the extensive corneal remodeling that occurs after birth and the continuous changes that occur with aging, it is possible that stromal keratocytes early in postnatal development demonstrate more plasticity to embryonic environmental cues than adult stromal keratocytes.
In conclusion, our results demonstrate that HFKs retain the ability to form nonocular neural crest derivatives and corneal endothelium, while also preserving their keratocytic phenotype upon incorporation into the corneal stroma. While the necessary embryonic environmental cues need to be elucidated, our results suggest that fetal human keratocytes may be capable of serving as sources for use in corneal regenerative cell therapy or tissue engineering.
Footnotes
Acknowledgments
This work was supported by the National Eye Institute EY022158 (P.Y.L.), the National Institute of Health DE16459 (M.E.B.), the National Eye Institute K08EY019714 (J.R.C.), and grants from Fight for Sight, the Knights Templar Eye Foundation, and an Unrestricted Grant from Research to Prevent Blindness (J.R.C.).
A portion of this work was previously presented in two abstracts:
1. JR Chao, M Bronner-Fraser, and PY Lwigale. Stem-cell properties of human corneal keratocytes. ARVO Meeting Abstracts, April 11, 2009, 50:2049.
2. JR Chao, M Bronner-Fraser, and PY Lwigale. Corneal plasticity: characterization of the multipotentiality of human keratocytes. ARVO Meeting Abstracts, April 11, 2008, 49:4812.
Author Disclosure Statement
The authors (J.R.C., M.E.B., and P.Y.L.) have no commercial associations in connection with this article to disclose.