
Abstract
In spina bifida, the neural tube fails to close during the embryonic period. Exposure of the neural tube to the amniotic fluid during pregnancy causes additional neural damage. Intrauterine tissue engineering using a biomaterial seeded with stem cells might prevent this additional damage. For this purpose, autologous cells from the amniotic fluid are an attractive source. To close the defect, it is important that these cells deposit an extracellular matrix. However, it is not known if amniotic fluid mesenchymal cells (AFMCs) from a fetus with a neural tube defect (NTD) share the same characteristics as AFMCs from a healthy fetus. We found that cells derived from fetuses with a NTD, in contrast to healthy human amniotic fluid cells, did not deposit collagen type I. Furthermore, the NTD cells showed, compared with both healthy amniotic fluid cells and fetal fibroblasts, much lower mRNA expression levels of genes that are involved in collagen biosynthesis [procollagen C-endopeptidase enhancer proteins (PCOLCE), PCOLCE2, ADAM metallopeptidase with thrombospondin type 1 motif, 2 (ADAMTS2), ADAMTS14]. This indicates that NTD-AFMCs have different characteristics compared with healthy AFMCs and might not be suitable for fetal therapy to close the defect in spina bifida patients.
Introduction
I
Amniotic fluid contains a population of mesenchymal progenitor cells that can be used as a cell source for fetal surgical therapy. This concept has been introduced about a decade ago by the group of Fauza [12 –19]. In short, some congenital anomalies detected by prenatal imaging can, in principle, be surgically treated with autologous amniotic fluid mesenchymal stem cells (AFMCs). These cells can be isolated and expanded from the amniotic fluid obtained from amniocentesis. By seeding these cells in a scaffold, a fetus can benefit from such an autologous graft in surgical procedures aimed at repairing the congenital anomaly (fetal tissue engineering) [20 –23]. Several reports describe the differentiation of the multipotent AFMCs into cell types such as adipocytes, chondrocytes, osteoblasts, or smooth muscle cells [24 –31]. Cartilage and tendon grafts derived from AFMCs have successfully been used in fetal repair efforts in large animal models [14,18,19]. In a previous study, we showed that AFMCs from healthy fetuses can be used to repair skin defects [32].
Since the NTD coverage must be complete to prevent leakage of amniotic fluid to the underlying tissues, we questioned whether AFMCs can be used to accelerate the repair in utero. For future therapy, autologous AFMCs are preferred, to reduce the chance of rejection. However, no studies have been performed with AFMCs derived from fetuses with a NTD; therefore, it is not known whether these cells can be used for tissue engineering. In the current study, we have investigated whether AFMCs from NTD fetuses can be used as an autologous source for skin tissue engineering by analyzing their ability to produce collagen type I and therefore their capability of closing the defect.
In tissue engineering, mesenchymal progenitor cells need to be differentiated into specialized cells to produce the required connective tissue (eg, chondrocytes to produce cartilage, or osteoblasts to produce bone). However, mesenchymal progenitor cells have many fibroblast-like properties [33 –36]. We hypothesized that a one-step procedure might be possible, namely that amniotic fluid cells derived from a fetus with a neural tube defect (NTD-AFMCs) can be placed in the biomaterial without performing a differentiation step, and that these AFMCs subsequently produce a covering connective tissue rich in collagen type I.
Materials and Methods
Cell culture
Confluent back-up human amniocentesis cultures were received from the clinical cytogenetics laboratory of the University Medical Center Groningen. Cells were cultured from healthy fetuses (H-AFMCs, n=4, with a normal karyotype) and NTD-AFMCs (n=4, with a normal karyotype) in uncoated plates (Corning B.V. Life Sciences, Amsterdam, the Netherlands). Cells were harvested and expanded in the DMEM (Lonza, Breda, the Netherlands) supplemented with 20% fetal calf serum (FCS; Perbio Science, Etten-Leur, the Netherlands) and 1% penicillin, 1% streptomycin (10,000 U/mL; Gibco, Paisley, United Kingdom), and 2 mM L-glutamine (Lonza). After passage 4, cells were cultured in the DMEM supplemented with 10% FCS, 1% penicillin, 1% streptomycin, and 2 mM L-glutamine.
Primary fetal human dermal fibroblasts [HDF-f; Sciencell (#2300), Carlsbad, CA and Cell Applications (106-05f ), San Diego, CA] were cultured in the DMEM supplemented with 10% FCS, 1% penicillin, 1% streptomycin, and 2 mM L-glutamine.
