
Abstract
Therapeutic approaches using multipotent mesenchymal stromal cells (MSCs) are advancing in regenerative medicine, transplantation, and autoimmune diseases. The mechanisms behind MSC immune modulation are still poorly understood and the prediction of the immune modulatory potential of single MSC preparations remains a major challenge for possible clinical applications. Here, we highlight galectin-9 (Gal-9) as a novel, important immune modulator expressed by MSCs, which is strongly upregulated upon activation of the cells by interferon-γ (IFN-γ). Further, we demonstrate that Gal-9 is a major mediator of the anti-proliferative and functional effects of MSCs not only on T cells but also on B cells. Here, Gal-9 and activated MSCs contribute to the suppression of antigen triggered immunoglobulin release. Moreover, we determined that Gal-9 expression could serve as a marker to predict a higher or lower immune modulatory potential of single cell preparations and therefore to distinguish the therapeutic potency of MSCs derived from different donors. Also in vivo co-administration of MSCs or murine Gal-9 resulted in significantly reduced IgG titers in mice immunized with human coagulation factor VIII (FVIII). In conclusion, Gal-9 acts as an immune modulator interfering with multiple cell types including B cells and Gal-9 may serve as a predictive indicator for clinical MSC therapy.
Introduction
M
Galectins are a β-galactoside-binding family that is expressed in various tissues [19]. These lectins form lattices on the cell surface [20] to interact with immune cells for example, T cells. These interactions may allow new insights into MSC versus T cell “communication.” Among the 15 known mammalian members, galectin-9 (Gal-9) is a 36 kDa tandem-repeat galectin, which can be found in immune cells, endothelial cells, or fibroblasts. It is a known inducer of T cell suppression and apoptosis [21]; these effects are mediated via the Tim-3 receptor or protein disulfide isomerases (PDI) [22,23]. In addition, Gal-9 expression is upregulated via IFN-γ stimulation in endothelial cells or fibroblasts [24,25]. In mice, Gal-9 was used to successfully treat GvHD in a bone marrow model [26].
Here, we identified Gal-9 as an important regulator of MSC immunosuppression. We could verify that Gal-9 is the only upregulated galectin in MSCs after activation with IFN-γ. Additionally, we introduce Gal-9 as a novel MSC related immune modulator not exclusively for T cells but more importantly for B cells. An in vivo model for alloimmune antibody formation in hemophilia A supports these findings, where activated MSCs and Gal-9 reduced the IgG response against FVIII in mice. Additionally, we introduce Gal-9 as a potential marker to distinguish between potent and less potent donor preparations.
Materials and Methods
Culture and analysis of MSCs
MSCs of different healthy donors under the age of 35 were derived from dispensable material (filters) of standard bone marrow harvests after informed consent and approvement of the local ethics committee. MSCs were isolated using standard protocols. In short, they were cultured in low glucose DMEM (1g/l; PAA) supplemented with 20% MSC qualified FCS (Invitrogen), 1% penicillin/streptomycin and 10 ng/mL hFGF (Peprotech). In short, MSCs were gained from dispensable materials of bone marrow sections. Bone marrow filters were flushed with DPBS and cells were separated by centrifugation. Isolation of MSCs was performed by plastic adherence. To maintain consistent and comparable experimental conditions MSC were used from passage 4 until 9. After passage 3, cells were analyzed for commonly known MSC characteristics. Differentiation and surface markers were performed as described before [17] and fluorescence activated cell sorting (FACS) data were analyzed via the Flowjo software (Stanford University).
After MSC characterization, four donors (MSC1-4) were randomly chosen for all further experiments. For cell culture experiments, 105 MSCs of different donors were seeded into six-well plates and grown until 80% confluence. In the presence of 20 ng/mL IFN-γ (Peprotech), MSCs were incubated for 24 h.
Lymphocyte isolation
Human peripheral blood mononuclear cells (PBMCs) were enriched from Buffy coats of healthy blood donors. PBMCs were isolated via Biocoll (Biochrome AG) separation. For each experiment, cells from a new donor were isolated.
For isolation of murine cells, C57Bl/6 mice were sacrificed and spleens were harvested.
Quantitative RT-PCR
Isolation of mRNA was performed according to the RNeasy-kit® protocol from Qiagen. mRNA translation into complementary DNA (cDNA) was performed by using the Applied Biosystems High-capacity cDNA reverse transcriptase kit protocol. cDNA was amplified by specific primers (Table 1, galectin primer sequences were adapted from [17]) using the Power SYBR Green master mix protocol (Applied Biosystems). β-actin was used as control. Probes were analyzed in an RT-PCR Step One Plus cycler (Applied Biosystems).
MSC transfection
Cells were nucleofected using the Amaxa Cell Line Optimization MSC Nucleofector Kit (Lonza). Transfection was performed according to the Amaxa manufacturing protocol with 5 μg Gal-9 cDNA (Thermo Fischer-Open Biosystems) or 5 μg His-tagged Gal-9 (GeneCopoeia). Control cells were transfected with 2 μg GFP coding pcDNA3.1 plasmid DNA.
Gal-9 analysis in MSCs
Fluorescence staining was performed as previously described [27]. (Transfected) MSCs were eventually mixed with immune cells, which were stimulated with 2 μg/mL PHA-P (Sigma-Aldrich) and 2 μg/mL LPS (Sigma-Aldrich) and incubated for 48 h. The anti-human Gal-9 Ab (Acris) was used as the primary antibody, followed by the appropriate Alexa-Fluor 488-coupled or Alexa-Fluor594-coupled secondary Ab (Invitrogen). His-tagged Gal-9 was detected by a anti-His Alexa Fluor 594 coupled Ab (MBL). As control, MSCs were stained only with the secondary antibody. Dapi (Santa Cruz Biotechnology) was used to stain nuclei.
For Gal-9 ELISA MSCs were lysed 24 h after stimulation with RIPA (Thermo Fischer) buffer. Supernatants were reduced in 10 kDa MWCO concentration tubes (Sartorius) 1:10. Gal-9 ELISA was arranged as described by MBL. In short, Anti-Gal-9 monoclonal antibody (ECA8; MBL) was coated on 96-well plates. Reduced supernatants or cell lysate samples were added and incubated overnight at 4°C. Recombinant Gal-9 (R&D Systems) served as standard. Then, samples were incubated with Anti-Gal-9 biotinylated (Abcam) followed by HRP-conjugated streptavidin (Biolegend). Gal-9 levels were detected via o-phenylendiamine (Roth) solution at 492 nm via an ELISA reader.
