
Abstract
Reprogramming of somatic cells into induced pluripotent stem (iPS) cells by defined pluripotency and self-renewal factors has taken stem cell technology to the forefront of regenerative medicine. However, a number of challenges remain in the field including efficient protocols and the threat of cancer. Reprogramming of plant somatic cells to plant embryonic stem cells using a combination of two plant hormones was discovered in 1957 and has been a routine university laboratory practical for over 30 years. The plant hormones responsible for cell reprogramming to pluripotency, indole-3-acetic acid (IAA) and isopentenyl adenosine (IPA), are present in human cells, leading to the exciting possibility that plant hormones might reprogram mammalian cells without genetic factors. We found that plant hormones on their own could not reprogram mammalian cells but increase the efficiency of the early formation of iPS cells combined with three defined genetic factors during the first 3 weeks of reprogramming by accelerating the cell cycle and regulating pluripotency genes. Moreover, the cytokinin IPA, a known human anticancer agent, reduced the threat of cancer of iPS cell in vitro by regulating key cancer and stem cell-related genes, most notably c-Myc and Igf-1. In conclusion, the plant hormones, auxin and cytokinin, are new small chemicals useful for enhancing early reprogramming efficiency of mammalian cells and reducing the threat of cancer from iPS cells. These findings suggest a novel role for plant hormones in the biology of mammalian cell plasticity.
Introduction
I
Interestingly, reprogramming of plant somatic cells into plant embryonic stem cells using two plant hormones, auxin and cytokinin, was first demonstrated by Skoog and Miller in 1957 [6]. With these plant hormones, following an exhaustive and intensive testing of cell culture conditions in several species, including the model plant, Arabidopsis, plant regeneration systems have been developed [7 –10]. Auxins include several classes of compounds, indole-3-acetic acid (IAA) being the natural auxin found in plants, whereas there are several classes of cytokinins, including isopentenyl adenosine (IPA), a natural compound found in plants that is active in several plant tissues [10]. IPA is known to bind transfer-RNA to protect against oxidation and assist in protein production [11 –13]. Furthermore, the anticancer properties of IPA are very well established in many types of cancer, including leukemia, breast and colon carcinoma, suggesting that they might also reduce the tumorigenicity of iPS cells [14 –16].
Both auxins and cytokinins have been found to be present in human cells suggesting that reprogramming plant cells can be recapitulated in mammalian cells [11]. Plant and human auxin have identical hydrocarbon structures, suggesting potentially conserved roles for these molecules between plants and animals. Given the recent interest in reprogramming fibroblasts to iPS cells and the potential for the application of these cells in regenerative medicine, we sought to investigate if we could reprogram fibroblasts with plant hormones. We show that plant hormones alone cannot reprogram fibroblasts but can increase efficiency of reprogramming with three or four defined genetic factors (Oct4, Sox2, Klf4, c-Myc). Moreover, we found that they specifically act during the first weeks of reprogramming and effectively reduce the threat of cancer from iPS cells. We show for the first time the effect of plant hormones on global gene expression levels of mammalian cells. Interestingly, we demonstrate that oncogenes such as c-Myc are downregulated, pluripotency genes such as Oct4 and Rex1 are upregulated and most notably the aging gene IGF-1 is a target of plant hormones in mammalian cells.
Methods and Materials
Generation of early iPS cells
The generation of iPS cells using mouse embryonic fibroblasts (MEFs) from C57BL/6 mice has been previously described [17]. MEFs were engineered to express an Oct4-promoter-driven GFP tag (Oct4-GFP MEFs) and used at passage 1 to 3 for the reprogramming experiments. Briefly, for reprogramming experiments, 50,000 or 100,000 cells were seeded per well of a six-well plate and infected with retroviral supernatants of FLAG-tagged pBabe Oct4, Sox2, Klf4, and/or c-Myc. Infection consisted of a 45-min spinfection at 750 g, washed with phosphate-buffered saline (PBS), and keratinocyte medium replaced. Two rounds of infections on consecutive days were performed. Two days after beginning the last round of infection, cells were trypsinized and seeded onto feeder layers of irradiated MEFs in the same culture medium. The medium was changed upon platting to G4 with LIF. The medium was supplemented with a ratio of 1:5 μM of auxin:cytokinin or methanol alone as a control, following established protocols for reprogramming plant cells from the first day on feeder layers. Cultures were maintained at 37°C, 5% CO2, with media changes every other day.