Fluorescence-activated cell sorting
AFMCs (from passage 5–7) were incubated with FITC-conjugated CD34 antibodies [1:10; IQ products (IQP-114F), Groningen, the Netherlands], FITC-conjugated CD44 antibodies [1:10; IQ products (IQP-118F)], FITC-conjugated SSEA4 antibodies [1:10; Abcam (ab16287), Cambridge, United Kingdom], PE-conjugated CD45 antibodies [1:10; IQ products (IQP-124R)], and PE-conjugated CD90 antibodies [1:10; R&D systems (FAB2067P), Abingdon, United Kingdom] at 4°C for 30 min, followed by a wash step in PBS containing 0.2% bovine serum albumin (BSA; Sanquin, Amsterdam, the Netherlands). For Oct4 staining, cells were first fixed with 2% paraformaldehyde (Merck, Darmstadt, Germany) for 10 min, permeabilized, and incubated with an unconjugated Oct4 antibody [1:100; Abcam (ab19857)] for 30 min at 4°C. Cells were washed and incubated for 15 min with swine anti-rabbit TRITC (1:20; Dako, Glosstrup, Denmark), followed by a wash step in PBS containing 0.2% BSA. Cells were analyzed using a FACSCalibur system (BD Biosciences, San Jose, CA). Data were analyzed by FlowJo software (Tree Star, Ashland, OR).
Collagen I deposition by HDF-f, H-AFMCs, and NTD-AFMCs after transforming growth factor beta1 stimulation
HDF-f [ATCC (CCL-110), Manassas, VA], H-AFMCs, and NTD-AFMCs were seeded at a density of 15,000/cm2 in Permanox chamberslides (Nunc, New York, NY) in the DMEM with 10% FCS. After 24 h, the medium was removed and replaced with a medium containing 0.5% FCS and 50 ng/mL L-ascorbic acid 2-phosphate sesquimagnesium salt hydrate (Sigma-Aldrich, Zwijndrecht, the Netherlands) overnight. Subsequently, cells were exposed with or without 10 ng/mL recombinant human transforming growth factor beta1 (TGF-β1; Peprotech, Rocky Hill, NJ) for 3 days, with daily medium replacements.
After 3 days of culture (with or without TGF-β1), cells were washed twice with PBS and fixed in 2% paraformaldehyde at room temperature for 10 min. Fixed cells were incubated with mouse anti-human collagen I [1:100; Abcam (ab90395)] diluted in PBS containing 1% BSA for 1 h at room temperature. After three washes with PBS, cells were incubated with biotinylated goat anti-mouse IgG1 (1:100; SouthernBiotech, Birmingham, AL) diluted in PBS containing 1% BSA for 30 min at room temperature. Then, the cells were washed three times with PBS and incubated with streptavidin-CY3 (1:400; Dako) in PBS containing 1% BSA, 2% normal human serum, and DAPI (1:5,000) for 30 min. After three washes with PBS, slides were mounted in Citifluor (Agar Scientific, Stansted, United Kingdom) and visualized using a Leica DMRA immunofluorescence microscope and Leica software (Leica Microsystems, Wetzlar, Germany).
Gene expression analysis
HDF-f, H-AFMCs, and NTD-AFMCs (for all cell type passages 5 or 6) were seeded at a density of 15,000/cm2 in uncoated 6-well plates (Corning B.V. Life Sciences) in the DMEM supplemented with 10% FCS, 1% penicillin, 1% streptomycin, and 2 mM L-glutamine. After 3 days of culture, total RNA was isolated from the cells using the RNeasy Micro Kit (Qiagen, Hilden, Germany) according to the manufacturer's protocol. Reverse transcription was carried out using the First Strand cDNA synthesis kit (Fermentas, St. Leon-Rot, Germany). Expression levels of genes known to be involved in collagen biosynthesis and homeostasis (Table 1) were analyzed with a custom made microfluidic card-based low density array (Table 1; Applied Biosystems, Foster City, CA) using the ViiA™ 7 Real-Time PCR System (Applied Biosystems). For each sample, 100 ng cDNA was diluted in 50 μL of distilled water and mixed with 50 μL of TaqMan PCR master mix (Applied Biosystems). Standard recommended PCR protocols were performed (50°C for 2 min, 95°C for 10 min, and the next two steps were repeated for 40 cycles: 95°C for 12 s and 60°C for 1 min). Relative mRNA levels were calculated as 2−ΔCT, in which ΔCT is CT value of target gene − CT value of GAPDH.