Proliferation assays
2×104 MSCs of different donors were seeded in 24-well plates or 6.5 mm transwells® with 0.4 μm pore size (Corning) and incubated with 20 ng/mL IFN-γ (Peprotech) for 24 h. Wells were thoroughly washed with phosphate buffered saline (PBS). For analysis of human cells either 105 PBMCs or T cells or B cells were added into the wells and incubated for 3 days. PBMCs and T cells were stimulated with 2 μg/mL PHA-P (Sigma-Aldrich) and 2 μg/mL LPS (Sigma-Aldrich). B cells were stimulated with 2 μg/mL CD40L (Abbiotec) and PHA-P (Sigma-Aldrich). Moreover, the influence of MSC expressed Gal-9 on immune cells was tested by adding 2 μg/mL of a Gal-9 blocking monoclonal antibody (ECA8; MBL). To determine the impact of Gal-9 on all three cell types, they were incubated with varying concentrations of recombinant Gal-9 (R&D Systems). Stimulated immune cells without MSCs were used as positive control. Immune cells were incubated with MSCs for 3 days and then thoroughly transferred into 96-well plates. To detect the proliferation rate, immune cells were mixed with BrdU and incubated for 22 h. BrdU incorporation was measured according to the Calbiochem®BrdU cell proliferation assay. Results were normalized for each single set of experiments.
ELISPOT
Ninety-six-well multiscreen immobilon-p PVDF membrane plates (Millipore) were coated with mouse anti-human κ chain specific antibody (SouthernBiotech) for 24 h at 4°C. Plates were thoroughly washed with PBS and MSCs of different donors were added to each well and incubated with or without 20 ng/mL IFNγ (Peprotech) for 24 h. Wells were thoroughly washed with PBS to reduce any residual IFN-γ. Then, 1:5 B cells were added and stimulated with 2 μg/mL CD40L (Sigma Aldrich) and 2 μg/mL VZV gE protein (Abcam). For blocking Gal-9, 25 mM of lactose were added. The cell mix was incubated for 6 days. Afterward, cells were washed off the plate and HRP conjugated anti-Human-IgG (SouthernBiotech) was added and incubated overnight at 4°C. Spots were developed with AEC-solution (Sigma-Aldrich). Results were calculated with the A.EL.VIS (Hannover) ELISPOT reader and software.
Western blot
Cell extracts and Supernatants of (IFN-γ activated) MSCs were prepared and processed at the indicated times. Probes were blotted via SDS-page on Biorad 4–10% gels. Immobilon-FL PVDF-membranes (Millipore) were incubated with a monoclonal rabbit anti-Gal-9 Ab (Abcam) in PBST. As secondary Ab, an anti-rabbit IRDey-700CW (LI-COR, Lincoln) was used and analyzed on an Odysee reader (LI-COR, Lincoln).
In vivo immune modulation
All animal procedures have been approved by the local animal care, protection, and use authorities (University and Regierungspräsidium Darmstadt). 8–10 week old female C57Bl/6 mice were simultaneously injected with 80U/kg recombinant Factor VIII (FVIII, Kogenate, Bayer) and followed by injection with either 5×105 activated MSCs or 3 μg murin Gal-9 (R&D Systems) or 3 μg polyclonal anti-Gal-9 antibody against murine Gal-9 (Acris) in PBS per mouse. Highest effective doses were adapted as previously described [26]. Mice were treated once a week. The treatment was repeated weekly for four times. 24 h after treatment and 7 days after the last treatment blood, samples (120 μL) were taken and analyzed for murine anti-FVIII IgG1 antibodies via ELISA. Then, mice were sacrificed and spleens were taken for the analysis of immune cells.
Fluorescence activated cell sorting
For analysis of mice immune cells, spleens were homogenized and investigated for B cells and T cells. Further, we investigated the distribution of CD8+ and CD4+(CD25+)-positive T cells. Moreover, CD4+CD25+ cells were segmented into Th1 (IFN-γ), Th2 (IL-4), and Treg–cells (FoxP3) by intracellular staining. For B-cell detection, we stained with CD19 and B220. Distribution of cells was then monitored with a BD FACSCantoII and analyzed with the Flowjo software. Anti-CD3e, anti-IFN-γ, anti-CD8a, anti-CD25, anti-IL-4, and anti-CD45R/B220 were all obtained from BD Pharmingen, anti-CD19 was from ABR, anti-CD4 was from Thermo Scientific, and anti-FoxP3 as well as the kit for fixation and permeabilization was purchased from eBioscience.
T-cell subset apoptosis assay
2*105 MSCs of nine different MSC donors were seeded into six-wells and incubated with or without IFN- γ for 24 h. Then, CD4+ T cells were added 1:5 and incubated for 72 h. T cells were stimulated with 2 μg PMA (Abcam) and ionomycin (Abcam). After 64 h, brefeldin (Abcam) was added 1:1000 to each well. T cells were then investigated for T-cell subsets such as TH1(CD3, CD4, IFN-γ), TH2(CD3, CD4, IL-4), and Treg(CD3, CD4, CD25, CD127, and Fox-P3) as well as annexin (all purchased from eBioscience) staining via FACS in separate preparations. T helper cells were first gated for CD3, then for CD4 until the gate for the intracellular staining was applied. Tregs were subsequently gated for CD3 and CD4 positivity, then CD4 and CD25 high. The last gate excluded CD127+ cells to validate the Fox-P3-positive cells.
Anti-FVIII ELISA
For murine anti-FVIII ELISA plasma was isolated from murine blood samples and ELISA was performed as described before [28].
Statistics
Student's t-test or ANOVA with the Dunnett test or Tukey test for multiple comparisons were used to validate significance of the results. Data were analyzed with the InStad Version 3.06 software. Data were considered as significant when P<0.05.