Culture of established mouse iPS cells with or without plant hormone
The use of previously characterized and established mouse iPS cells using MEFs from C57BL/6 mice has been previously described [17]. Mouse iPS cells were maintained in G4 media, which was changed every day. The final media formulation included: DMEM (Gibco), 10% fetal bovine serum (FBS, Hyclone), MEM nonessential amino acids (Gibco), penicillin/streptomycin (10 mg/mL each, Gibco), GlutaMAX (200 mM, Gibco), sodium pyruvate (Gibco), 2-mercaptoethanol (50 mM, Gibco), mouse leukemia inhibitory factor (LIF, 1,000 U/mL, Chemicon). We used plant hormones auxin (IAA) and cytokinin (IPA) following a defined University of Barcelona student protocol for reprogramming plant cells and also referred to the method described by the group of Meyerowitz [9]. A ratio of 1:5 μM of auxin:cytokinin or methanol alone as a control was used following established protocols for reprogramming plant cells.
Constructs and virus production
cDNAs for Oct4 and Sox2 were amplified from mouse embryonic stem cell RNA by RT-PCR. Klf4 was amplified from IMAGE clone 5111134. c-Myc T58A mutant cDNA was amplified from DNA kindly provided by Dr. Luciano DiCroce. The amplified cDNAs were cloned into the EcoRI/ClaI sites of a modified pMSCVpuro vector, which allows the expression of N-terminal FLAG-tagged proteins. Retroviruses for the four factors were independently produced after transfecting the cell line Phoenix Amphotropic using Fugene 6 reagent (Roche) according to the manufacturer's directions. After 24 h, the DMEM medium was replaced, cells were incubated at 32°C, and the viral supernatant was harvested after 24 and 48 h.
Immunofluoresence and AP analysis
Cells were grown on plastic coverslide chambers and fixed with 4% paraformaldehyde. Secondary antibodies used were all the Alexa Fluor Series from Invitrogen (all 1:500) as described in the figures. Images were taken using a Leica SP5 confocal microscope. Direct alkaline phosphatase activity was analyzed using an Alkaline Phosphatase Blue/Red Membrane Substrate solution kit according to the manufacturer's guidelines (Sigma).
In vitro differentiation
Differentiation toward endoderm, mesoderm, and neuroectoderm was carried out by plating embryoid bodies (EBs) on gelatin and DMEM medium with 20% FCS changed every second day for 2–3 weeks. Differentiation toward neurons was performed by adding EBs to matrigel thin-coated plates and kept in neural basal medium (50% neural basal medium, 50% F12 medium) with N2/B27 growth supplement and 1 μM retinoic acid for 3 week. Cells were then stained for appropriate markers described in the figures.
Flow cytometry analyses
All analyses were performed on a MoFlo cell sorter (Dako Cytomation) running Summit software. For measuring proliferation, we have used the commercial kit from Invitrogen the Click-iT EdU AlexaFluor647 Flow Cytometry Assay kit following the manufacturer's instructions (LifeTEch). For the proliferation assay using the click-IT kit, instead of using the supplied DNA dyes, we used a home-made DNA DAPI staining solution (0.1 M Tris Base pH 7.4; 0.9% or 150 mM NaCl; 1 mM CaCl2; 0.5 mM MgCl2; 0.2% BSA; 0.1% Nonidet P40; 10 mg/mL DAPI) and 0.5 mL/test (2 h at RT or overnight 4°C). Details of the procedure have been published [18].
Microarray
Microarray was performed at the Val d Hebron Research Institute microarray platform. Mouse Agilent 4×44K, two colors, gene expression arrays were used. Data were analyzed by the same platform and presented either as global gene expression differences or as gene sets based on cell function in the respective figures.
Real-time PCR
Total mRNA was isolated using TRIZOL and 1 μg was used to synthesize cDNA using the Invitrogen Cloned AMV First-Strand cDNA synthesis kit. 1–2 μL of the reaction was used to quantify gene expression by qPCR for transgenes and endogenous pluripotent genes as previously described.
