
Abstract
Although tumor angiogenesis in relation to cancer therapy has been widely investigated for more than four decades, its counterpart tumor hematopoiesis has not been equally considered. In that respect, in our long-term immunohistochemical examination of fetal hemoglobin (HbF) cells in various solid tumors, we have observed signs of fetal hematopoiesis in situ within the tumors. We hypothesize that this observed fetal hematopoiesis, involving angiogenesis, mirrors mammalian blood system development in the embryo and the fetus; this is consistent with the concept of the hemogenic endothelial progenitor, common to endothelial and hemopoietic cells. Based on this assumption, there should exist in tumors at least two routes of hematoangiogenesis: one of fetal (HbF) hematopoiesis and the other of adult (HbA) hematopoiesis, each one deserving a different therapeutic approach. In the fetal route, HbF should support tumor growth by virtue of its high oxygen affinity.
Introduction
I
Primitive and Definitive Erythroblasts in Ontogenetic Blood Development
The first “wave” of blood development in avian and mammalian embryos takes place extraembryonally in the yolk sac as “blood islands” consisting of clusters of nucleated primitive (indefinitive) erythroid cells, encircled by primordial endothelium [13]. At this stage, hematopoiesis is restricted to the primitive, erythroid lineage [14,15], exclusively producing embryonic (epsilon chain) and a small quantity of fetal (gamma chain) hemoglobins [5,16]. This lineage of primitive nucleated erythroid cells supplies the embryonic circulation where they eventually mature into enucleated erythrocytes; they are subsequently replaced by a second wave of definitive erythroid cells maturing intraembryonally in which a switch takes place from embryonic hemoglobin into HbF [17]. Such a discrimination between primitive and definitive erythroid cells was recapitulated in vitro in cultures of human embryonic cell lines [5]. In these cell lines, a first wave of hematopoiesis was predominated by primitive erythroid cells, expressing glycophorin A, and exclusively embryonic hemoglobin with a small amount of HbF. This was followed by a second wave of hematopoiesis of definitive erythroid cells largely expressing HbF, in addition to granulocytes-macrophage progenitors and multilineage precursors. In a few of our tumor specimens, we observed in the mesenchyme adjacent to the tumor tissue, clusters, predominantly consisting of nucleated erythroid cells, with irregular cell boundaries, positively stained for HbF. These cells found within sinuses and loosely surrounded by epithelial cells (Fig. 1) might resemble the primitive erythroid cells of the yolk sac as previously described (Figs. 1 and 2 in Ferkowicz and Yoder [18]). However, in most of our tumor cases, we found much stronger stained HbF-nucleated erythroid cells, intermingled with their HbF-erythrocyte descendants (Fig. 2a, b). These cells resemble the definitive erythroid cells inside the embryo. Yet, in order to clearly distinguish between primitive and definitive erythroid cells in the tumors, we still need to comparatively stain those cells for embryonic hemoglobin and HbF, and expect HbF to be the predominant hemoglobin type in the definitive erythroid cells.