Statistical analysis
All data are represented as mean±standard error of the mean of at least three independent experiments and were analyzed by one-way ANOVA followed by the Tukey's post hoc analysis test using GraphPad Prism Version 5 (GraphPad Software, Inc., La Jolla, CA). Values of P<0.05 were considered to be statistically significant.
Results
Characterization of amniotic fluid-derived cells
Human amniotic fluid cells contained a heterogeneous cell population (containing cells with an epithelial and mesenchymal morphology) at the first passage. After five passages, all cells showed a mesenchymal phenotype. Cell populations derived from both healthy fetuses (H) and fetuses with a NTD showed a spindle-shaped morphology (Fig. 1A). No differences were observed in morphology between these two cell populations. For characterization, amniotic fluid mesenchymal cells (AFMCs) were analyzed by FACS for markers that are used for mesenchymal stem cell characterization. Both cell populations were negative (<4% positive cells) for hematopoietic markers CD34 and CD45 (Fig. 1B). Furthermore, healthy and NTD-AFMCs were positive for mesenchymal markers CD44 (>98%) and CD90 (>65%) and embryonic stem cell markers SSEA4 (>70%) and Oct4 (>94%). The expression pattern of these markers indicates that both populations contain predominantly amniotic fluid derived-mesenchymal stem cells. No significant differences for all 6 markers were detected between healthy and NTD-AFMCs, indicating that, based on these markers, a similar cell population can be isolated from healthy fetuses and fetuses with a NTD. This suggests that NTD-AFMCs might be used for fetal tissue engineering.
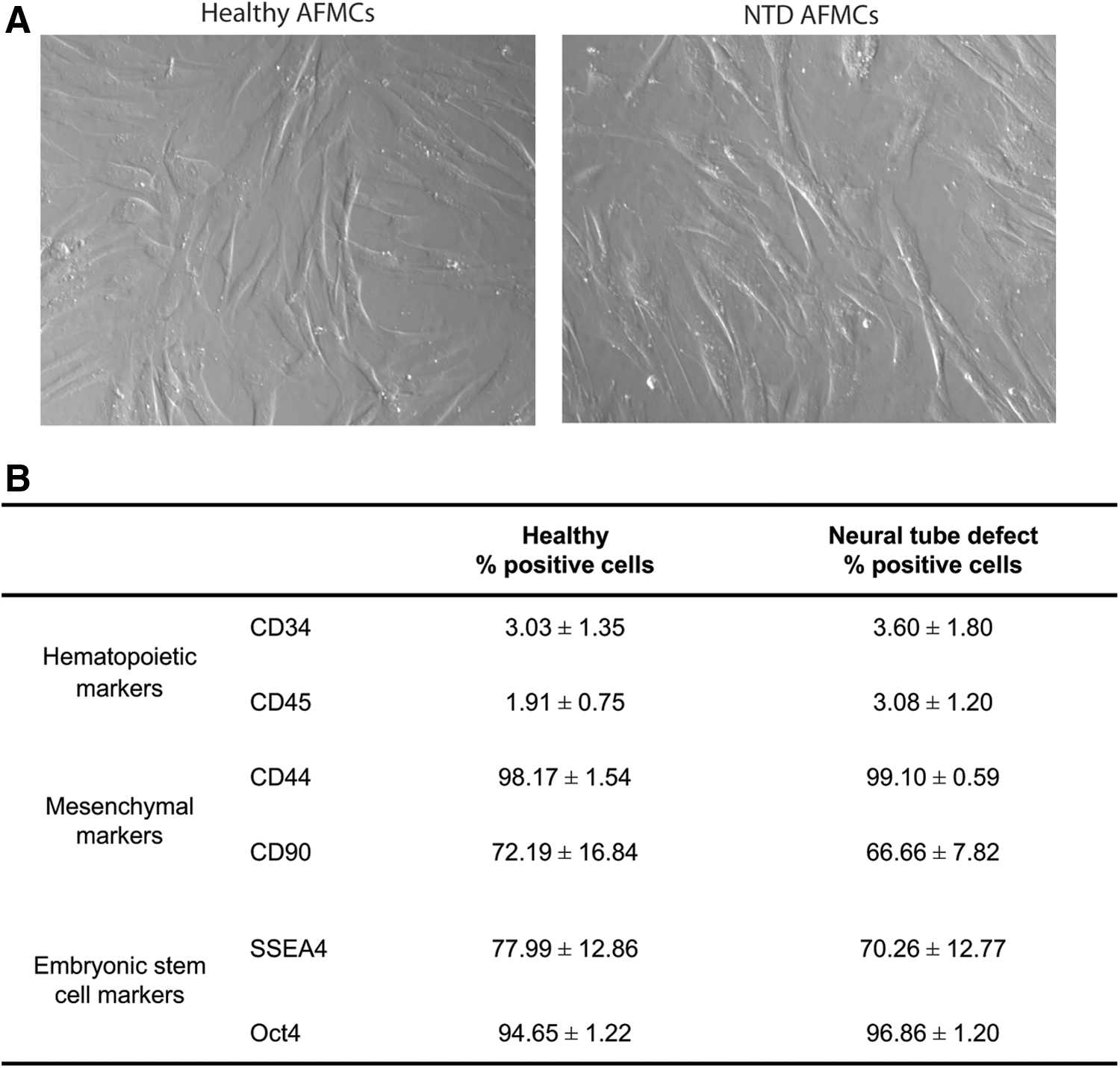
Characterization of amniotic fluid-derived mesenchymal cells.