Soft agar assay
Briefly, six-well plates were coated with 1 mL of 0.4% Agar/G4 medium gel, between 25,000 and 2,500 mESC, mouse leukemia cells or iPS cells plated in a 0.3% gel placed on top. LIF (1,000 Units/mL) was maintained in all conditions, but low serum (1% FCS) and high serum (15% FCS) was used in the agar gels to control growth factor conditions. 0.5 mL of G4 media was changed every 3 days at 37°C, 5% CO2, and after 2–3 weeks, stained with 1% commassie blue for 10 min, washed with PBS once and the number of colonies counted. The media and soft agar gel contained 5 μM of cytokinin or methanol alone, as a control throughout the experiment.
Results
Plant hormones increase efficiency of mammalian cell reprogramming to iPS cells
We first sought to test if plant hormones could reprogram mammalian cells on their own. To do this, we first examined if the plant hormone auxin and a panel of plant hormone cytokinin forms (including ZR, zeatin riboside; Z, zeatin; DHZ, dihydrozeatin; K, kinetin; 2-IP, isopentenyl; IPA, isopentenyl adenine) could regulate pluripotency gene expression. We chose the auxin IAA (Fig. 1A) because it is the preferred auxin for plant cell reprogramming while we tested a battery of cytokinins all known for plant cell reprogramming (Fig. 1B). For plant cell reprogramming, a 5:1 ratio of IAA:IPA was used. We found that after a series of experiments for mammalian cells the reverse ratio of 1:5 IAA:IPA had the best effect on mammalian pluripotency gene expression (Supplementary Fig. S1 and S2; Supplementary Data are available online at

Given we had shown that plant hormones could regulate pluripotency genes, we next tested if the ratio of 1 μM auxin IAA:5 μM cytokinin IPA could reprogram MEFs by their own or in combination with three or four standard reprogramming factors Oct4, Sox2, Klf4, and c-Myc. We found that plant hormones cannot reprogram MEFs on their own nor could they reprogram MEFs in combination with one factor (Oct4) or with two factors (Oct4/Sox2) (Fig. 1C). As a control for this experiement, we observed with three factors or four factors iPS cell colonies at 2–3 weeks, as expected.
We next asked the question if plant hormones could increase the efficiency of reprogramming with defined factors. We engineered a MEF cell line with an Oct4-promoter-driven GFP reporter gene (Oct4-GFP MEFs) and reprogrammed these cells either three factors (Oct4, Sox2, Klf4) or four factors (plus c-Myc) with and without 1 μM auxin IAA:5 μM cytokinin IPA treatment. We demonstrate that in the presence of plant hormones an increase in the number of colonies and intensity of GFP signal during the early (3 to 4 weeks) stage of reprogramming by colony formation assay (Fig. 1D). We show by microarray gene expression analysis that plant hormones can increase the expression of the key pluripotency genes Oct4 and Nanog during the early reprogramming period (Fig. 1D). To quantify the colony formation assay, we used flow cytometry to measure the number of GFP-positive cells during reprogramming of Oct4-GFP-MEF cells with and without plant hormone treatment. In the presence of IAA and IPA, Oct4-GFP-MEF demonstrated a twofold increase in the number of GFP-positive cells after 3 weeks of reprogramming with Oct4/Sox2/Klf4/c-Myc (Fig. 1E). Taken together, we demonstrate that 1 μM auxin:IAA 5 μM cytokinin IPA plant hormone treatment of MEFs can increase the efficiency of reprogramming twofold by regulating the expression of key pluripotency genes.
Plant hormones auxin and cytokinin regulate cell cycle of mammalian cells during reprogramming
We next sought to understand the mechanism by which plant hormones auxin IAA and IPA can increase the efficiency of reprogramming. It is well established that IPA regulates the cell cycle of plant cells and that IAA regulates plant cell senescence. Therefore, we analyzed the effect of IPA and IAA on the cell cycle of the MEFs by flow cytometry using EdU staining. We found that plant hormones can increase the S-phase stage of the cell cycle by approximately double with a concurrent reduction of cells in the G1-phase of the cell cycle (Fig. 2A). We found that there is little to no effect on apoptosis of MEFs during the early reprogramming stage (Fig. 2A).