Results
IFN-γ and B cells enhance Gal-9 expression in MSCs
To investigate whether there are donor-specific differences between MSC populations, we focused on potential markers of MSC potency and established an mRNA profile. Chosen markers consisted of several chemokine receptors, as demonstrated by Ciuculescu et al. [29] and galectins. At first we established and characterized MSC preparations of four different donors for further investigation (Supplementary Fig. S1A, B; Supplementary Data are available online at
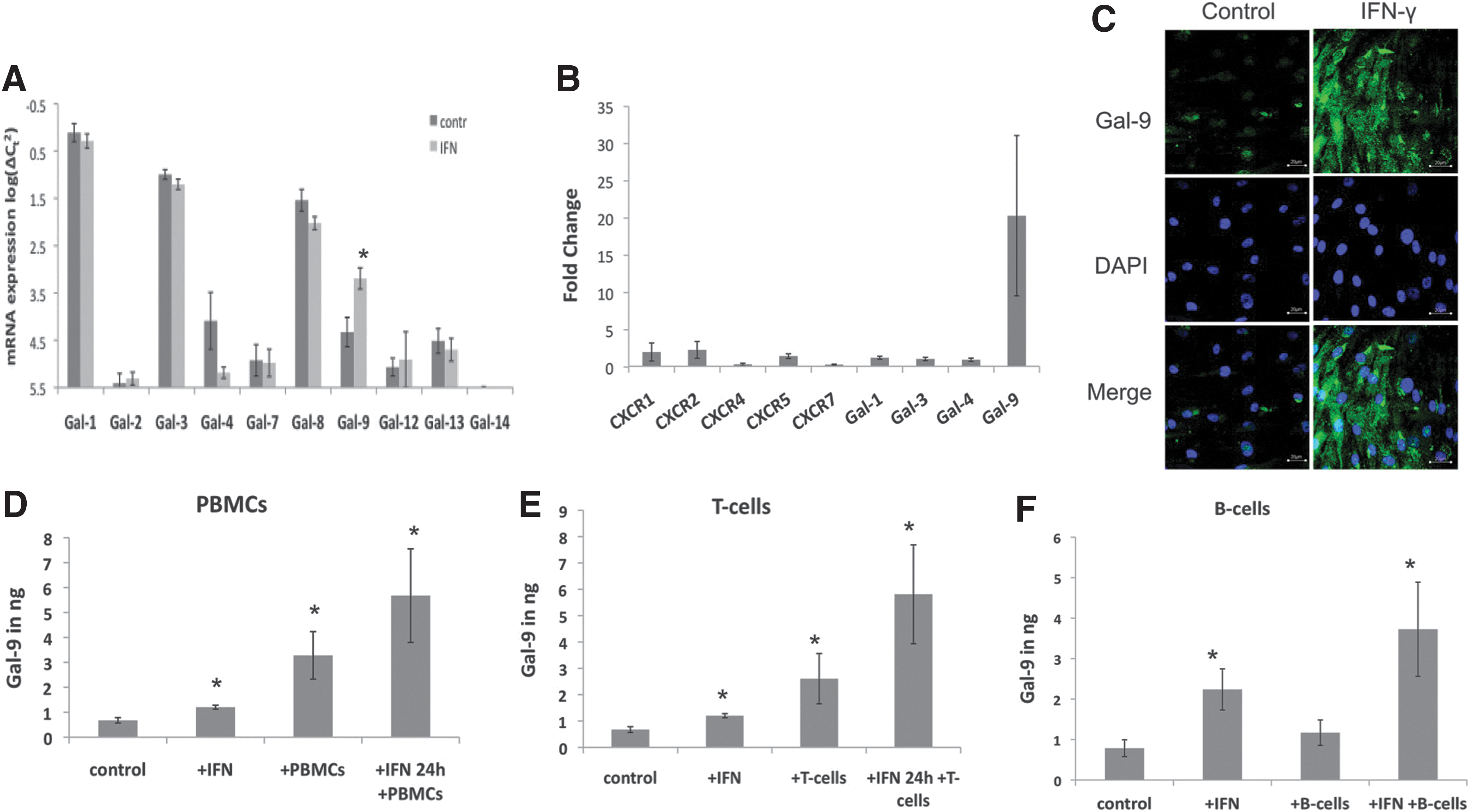
Galectin-9 (Gal-9) as an enhanced immunomodulatory agent in interferon-γ (IFN-γ)-activated mesenchymal stromal cells (MSCs).
Gal-9 is responsible for B-cell modulation
Because of the prominent Gal-9 upregulation in activated MSCs, we first focused on the functional role of Gal-9. Therefore, we added different concentrations of the recombinant protein to PBMCs, T cells, and B cells and performed proliferation assays. In agreement with published results [31], Gal-9 reduced the proliferation of PBMCs (Fig. 2A) and T cells (Fig. 2B) at increasing concentration. Rather unexpected, we observed a comparable effect on the proliferation of B cells, which resulted in the same extent as for T cells (Fig. 2C). Therefore, we wondered whether Gal-9 is not only a suppressor of B-cell proliferation but also might influence the functionality of B cells, for example, antibody release. We consequently performed ELISPOT assays to demonstrate that Gal-9 could reduce varicella zoster virus (VZV) triggered IgG release as depicted in figure 2D. In conclusion, Gal-9 decreases B- and T-cell proliferation and, more importantly, B-cell activation.