Collagen type I deposition by HDF-f, H-AFMCs, and NTD-AFMCs after TGF-β1 stimulation
For skin tissue engineering, the formation of a new extracellular matrix (ECM) is a prerequisite. The most abundant ECM protein in fetal skin is collagen type I, which is produced by fetal fibroblasts. To stimulate collagen deposition by fibroblasts, the profibrotic cytokine TGF-β1 can be used. We investigated whether TGF-β1 stimulation also increases collagen deposition by AFMCs. Collagen type I deposition was observed by HDF-f after 3 days of TGF-β1 stimulation (Fig. 2A), as was the case with H-AFMCs (Fig. 2B). However, no collagen type I was deposited by NTD-AFMCs (Fig. 2C). These results indicate that NTD-AFMCs fail to produce mature collagen type I.
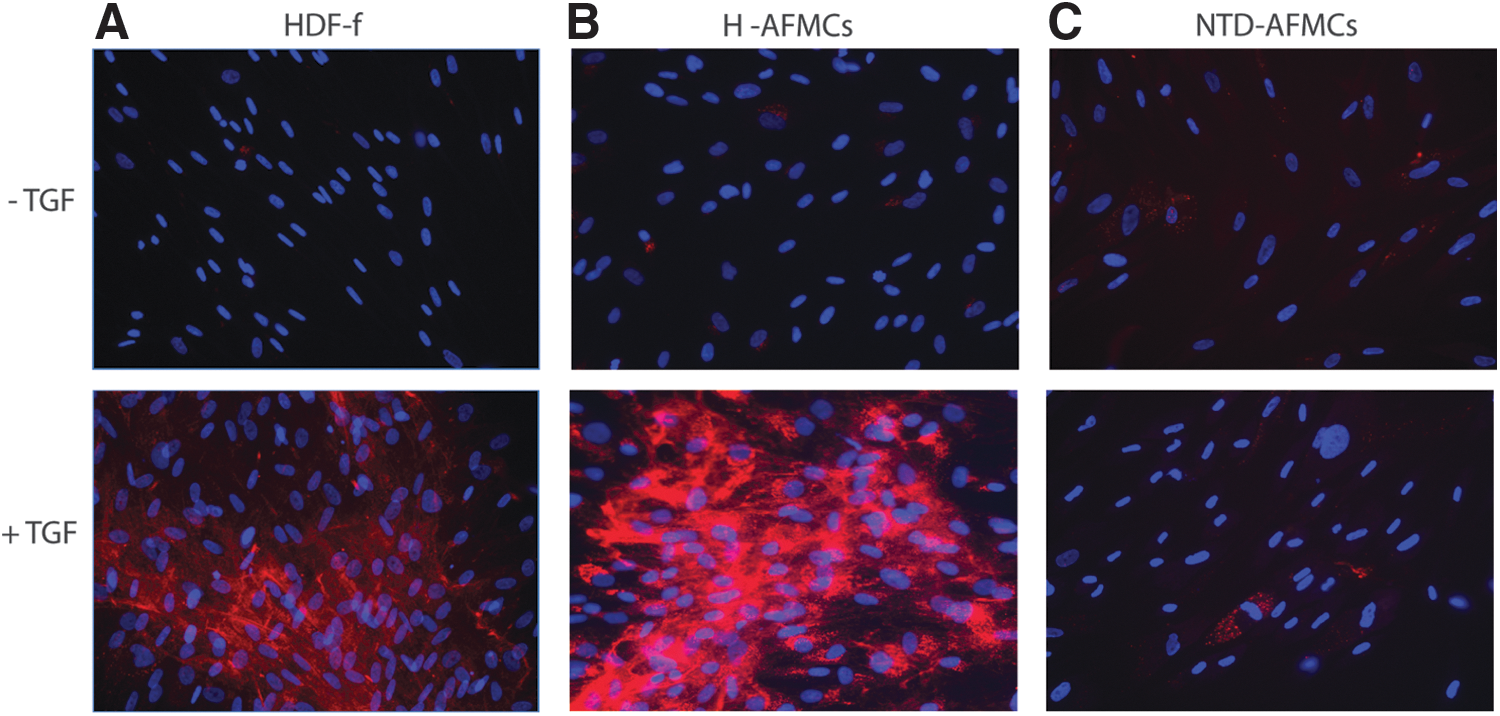
Collagen type I deposition by human fetal dermal fibroblasts, healthy AFMCs, and NTD-AFMCs after TGF-β1 stimulation. COL1A1 protein staining on TGF-β1 stimulated HDF-f
Expression pattern of genes involved in collagen synthesis
Collagen biosynthesis is a complex process that involves numerous modifying, folding and processing enzymes. Differences in expression levels of these enzymes might give an explanation as to why NTD-AFMCs are not able to deposit collagen I protein. We investigated the expression levels of genes that are involved in collagen biosynthesis and homeostasis of HDF-f, H-AFMCs, and NTD-AFMCs with a custom made microfluidic card-based low density array.
Collagen synthesis starts intracellular with the synthesis of α-chains of preprocollagen. Collagen type I is composed of a triple helix, which consists of two identical α1 chains (COL1A1) and one α2 chain (COL1A2). Healthy and NTD-AFMCs show a similar expression of COL1A1 compared with HDF-f (Fig. 3A). However, expression of COL1A2 in NTD-AFMCs was very low (NTD-AFMCs have a 70- and 80-fold lower expression compared with HDF-f (P<0.05) and h-AFMCs (P<0.05), respectively). Collagen type III is composed of a triple helix consisting of three α1 chains (COL3A1). No significant differences were observed between HDF-f, H-AFMCs, and NTD-AFMCs, although NTD-AFMCs showed a sixfold lower expression of COL3A1 compared with HDF-f (P=0.128) and