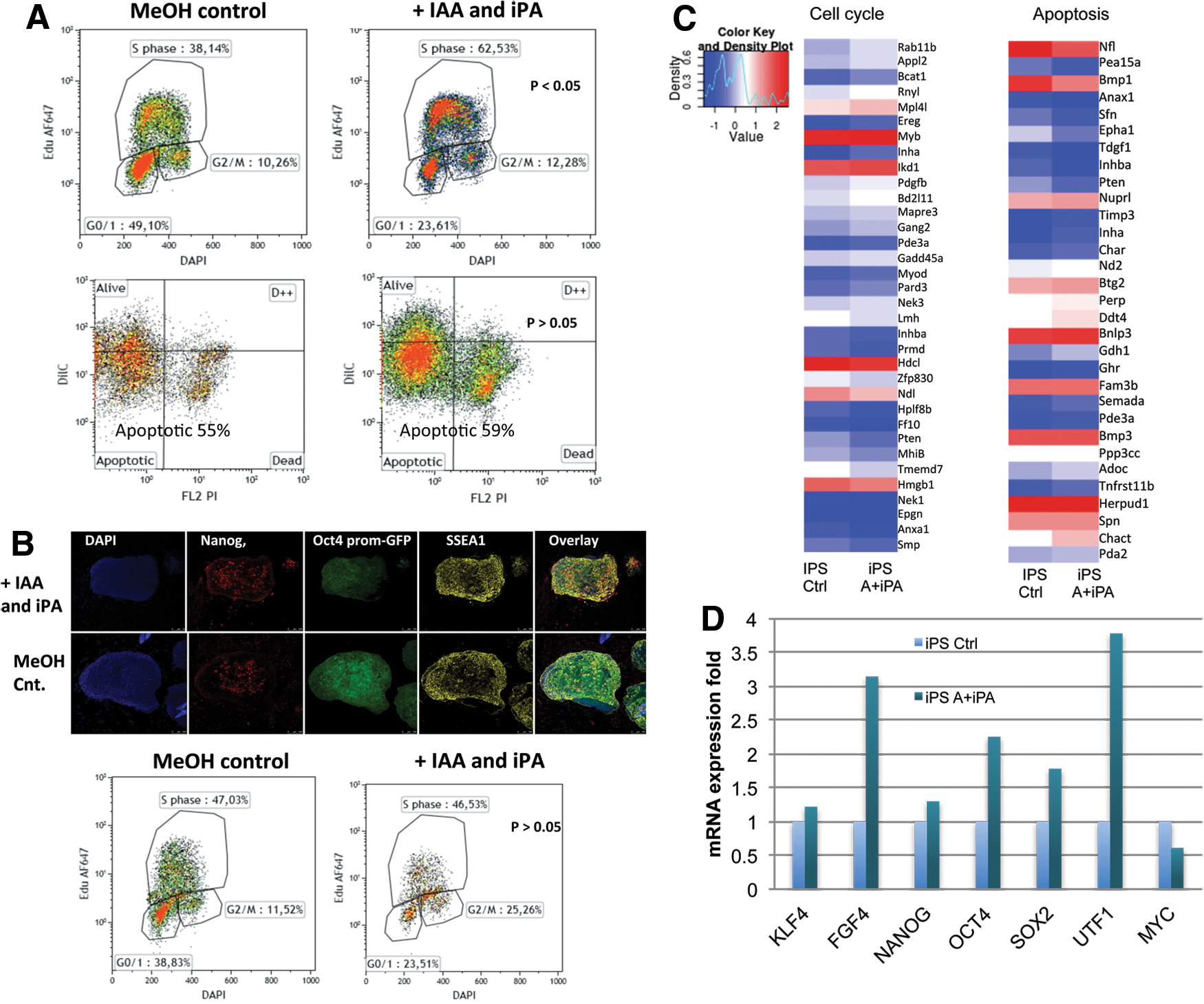
To determine if plant hormones affected the pluripotency of iPS cells over long term, we characterized iPS cells after long-term treatment (longer than 4 weeks) and demonstrated no detectable differences in pluripotency marker expression for Nanog and SSEA1 (Fig. 2B). Moreover, we did not detect an effect on the S-phase with plant hormone treatment of established iPS cells, suggesting that plant hormones only act during the early reprogramming stages (Fig. 2B).