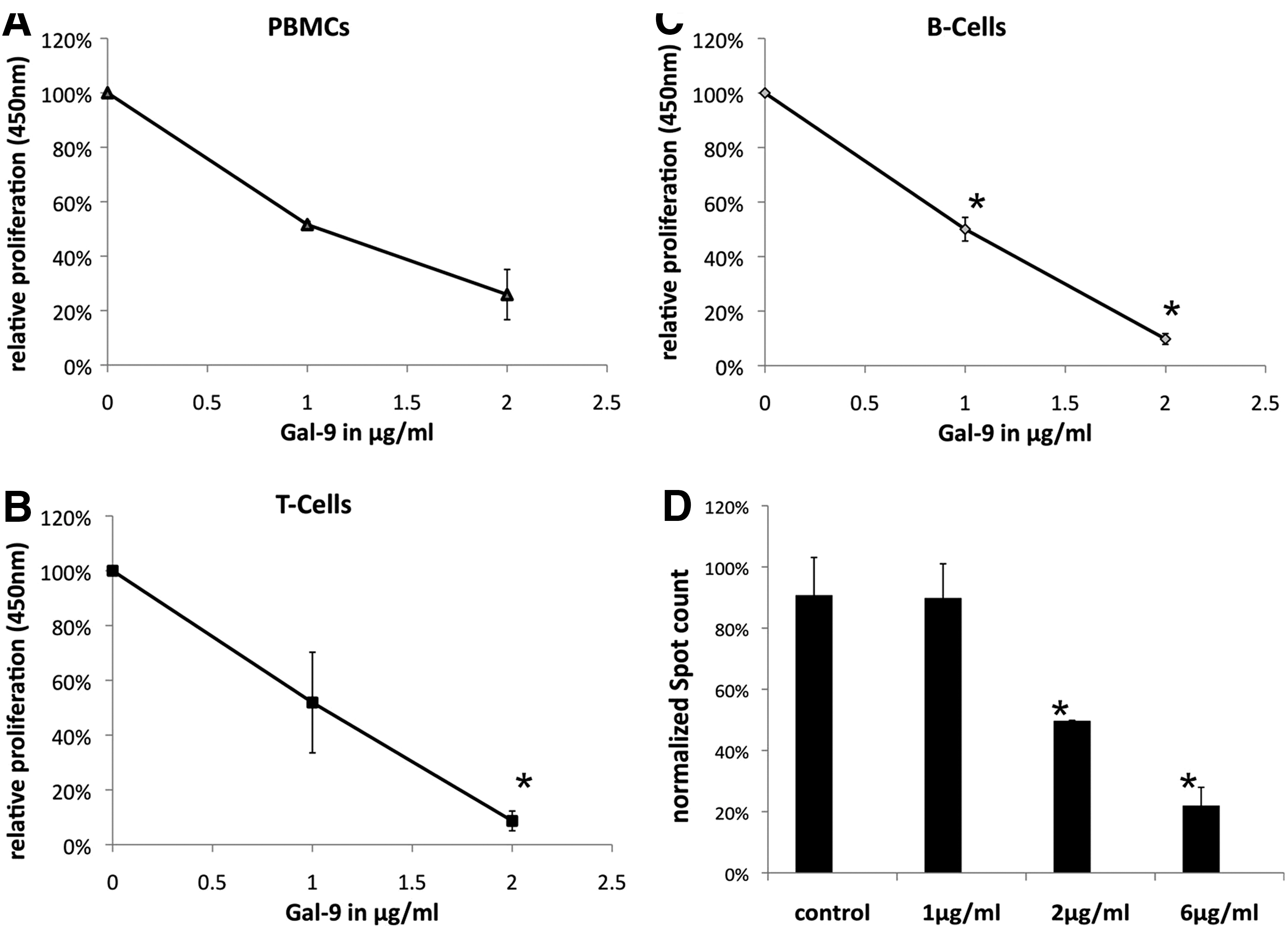
Proliferation of immune cells and antibody formation is inhibited by Gal-9. Stimulated PBMCs
Galectin-9 is an immune modulator of activated MSCs
To determine the importance of Gal-9 among the multiple immune modulatory mechanisms of activated MSCs, we mixed stimulated T or B cells of several donors with either untreated or preactivated MSCs and monitored the cell proliferation of stimulated immune cells. Unstimulated immune cells served as negative control (not shown). Reduction of T-cell proliferation by IFN-γ activated MSCs exceeded the effect of nonactivated MSCs. As demonstrated for T cells the activation of MSCs achieve a stronger suppressive effect on B-cell proliferation. As expected, T-cell proliferation could be restored almost completely after blocking Gal-9 in activated MSCs with a monoclonal anti-Gal-9 antibody (Fig. 3A). Surprisingly, B-cell proliferation was elevated to normalized levels by blocking Gal-9, when coincubated with naïve MSCs but did not reach normalized levels when MSCs were activated previously (Fig. 3B). Because Gal-9 co-incubation reduced the IgG release of VZV triggered B cells, we also wanted to investigate the influence of activated MSCs. As Figure 4C depicts, nonactivated MSCs had no influence on IgG release. In contrast, activation of MSCs with IFN-γ decreased the IgG release by around 