healthy AFMCs (P=0.154). An important step of collagen synthesis is the 4-hydroxylation of proline residues to obtain 4-hydroxyproline, this step involves the enzymes prolyl 4-hydroxylase, alpha polypeptide (P4HA) 1, P4HA2, and P4HA3. No significant differences were found in the expression of these genes between all three cell types (data not shown). In collagen, certain lysine residues can be hydroxylated by procollagen-lysine 1, 2-oxoglutarate 5-dioxygenase (PLOD) 1, PLOD2, and PLOD3 to form hydroxylysine. No significant differences were found in PLOD1 and PLOD2 expression levels, but PLOD3 is significantly lower expressed by NTD-AFMCs compared with H-AFMCs (P<0.05) and HDF-f (P<0.05) (Fig. 3B). Subsequently, some of the hydroxylysine residues can be glycosylated. Galactose can be added to the residues by glycosyltransferase 25 domain containing 1 (GLT25D1) and GLT25D2. No differences were found in the expression of these two genes between the cell types (data not shown).

Expression pattern of genes involved in collagen synthesis. Gene expression levels of
After hydroxylation of proline and lysine and glycosylation of hydroxyprolines, the three alpha chains assemble to form the triple helix resulting in procollagen. Next, procollagen is secreted and N- and C-propeptides are cleaved, to form mature collagen. Bone morphogenetic protein 1 (BMP1), also known as procollagen C proteinase, cleaves the C-propeptides of procollagen. No differences in BMP1 gene expression were found between HDF-f and AFMCs (Fig. 3C). PCOLCE and PCOLCE2 are known to increase the catalytic activity of BMP1. PCOLCE is expressed by HDF-f and H-AFMCs, but was not expressed by NTD-AFMCs. PCOLCE2 is much lower expressed in NTD-AFMCs and H-AFMCs compared with HDF-f (both P<0.001). The N-propeptides are cleaved off from procollagens by the metalloproteinases ADAMTS2, ADAMTS3, and ADAMTS14. A significant lower expression of ADAMTS2 and ADAMTS14 (Fig. 3D) was observed in NTD-AFMCs compared with HDF-f (P<0.05) and H-AFMCs (P<0.05). No differences were found in ADAMTS3 expression. Overall, these results indicate that NTD-AFMCs show impaired synthesis and processing of collagen compared with H-AFMCs.