To understand in more detail the mechanism, we performed a microarray analysis of the effect of plant hormones on mouse cell cycle and apoptosis genes with and without treatment with plant hormones. We found that there are a number of cell cycle-related genes expression upregulated by plant hormones (Fig. 2C). Moreover, we show by real-time PCR analysis that pluripotency genes FGF4, Oct4, Sox2, and UTF1 gene expression are upregulated with plant hormone treatment in early reprogramming cells (Fig. 2D). Interestingly, we show that plant hormone treatment reduced the expression of the oncogene c-Myc during reprogramming (Fig. 2D). Taken together, we show that the mechanism of action of plant hormones to increase efficiency of reprogramming of mammalian cells is by accelerating the cell cycle and increasing pluripotency gene expression levels, without significantly affecting apoptosis levels, and this effect does not affect established iPS cells (Fig. 2B).
Cytokinin IPA reduces tumorigenicity of iPS cells
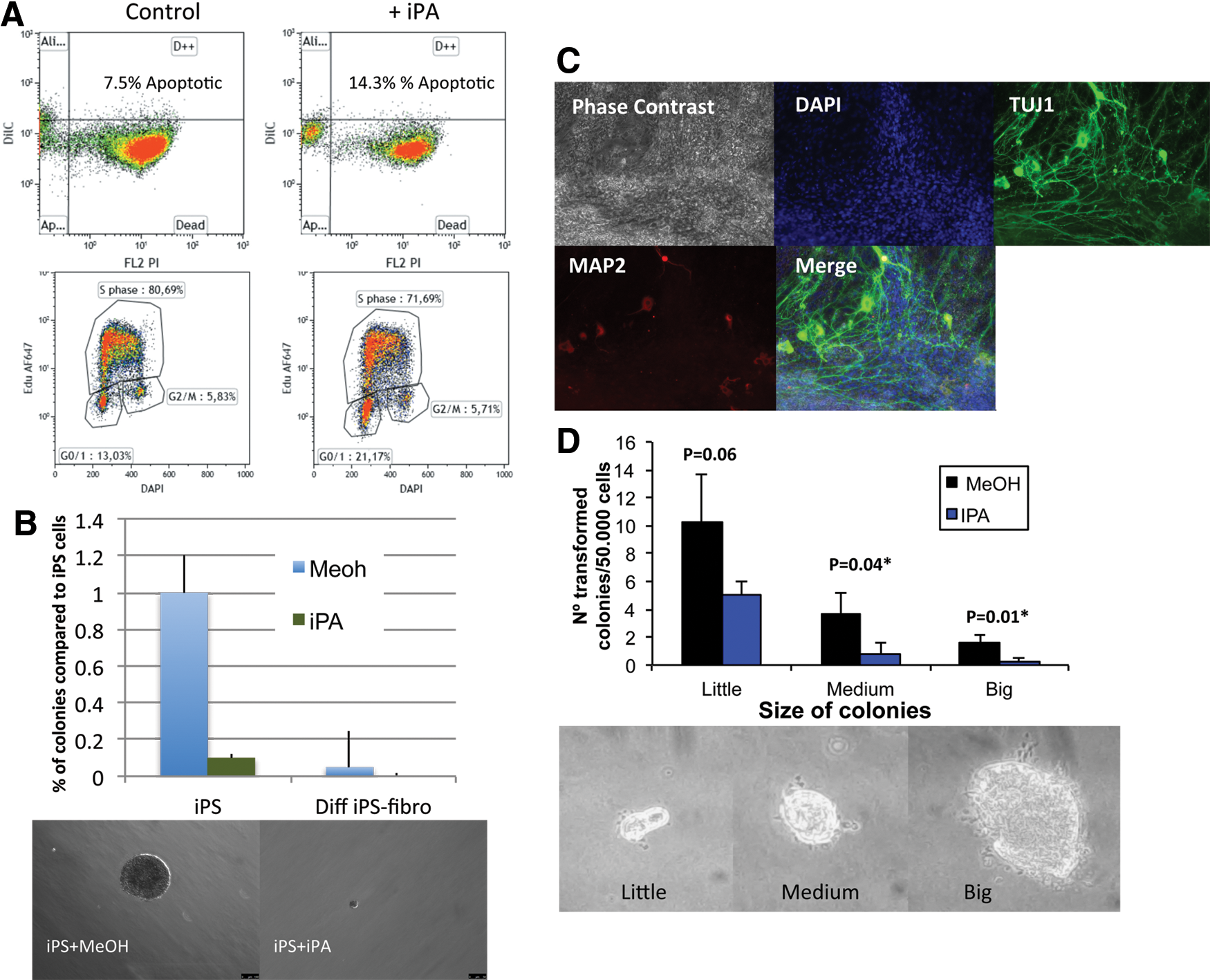
Given our observation that plant hormone treatment reduced c-Myc expression (Fig. 2D) and that IPA is a well-established anticancer agent in many different cancer types, we next asked if plant hormone treatment could reduce the tumorigenicity of iPS cells their differentiated progeny in vitro. We confirmed previous published data that plant hormone IPA can kill mouse leukemia cancer cells by slowing the cell cycle and increasing apoptosis (Fig. 3A). Next, we differentiated iPS cells under