50% compared with the control group. As expected, addition of lactose, a previously described natural inhibitor of Gal-9, to block Gal-9 activity, abolished the Gal-9 suppressive effect of activated MSCs to a high extent.
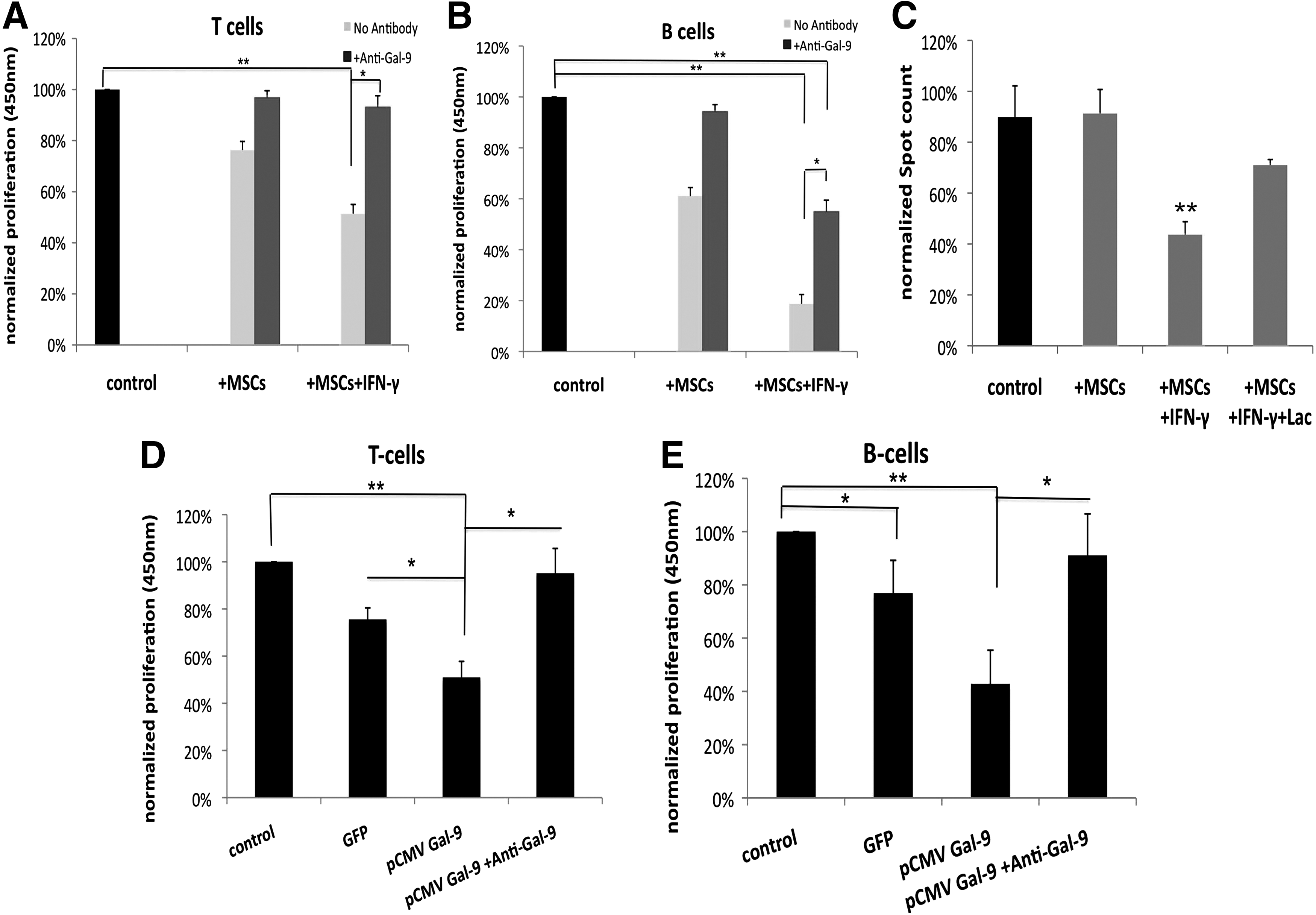
Gal-9 is a mediator of immune suppression by IFN-γ-activated MSCs. Stimulated T cells
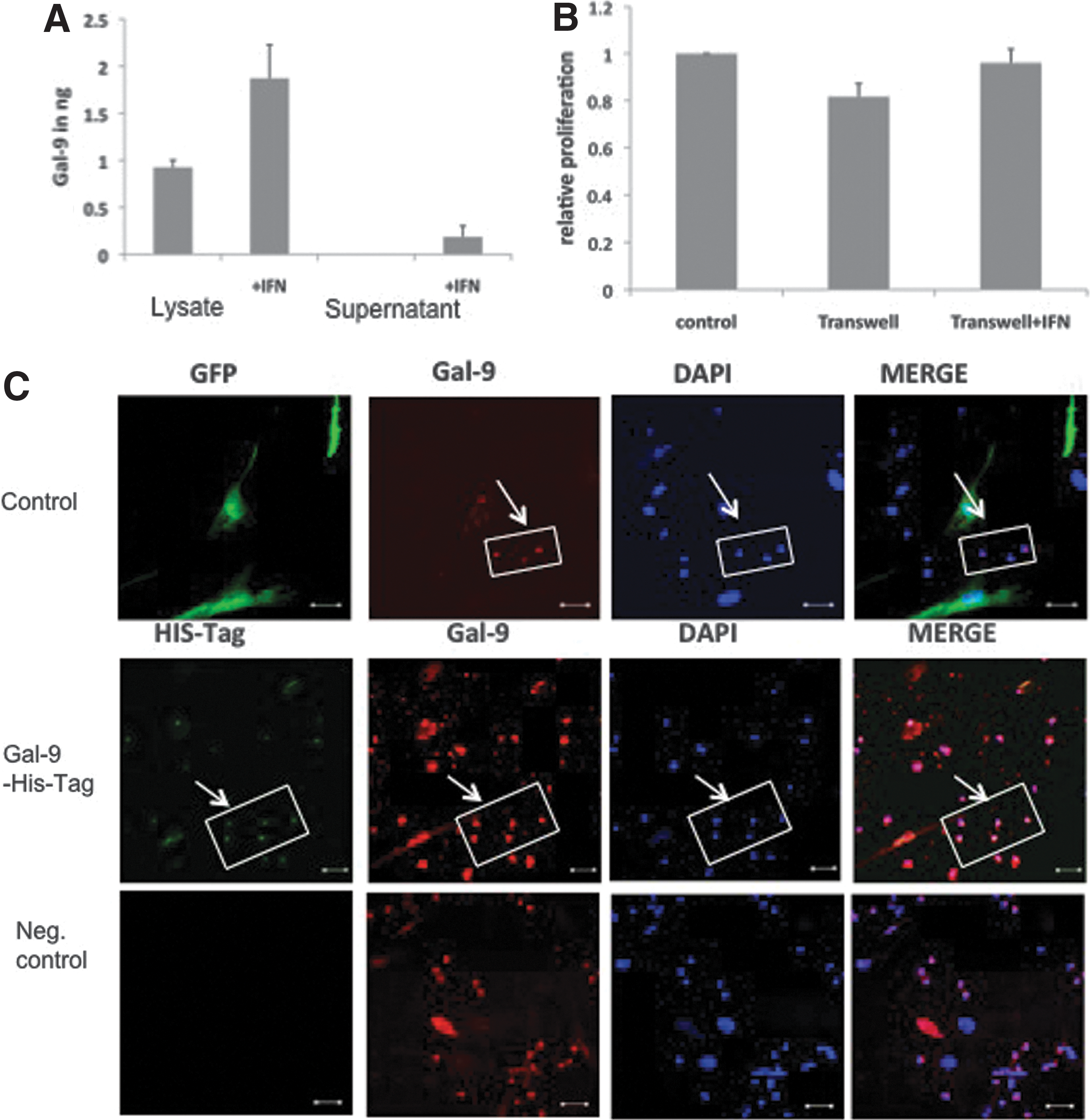
MSCs mediate Gal-9 immune modulation through cell–cell contacts.
To confirm that Gal-9 mediates MSC-related immune suppression, we overexpressed the protein in MSCs and measured inhibition of T- and B-cell proliferation. Transfection with a GFP-expressing construct was used as transfection and negative control. The MSC transfection efficacy was around 70%. Successful transfection of MSCs with the Gal-9 encoding pCMV plasmid was monitored via mRNA isolation and SYBR Green RT-PCR. We detected a 6000-fold increase of Gal-9 mRNA (Supplementary Fig. S2A). Gal-9 overexpressing MSCs were then employed in T cell (Fig. 3D) and B cell (Fig. 3E) proliferation assays. MSCs were mixed at a 1:5 ratio with immune cells and incubated for 3 days. Gal-9 overexpression resulted in a significant higher inhibition of B cell proliferation compared with GFP-transfected control MSCs. This Gal-9 mediated anti-proliferative effect on T and B cells was completely abolished by blocking Gal-9 with a monoclonal antibody.