Expression pattern of genes involved in collagen degradation
Another explanation that NTD-AFMCs do not show collagen protein deposition might be that secreted collagen is quickly degraded by proteolytic enzymes. Therefore, we investigated the expression levels of enzymes that are involved in matrix degradation (Fig. 4). Matrix metalloproteinase 1 (MMP1) expression is 10-fold lower in NTD-AFMCs (P<0.01) and 6.5-fold lower expressed in H-AFMCs (P<0.01) compared with HDF-f. The activity of MMPs can be inhibited by tissue inhibitor matrix metalloproteinase inhibitor 1 (TIMP1). No differences were found in TIMP expression. Cathepsin K (CTSK) has been described to play a role in dermal ECM turnover [37,38]. In this study, we show that HDF-f and H-AFMCs express CTSK, but that is not expressed by NTD-AFMCs (P<0.05).
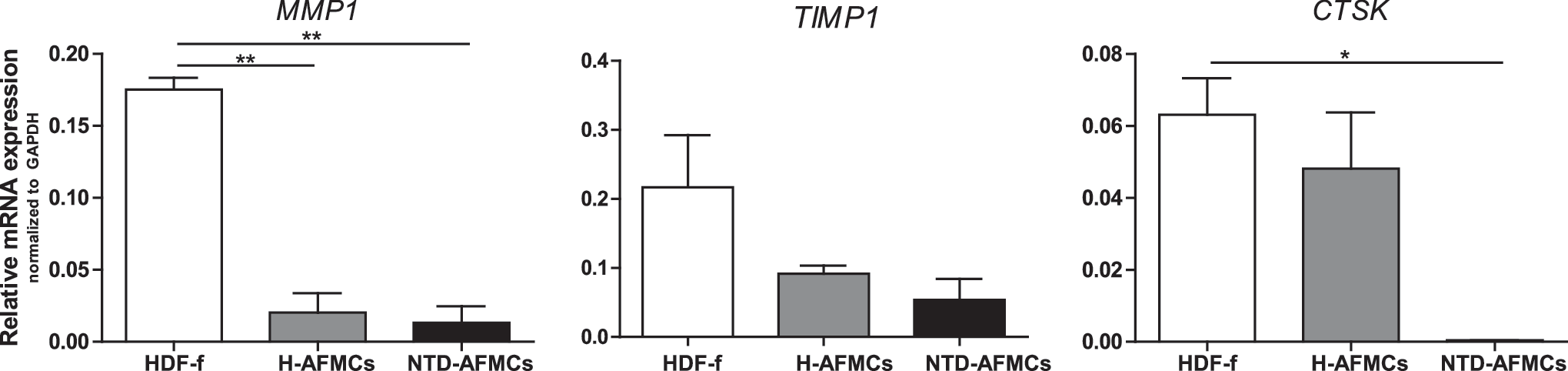
Expression pattern of genes involved in collagen degradation. Gene expression levels of MMP1, TIMP1, and CTSK in HDF-f, H-AFMCs, and NTD-AFMCs. *P<0.05, **P<0.01. CTSK, cathepsin K; MMP1, matrix metalloproteinase 1; TIMP1, tissue inhibitor matrix metalloproteinase inhibitor 1.
These results indicate that the lack of collagen deposition by NTD-AFMCs cannot be caused by an increased expression of enzymes with a collagenolytic activity.