general conditions (DMEM plus 20% FBS) and guided conditions toward neuronal stem cells (retinoic acid plus N2/B27 in F12/DMEM) and tested the ability of IPA to reduce the threat of cancer. We used an in vitro soft agar approach to test a major hallmark of malignant cancer, namely, anchorage-independent and growth factor-deprived colony formation. We show from repetitive experiments a reduced colony formation in both iPS cells and fibroblast-like cells from general differentiation conditions are significantly reduced with the treatment of plant hormone IPA (5 μM) (Fig. 3B). Importantly, we show that plant hormone can reduce the ability of differentiated fibroblast-like cells to undetectable levels in a soft agar assay (Fig. 3B). In addition, using directed differentiation toward neuronal stem cells over 4 weeks in a soft agar assay iPA treatment reduced colony formation in soft agar (Fig. 3C, D). Histological analysis by immunofluorescence demonstrated the neural stem cells were positive for TUJ1 and MAP2 markers (Fig. 3C). Soft agar assay of the neural stem cells with and without plant hormone cytokinin IPA significantly reduced the ability of these cells to grow in anchorage-independent and growth factor-deprived conditions (Fig. 3D). Taken together, the data demonstrate plant hormone IPA treatment of iPS cells and differentiated progeny. The differentiated stem cells would in theory be the cell state used for cell replacement therapy and plant hormone IPA can significantly reduce the threat of cancer to low or undetectable levels.