Immune modulation of MSCs via Gal-9 is mediated by cell–cell contacts
Since we could clearly demonstrate the Gal-9 dependent immune modulation of activated MSCs, we further wanted to determine whether these effects are based on cell–cell contacts or by secreted Gal-9. Therefore, supernatants and lysates of MSCs were investigated for Gal-9 via ELISA (Fig. 4A) and western blot (Supplementary Fig. S2B). In contrast to supernatants of untreated MSCs, only lysates of naïve MSCs (1 ng) showed good detectable levels of Gal-9. Activation increased the levels in lysates (2 ng) and minor amounts of Gal-9 were detected in concentrated supernatants (0.3 ng). Therefore, we performed a proliferation assay where we physically separated MSCs from T cells (Fig. 4B). Interestingly, we could not detect a significant decrease in proliferation when MSCs were separated by a semipermeable transwell. Further, the addition of Gal-9 overexpressing MSC supernatants had no influence on immune cell proliferation. To enlighten the immune modulatory mechanism we overexpressed a His-tagged Gal-9 in MSCs and mixed them with PBMCs. After 24 h, immune cells were clustered onto the MSCs (Supplementary Fig. S2C) and immunofluorescence staining depicted that detectable amounts of His-tag-Gal-9 were traceable in immune cells, which are in close proximity to successful transfected MSCs (Fig. 4C).
Galectin-9 is upregulated donor dependent
Although we depicted Gal-9 as an immune suppressor in MSCs, the question still remains whether there are donor-dependent differences in Gal-9 expression and whether Gal-9 might serve as a marker for MSC potency with respect to their immune modulation. Initially, randomly selected donors were compared for their Gal-9 expression at mRNA and protein level. Similar Gal-9 protein levels were detected in all donors by ELISA (Fig. 5A). As immune modulation of T and B cells was enhanced by IFN-γ, we also investigated Gal-9 levels of each donor after activation. Donor MSC4 (black box) showed decreased protein and mRNA levels upon stimulation (Fig. 5A, B). The expression pattern did not change over subsequent passages of MSCs, for example, from passage 4–7. Consequently, we performed proliferation assays with these donors. Again, nonstimulated MSCs showed minor differences in their immune modulative potential regarding PBMCs, T and, most importantly, B cells. Interestingly, MSC stimulation with IFN-γ resulted in a significant decrease of PBMC, T, and B-cell proliferation in all donors except MSC4 (Black box, Fig. 5C).

Immune suppression of IFN-γ activated MSCs is donor and Gal-9 increase dependent.
Here, we distinguished between donors according to their Gal-9 expression after activation. To confirm these initial results, we broadened our investigation and included nine additional characterized donors (Supplementary Fig. S3A). We measured changes of Gal-9 protein level after activation with IFN-γ. As depicted in Fig. 5D, MSC donor 9 (MSC9) did not exhibit any increase of Gal-9 in response to stimulation. As seen with donor MSC4, this donor did not decrease B cell proliferation after MSC activation (Fig. 5E). Because Gal-9 is a know modulator of TH1 cells we investigated CD4-cell subsets. TH1 cell counts are decreased by Gal-9 and co-incubation with most MSCs donors. These suppressive effects were much more enhanced by MSC activation. As predicted, Treg cell counts were slightly enhanced by MSC co-incubation. Interestingly, Treg cell counts were decreased by activated MSCs below control levels and by Gal-9. As predicted, donor MSC9 did not influence TH1 and Treg cell counts after IFN-γ activation compared to the naïve state (Fig. 5F, G). Apoptosis rates of CD4+-cells remained unchanged after activation (Supplementary Fig. S3C). As expected Gal-9 or MSC activation had no effect on TH2 cell counts.
MSCs and Gal-9 inhibit antibody formation in mice
Because we identified Gal-9 as a regulator of human T and B cells, we strived to investigate the effect of Gal-9 and activated MSCs in vivo. As disease model with a determined antigen, we chose antibody formation against FVIII. We immunized mice with human FVIII and tested the effect of murine Gal-9 (mGal-9), a polyclonal mGal-9 inhibiting antibody, and IFN-γ-activated human MSCs. In mice, which were treated with MSCs in addition to FVIII immunization, we monitored a significant reduction of anti-FVIII-IgG formation. Also, mGal-9 co-treated animals developed reduced antibody titers against FVIII compared with blocking of Gal-9 or FVIII alone. Anti-FVIII titers rapidly increased after 7 days and exceeded the impact of immunized-only mice many fold (Fig. 6A). Only MSCs stabilized the equilibrium of T and B cells in spleen cells after spleen sections, whereas Gal-9 co-treatment, and Gal-9 blocking, resulted in elevated B cell counts (Fig. 6B). Further, primarily mGal-9 decreased Th1, TH2, and Treg cell counts. MSCs application, in contrast, significantly increased Th1, TH2, and Treg cell counts. Blocking Gal-9 primarily increased TH2 cells (Fig. 6C).