Discussion
Neurological damage of spina bifida has been explained by the two-hit hypothesis: the primary closure defect and subsequent secondary damage due to exposure of neural tissues [39 –41]. Repair in utero, for example, by tissue engineering techniques, can prevent this secondary damage caused by the exposure to the intrauterine environment (eg, the amniotic fluid). Several scaffolds have been used [5 –11] in tissue engineering approaches for treatment of spina bifida, but always without the addition of AFMCs. In a previous study, we showed that healthy AFMCs can be used for skin tissue engineering [32]. It might be possible to use these healthy AFMCs for allogeneic cell therapy to treat spina bifida, because the immature fetal immune system might be less susceptible to immune rejection. However, the only clinically successful in utero allogeneic stem cell therapy using hematopoietic stem cells has been carried out in severely immunologically compromised fetuses, suggesting that the fetal immune system plays an important role in the success of in utero stem cell therapy [42]. Therefore, autologous cell therapy is preferred. We stimulated NTD-AFMCs with TGF-β1 (a cytokine that stimulates collagen production), and compared the collagen type I deposition with that of H-AFMCs and HDF-f. Surprisingly, NTD-AFMCs did not show any collagen type I deposition, while both H-AFMCs and HDF-f showed collagen I deposition after TGF-β1 stimulation. These comparisons of collagen deposition were performed in monolayer cell cultures, under controlled TGF-β1 stimulation. Possibly, the results could have been different in vivo and/or under 3D conditions, both being more representative of the biological process. In like manner, other proteins of the TGF-β superfamily, other than TGF-β1, could have led to different results.
The etiology of spina bifida is multifactorial; both environmental and genetic factors play a role. So far, no mutations have been causally linked to spina bifida in humans. Folic acid deficiency due to nutrition problems is known to be associated with a higher incidence of spina bifida [43]. Furthermore, a number of pharmaceutical compounds can increase the risk of spina bifida [44,45]. The problem manifests itself already at the first month after fertilization. Since it is a highly localized defect, we expected that AFMCs derived from these fetuses are normal, and that they can be used as an autologous cell source for tissue repair purposes. However, much to our surprise, the NTD-AFMCs showed a markedly different expression of genes that are related to collagen synthesis compared with H-AFMCs and HDF-f. No higher expression of enzymes that degrade collagen was found in NTD-AFMCs, suggesting that collagen degradation cannot explain the lack of collagen type I deposition (although not all enzymes with collagenolytic activity were studied), but that there might be a defect in collagen synthesis and/or processing. Type I collagen is a heterotrimeric molecule composed of two α1 chains and one α2 chain. NTD-AFMCs did not express COL1A2. In addition, NTD-AFMCs had a much lower expression of genes (PCOLCE, PCOLCE2, ADAMTS2, and ADAMTS14) that are involved in the cleavage of propeptides from procollagen. Failure to remove the N- and C-terminal propeptides might result in a defective production of collagen fibrils. Despite the usage of AFMCs in a variety of studies, the current knowledge about the origin and function of these cells is limited. Therefore, it is hard to explain why NTD-AFMCs show such a different expression pattern of genes that are involved in collagen biosynthesis. It might be possible that the mesenchymal cells, which are isolated from fetuses with spina bifida, represent an abnormal subpopulation that does no deposit collagen I, while the healthy collagen-producing AFMCs possibly homed to the defect, as it has been shown that AFMCs populated fetal wounds and that the actual numbers of these cells can be affected by the presence of a NTD [46,47].
In spina bifida, two different types of tissue are affected; the nonclosure of the skin and the damaged neural tissue. In principle, both tissues can be repaired by the use of tissue engineering. Recently, in an experimental rat model for spina bifida [48], it has been shown that neural stem cells were present in the amniotic fluid derived from NTD rat fetuses, but not in the amniotic fluid from control fetuses. Although this has not been shown yet for humans, this could mean that even when NTD-AFMCs are not able to produce collagen (not suitable for skin tissue engineering, that is, closing the defect), these cells might still be used for tissue engineering to treat spina bifida to repair the neural tissue.
In summary, we have shown that NTD-AFMCs do not deposit collagen type I protein in vitro. This can be explained by a (much) lower expression of genes involved in collagen synthesis and processing of these cells compared with H-AFMCs and HDF-f. These results suggest that autologous fetal therapy as treatment for spina bifida to close the defect might not be possible.
Footnotes
Acknowledgments
Human amniocentesis cultures were kindly provided by Dr. R. Suijkerbuijk and J. van der Vlag (University Medical Center Groningen, the Netherlands). This work was supported by the EU-FP6 project EuroSTEC (soft tissue engineering for congenital birth defects in children; LSHB-CT-2006-037409). We thank Saskia de Rond for her excellent technical assistance.
Author Disclosure Statement
No competing financial interests exist.