Cytokinin downregulates mammalian IGF-1 gene expression
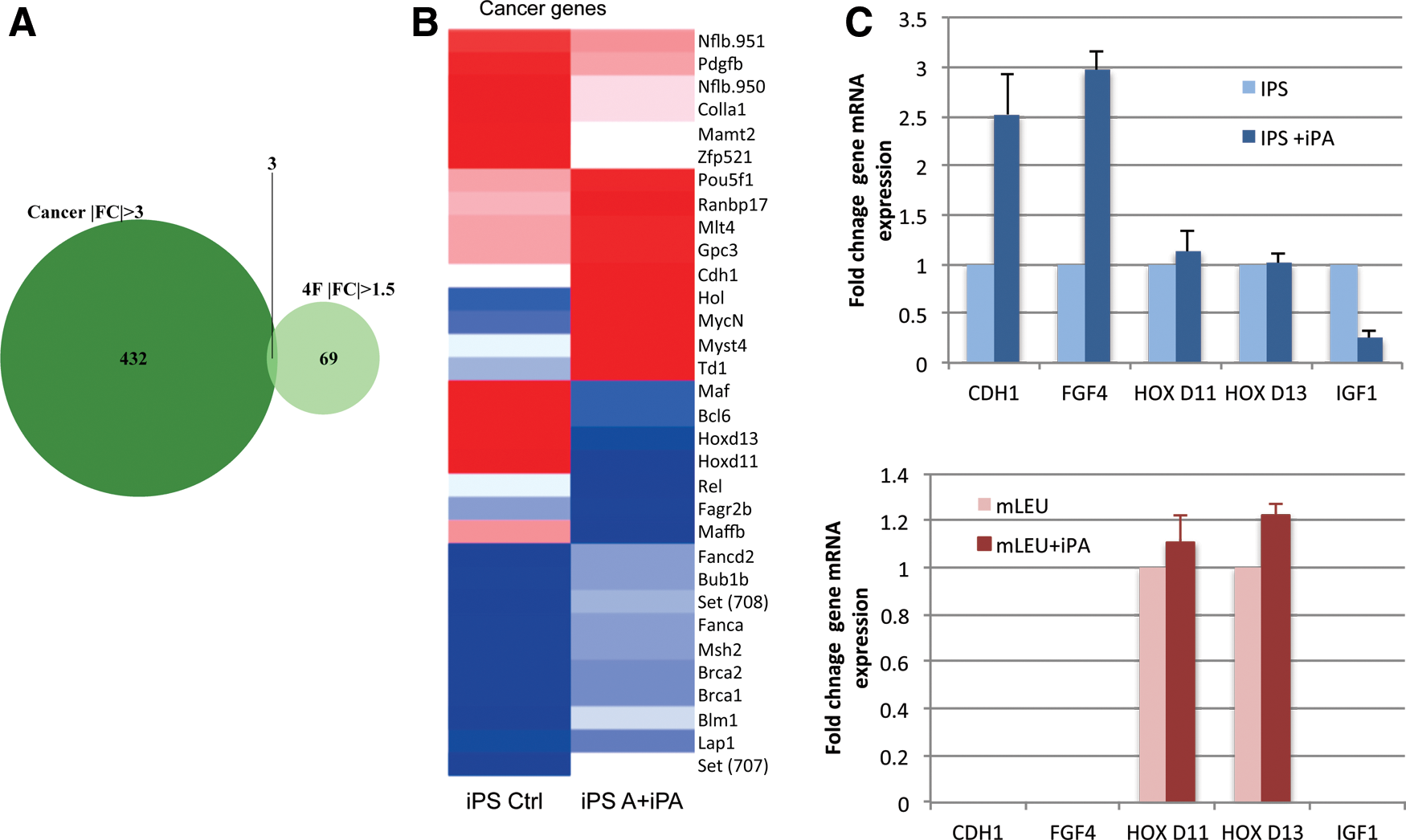
To investigate further how IPA plant hormone treatment is reducing the ability of iPS cells to form cancer, we performed a microarray gene expression analysis. We found from all cancer-related gene expression (mouse leukemia cell line) that IPA regulates the expression of 432 mammalian cancer-related genes in iPS cells and 69 cancer-related genes in mouse cancer (leukemia) cells (Fig. 4A). Only three genes were found to be regulated by IPA in both iPS cells and cancer cells (Fig. 4A). To validate the microarray experiment, we analyzed four genes that are of high interest to the field of cancer research, Cdh1, Igf-1, Hox d11, and Hox d13 by real-time PCR gene expression analysis. We found that the Hox genes were not validated in iPS cells; however, Cdh1 mouse gene expression is upregulated threefold by plant hormone and Igf-1 gene expression is downregulated fourfold (Fig. 4C). Taken together, the data demonstrate that plant hormone IPA treatment acts on key cancer-related genes c-Myc, Cdh1, and Igf-1 gene expression. These data suggest that plant hormone IPA reduces the threat of cancer of iPSC via these signaling pathways, most notably the suppression of c-Myc (Fig. 2D) and Igf-1 gene expression (Fig. 4C). This mechanism may be specific to iPS cells because we did not detect any change in a mouse leukemia cell line in Cdh1, Fgf4, Hox d11, Hox d13, or Igf-1 gene expression levels with plant hormone IPA treatment (Fig. 4C).