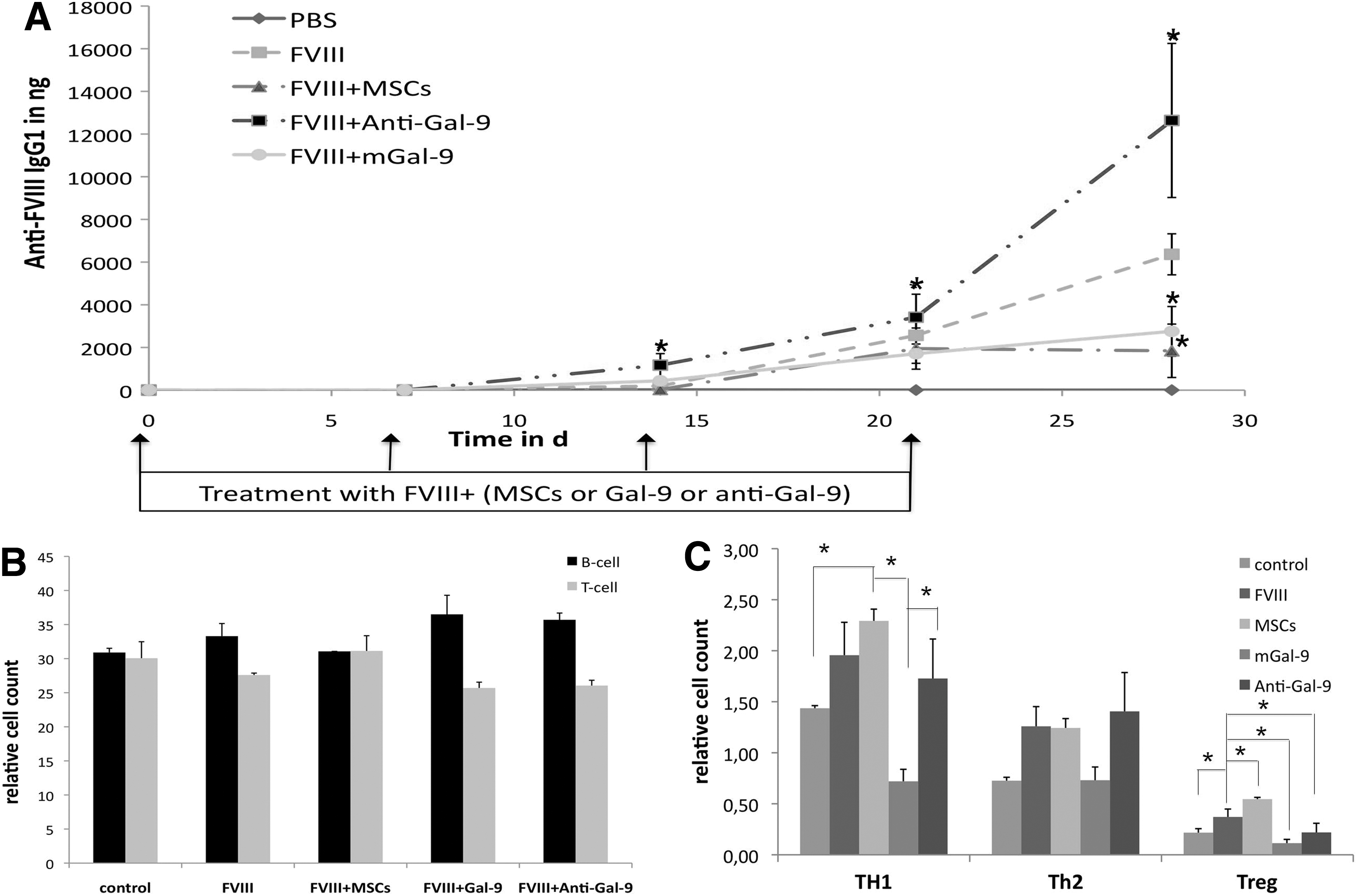
MSCs and Gal-9 inhibit anti-FVIII antibody response in vivo.
Discussion
The mechanisms behind MSC-mediated immune modulation remain controversial and most likely are the result of multifactorial pathways [32]. Known contributing factors include PGE2 [33], HGF [13], IDO [15], or TGF-β [13], which are connected with T cell or dendritic cell function. Recently, Gieseke et. al. [17] introduced Gal-1 as another immunomodulator expressed by MSCs, moving the group of immune regulatory galectins into focus. Shortly after, the role of galectin-3 was analyzed for its potential to suppress PBMC proliferation upon TLR activation [18]. Coinciding with our observations Gieseke et al. demonstrated the inducebility of Gal-9 in MSCs by pro-inflammatory stimuli [30] and delivered a possible explanation how MSCs might improve tissue regeneration and influence inflammatory tissue environment after transplantation [34]. Here, we confirmed these results for pooled MSC populations and added B cells as a new target to the immune modulation with Gal-9. We confirmed that exclusively Gal-9 is strongly increased following stimulation with IFN-γ and other pro-inflammatory stimuli. Further, not only co-incubation with T cells but, more importantly, with B cells increased Gal-9 expression in activated MSCs, which indicates a crosstalk between these cells. MSCs inhibited T and B cells without IFN-γ activation but the efficacy can be substantially enhanced by previous activation with IFN-γ [35,36]. Moreover, a very low MSC-T cell ratio of 1:2 or 1:3 is commonly used in most in vitro experiments to achieve strong effects [33] and the immune suppressive potency decreases with declining ratios [30]. The time-dependent upregulation of Gal-9 by IFN-γ had been described for endothelial cells. In the endothelium, Gal-9 acts as a chemoattractant for eosinophils and increased adhesion [25]. This process is supported by HDAC3, which induced the interaction of phosphoinositol 3-kinase (PI3K) and IFN response factor 3(IRF3), resulting in phosphorylation of IRF3, its nuclear translocation, and increased Gal-9 expression [37]. Whether this signaling cascade is responsible for enhanced Gal-9 expression in MSCs as well, still has to be elucidated. Nevertheless, activation of MSCs seems to be crucial for enhanced immunosuppression as it was described for the successful treatment of GvHD in mice [38]. Additionally, only activated MSCs have an elevated potential for the regulation of human PBMCs and T cells, but their immune modulatory potency depends on the cytokine milieu that is provided by stimulated immune cells [39]. The suppressive mechanism involved after IFN-γ stimulation is still controversial; exemplarily by studies on the role of IDO, which is upregulated after activation and immune cell proliferation can be partially restored by an antagonist [16]. On the other hand, MSCs can inhibit immune cell proliferation independently of IFN-γR1 and, moreover, independent of IDO [40]. In agreement with these findings, we demonstrated that the induction of Gal-9 in MSCs led to an enhanced immunosuppression of stimulated PBMCs, T cells, and B cells. Interestingly, T and B cells seem to increase Gal-9 levels in MSCs by different pathways. Stimulated T cells and PBMCs might increase Gal-9 levels by pro-inflammatory mediators [30], whereas B cells only significantly enhanced Gal-9 levels when MSCs were activated before.
Because soluble factors and cell–cell contacts are believed to be responsible for these actions [41], we could depict that MSC-derived Gal-9 can be found in immune cells which are in close proximity. Further, to modulate immune cells, very high level of recombinant Gal-9 are necessary compared to levels measured in MSC preparations. A possible explanation could be the N-glycan-binding character of galectins. They can bind a variety of surface glycoproteins, which do not entirely contribute to cell viability [20,42]. Therefore, we identified cell–cell contacts as the most probable scenario for MSC/Gal-9 immune modulation. Here, MSCs could directly influence T or B cells by Gal-9 transfer and interaction with glycan proteins. Blocking of endogenous or overexpressed Gal-9 restored the proliferation of T and B cells, which confirmed our hypothesis. Gal-9 is a known inducer of apoptosis of CD4+ and CD8+ T cells via the calcium-calpain-caspase-1 pathway [43]. More precisely, it was demonstrated that apoptosis of Th1-cells is induced via the Tim-3/Gal-9 pathway [44]. As predicted, Gal-9 does not inhibit Th2 cell counts. This is due to different glycosylation patterns of membranes of responsive cells. Th2 cells express significant amounts of α-2,6-linked sialic acid, which blocks Gal-9 binding to glycan receptors such as those required to induce cell death [31]. Additionally, T cells express the Gal-9 binding PDI on their surface, which promotes apoptosis of Th1 but induces the migration of Th2 cells [23]. Here, we demonstrated that co-incubation with Gal-9 or activated MSCs results in decreased cell counts of TH1 but not in Th2 cells. For Treg cells the described effects are rather different. As described in various in vitro and in vivo models MSCs increased Treg levels via Gal-9 independent mechanisms such as TGF-β or PGE2 [45,46]. Surprisingly, Gal-9 and activated MSCs restored or even decreased Treg cell counts. Because apoptosis rates did not differ between naïve and stimulated MSCs, Gal-9 appear to mediate its effect via altering functional pathways. In general, Gal-9 may bind to more than nine cell surface receptors [23,31] and might influence intracellular molecules as well. These multipotent binding properties of Gal-9 have been assigned to its two N- and C- terminal carbohydrate recognition domains (CRD). This bi-domain character of Gal-9 enables it to mediate different functions in innate and adaptive immunity [47]. In this study, we describe a formerly unknown property of Gal-9, which is the influence of Gal-9 on B cell proliferation and IgG release in vitro and in vivo. Although, no interaction partner has been described, CD45 or CD44 could serve as receptor for Gal-9 binding on B cells [23,48], whereas CD44 is upregulated in stimulated B cells [49]. Moreover, we could demonstrate that specifically Gal-9 is one important regulator in MSC-mediated immune modulation of human B cells (eg, proliferation or IgG release). For MSCs, it has been demonstrated that they inhibit B cell proliferation and IgG, IgM, or IgA release [50]. Moreover, IgG-release is believed to depend on the concentration of the antigen. Only at high antigen concentrations, mixed B cell/MSC cultures resulted in a diminished IgG production [51]. Herein, we demonstrated a decrease of IgG production at comparable low antigen concentrations by activating MSCs, which we definitely could attribute to MSC-expressed Gal-9. Although the suppression of terminal B cell differentiation and IgG1 and IgM release had been assigned to MSCs, the mechanisms behind were still unclear [52]. Also, clearly increased Gal-9 levels seem to be essential to maximize the immunomodulatory potential of MSCs not only on T cells but also on B cell.
For clinical applications, it is essential to distinguish between “good” or “bad” human MSC donors for immune modulation [53]. Our detailed investigation, for the first time, identifies Gal-9 as a potential marker for single donor preparations. In vitro, MSCs that express high changes in Gal-9 mRNA and protein levels after IFN-γ activation tend to be more potent suppressors of TH1 and Treg cells and B cells. We, so far, identified two donors (MSC4 and MSC9), which had very low effects on immune cell proliferation and presented only minimal Gal-9 response. General differences between donors may be connected with a different sensitivity of all these immune subtypes. To further validate Gal-9 as a new MSC-driven immune regulator, we extended our investigation and analyzed MSCs and Gal-9 in vivo in a disease model of allo immunization. Immunization against FVIII is the most frequent complication in hemophilia A treatment and FVIII triggers a profound IgG response in mice [54]. Because Rafei et al. [55] demonstrated that MSC application support the clearance of anti-FVIII titers in mice, we wanted to know whether activated MSCs were capable of suppressing FVIII immune response simultaneously. As expected, activated MSCs prolonged the antibody-free time and suppressed high titers in mice. Interestingly, blocking mGal-9 boosted the IgG response, but coadministration of mGal-9 significantly reduced IgG1 development. These findings are supported by recent publications, where Gal-9 reduced anti-dsDNA antibodies and induced TIM-3 independent apoptosis of plasma cells in a systemic lupus erythematosus model [56]. Unexpectedly, MSCs did not influence Th2 cell counts but boosted Treg cells, whereas mGal-9, in contrast to an inflammation model [57], reduced the number of all relevant immune cells. In this context, we assume that Gal-9 might be a potent regulator for (auto/allo)-immune diseases but the murine system might only deliver limited relevance to what happens in human. Interestingly, Gal-9 has been demonstrated to support the suppression of allergic symptoms [58,59] in vivo. At the time of this study, there are 15 active clinical trials running for the MSC-based treatment of autoimmune diseases such as type 1 diabetes, multiple sclerosis, or lupus nephritis, listed at
Footnotes
Acknowledgments
The authors wish to thank Halvard Bönig at the DRK for review of the article. Research was supported by the “Stiftung Hämotherapie-Forschung” by the German Red Cross Blood Donor Service Baden-Württemberg-Hessen, by a grant from the GRK-1172 and by a Bayer Hemophilia Award from Bayer HealthCare to J.S., C.U., and P.QL., who are students within the graduate study program GRK-1172 funded by the DFG, and E.S. received funding through ECCPS (DFG). E.S., and J.S. received support from the LOEWE Center for Cell and Gene Therapy, Frankfurt, funded by the Hessisches Ministerium für Wissenschaft und Kunst (HMWK), funding reference number: III L 4-518/17.004 (2010).
Poster presentation: 1. Perspectives in Cell and Gene-Based Medicines (CGT), Immune Suppression by Mesenchymal Stromal Cells (MSCs) is Mediated via Galectin-9, Frankfurt 2012.
2. Galectin-9 is a Suppressor of T- and B-cells and Predicts the Immune Modulatory Potential of Mesenchymal Stromal Cells, Ash, Atlanta 2012.
3. The Donor Dependent Immune Modulatory Potential of Mesenchymal Stromal Cell Preparations Can be Predicted by Galectin-9, EHA, Stockholm 2013.
Author Disclosure Statement
C.U., and J.S. are currently employed at Biotest AG. J.S. was supported via a Bayer Hemophilia Award.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.