Discussion
Plant cells, unlike animal cells, have the ability to regenerate themselves in response to injury using specific combinations of plant hormones. In 1957, it was discovered that plant hormones could reprogram plant somatic cells to plant embryonic stem cells that can recapitulate the development of an entire plant (Skoog, 1957 #65). Here, we have investigated if plant hormones auxin and cytokinin IPA can reprogram mammalian cells to pluripotency. Our findings demonstrate that IAA:IPA treatment alone is insufficient to reprogram mammalian cells to pluripotency. However, when combined with traditional three- and four-factor reprogramming protocols, plant hormone treatment increased the efficiency of reprogramming by accelerating the cell cycle and increasing pluripotent gene expression levels. Addition of plant hormone to established iPS cell clones did not alter the cell cycle nor pluripotency markers SSEA1 and Nanog, suggesting that the effect of the plant hormones affect the reprogramming process (Fig. 2B). Further work to characterize iPS cells made with plant hormone treatment is warranted; however, our data suggest that the quality of the iPS cells will be increased in the presence of IAA:IPA, given that they regulate key pluripotency genes during reprogramming. Furthermore, we demonstrate that the cytokinin IPA reduces oncogenic transformation of iPS cells and their differentiated progeny to very low levels in vitro.
These findings have a number of far reaching implications in the field of reprogramming and in regenerative medicine. First, the use of plant hormones could become a routine small chemical to increase efficiency of making iPS cells, similar to HDAC and MEK inhibitors. Second, characterization of the mechanism by which plant hormone treatment can regulate mammalian pluripotency genes could reveal new targets for reprogramming. Third, the use of cytokinin IPA would be an ideal naturally occurring small chemical compound for routine use to reduce the threat of cancer posed by iPS cell-derived cells for cell replacement therapy. Although it remains to be tested if IPA treatment of iPS cells and differentiated progeny can reduce cancer in vivo, this work has established the first observation that IPA, a well-known anticancer agent [14 –16], can reduce oncogenic transformation of iPS cells.
This study is the first of its kind to explore the effect of plant hormones auxin and cytokinin on mammalian cell gene expression levels. The data reveal interesting gene targets of plant hormones on mammalian genes, such as c-Myc, Cdh1, Fgf4, and Igf-1, all involved in cancer and stem cell biology. Cytokinin IPA suppression of oncogenic transformation in iPS cells most likely acts through its suppression of c-Myc gene expression (Fig. 1) and regulation of Cdh1 and Igf-1 and gene expression. Cdh1 encodes the E-cadherin protein and is upregulated by plant hormones in developing iPS cells. Loss of E-cadherin function or expression has been implicated in cancer progression and metastasis suggesting that plant hormones reduce the threat of cancer of iPS by upregulating E-cadherin. Cdh1 is emerging as a key regulator of human and mouse stem cell pluripotency and self-renewal; however, it remains to be seen if loss-of-function and gain-of-function studies can confirm this finding. Insulin-like growth factor (IGF) signaling mediates many critical cell responses, including mitogenesis, proliferation, growth, differentiation, and angiogenesis, and is downregulated by plant hormones during reprogramming of mammalian cells (Fig. 4C). The IGF signaling pathway has a pathogenic role in cancer, and recent studies have shown that increased levels of IGF lead to increased growth of existing cancer cells [19]. The regulation of these genes by plant hormones strongly suggests that IAA and IPA have important roles in both the regulation of the pluripotent state as well as the cellular defenses against oncogenesis.
In conclusion, given that the chemical structure of plant and human auxin is identical and that cytokinin IPA is found in human cells bound to tRNA assisting protein expression, presumably supplied by the consumption of vegetables in the diet, and that we observe a positive effect of plant hormones on pluripotency and key anticancer gene expression levels in mammalian cells, it is likely that plant hormones could offer a method for increasing the safety of iPS cells for therapeutic applications. While loss and gain of function studies as well as in vivo studies are needed to further define the mechanism of plant hormone action on Igf-1, c-Myc, and Cdh1 genes during iPS reprogramming, this study demonstrates a novel use of plant hormones for increasing the efficiency and safety of iPS cells for cell replacement therapy and regenerative medicine in the near future.
Footnotes
Acknowledgments
M.J.E. (RYC-2010-06512) is supported in part by the Program Ramon y Cajal from the MICINN and supported by a grant from MICINN BFU2011-26596. Additional support was provided by grant from the Fondo de Investigaciones Sanitarias (P1071209). The authors thank Dr. Jordi Mestres for comparative analysis of the chemical structure of plant and human hormones; Advanced Optical Microscopy Unit, University of Barcelona; and the Institut de Recerca Hospital, Val d'Hebron, UCTS.
Author Disclosure Statement
The authors declare no competing financial interests.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.