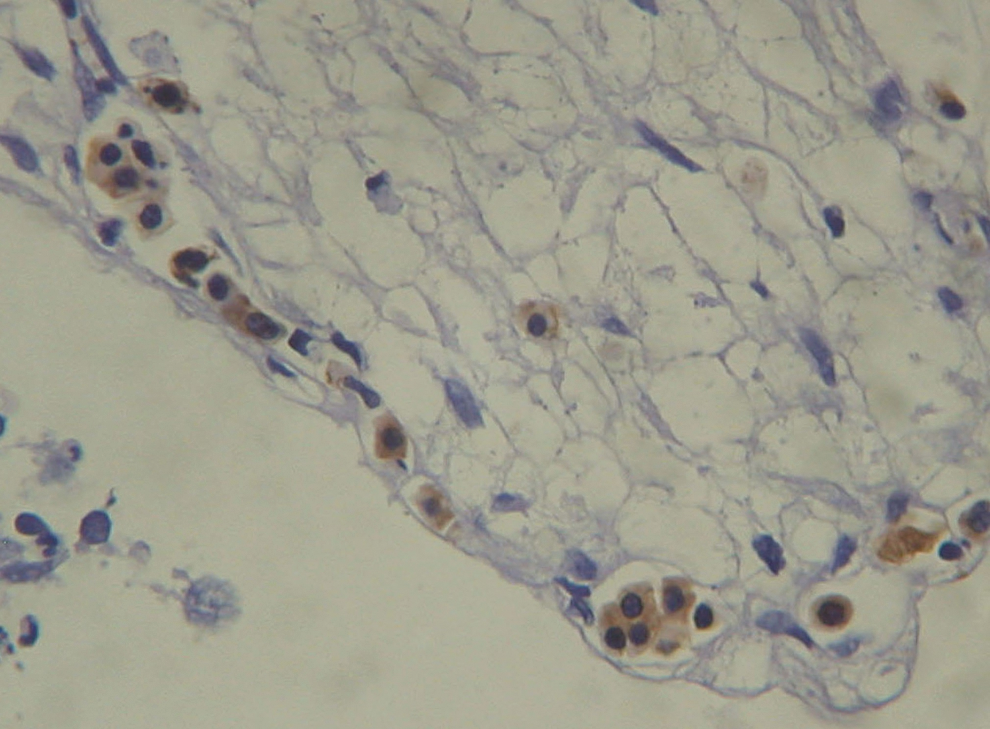
Clusters of nucleated fetal hemoglobin (HbF) cells, resembling the blood islands of the yolk sac primitive erythroblasts, in the mesenchyme, around tumor cells of the uterus. Color images available online at

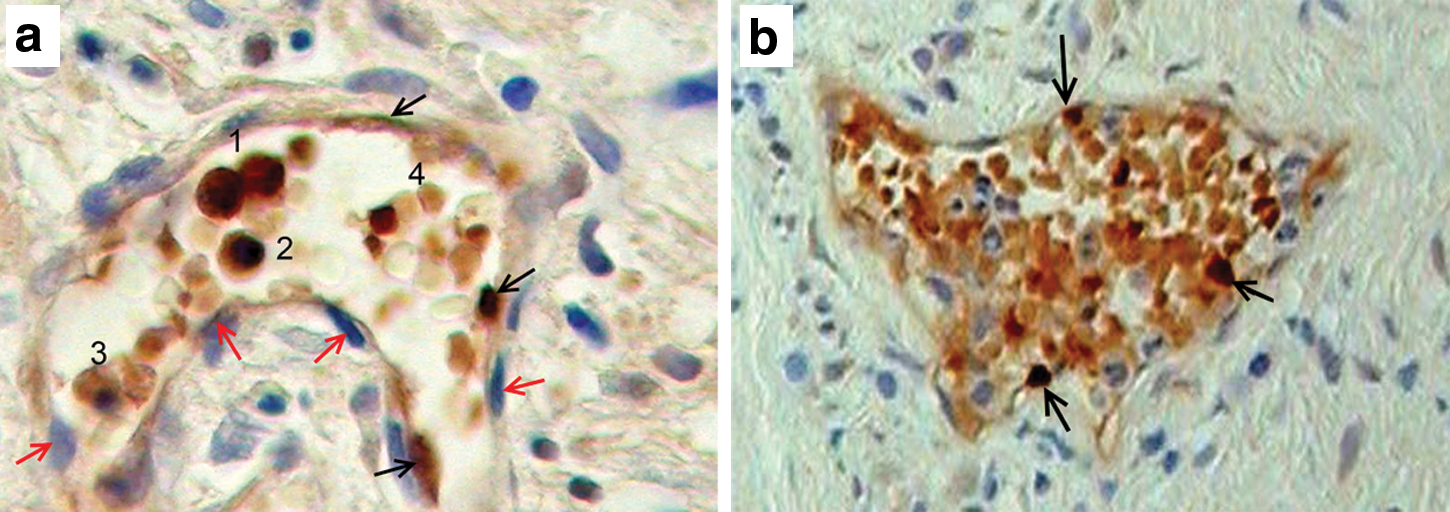
Proliferating HbF erythroid cells as a mixture of erythrocytes and their nucleated precursors.
Possible Endothelial Origin of Prenatal Hematopoiesis
There is experimental evidence from the mouse and avian embryos that suggests that at the stage of primitive erythropoiesis, erythroid and endothelial cells do not have a common progenitor; rather, they develop from separate precursors since in the yolk sac of the mouse embryo [19] during its gastrulation, the emergence of erythropoietic precursors precede the development of the yolk sac endothelium. Similarly, in the avian embryo [14], cells from the early yolk sac produced in vitro either endothelial or hematopoietic colonies, but not mixed ones. However, in an experimental model of cultured human embryonic cells [7], there was an indication of a common origin for endothelial and primitive erythroid cells within mesodermal epitheloid cells with endothelial gene expression (VE-cadherin+) (Table 1); these epitheloid cells differentiated into endothelial cells, while on the other hand, they could be transformed in response to the “fibroblast growth factor 2” into primitive erythroid cells (VE-cadherin−) expressing human embryonic hemoglobin (epsilon chain) with no adult hemoglobin (HbA, beta chain).
VEGFR, vascular endothelial growth factor receptor.
The vascular origin of hematopoiesis was first indicated as budding of hematopoietic cells from vascular endothelial cells as originally described in 1907 in the dorsal aorta of the aorta-gonad-mesonephros (AGM) region of the avian embryo [20]. Similarly, in the dorsal aorta of the human embryo (AGM region), such CD45+ (Table 1) hematopoietic stem cells (HSCs) were immunohistochemically observed, bulging from the ventral wall of the vascular endothelium [21]. Those CD45+ HSCs shared CD34 expression (Table 1) with the overall endothelial lining, indicating their endothelial origin. Although those CDs are not erythropoiesis specific, they could be related to erythroid development since CD45 [22] and CD34 [23] are expressed by embryonic erythroid precursors. However, in order to finally determine the vascular origin of prenatal erythropoiesis, we shall need to use erythroid-specific markers, like glycophorin-A and fetal or embryonic hemoglobin subsequent to the endothelial cell markers. Our question, however, is whether such vascular erythropoiesis involving HbF does take place in situ of the tumors. We have found such an indication in many of our immunohistochemical examinations [3], wherein we were able to observe HbF-stained cells in the endothelium of blood vessels (Fig. 3a, b), inside of which HbF erythroid cells were the predominant cell type, possibly originating in the endothelium from which they were shed into the vessel.
Comparing Blood Formation in Postnatal Life and in Tumor to the Prenatal State
As we have learned, hemogenic endothelial cells take a central role in the embryo as progenitors of definite erythroid cells [5 –7]. However, such hemogenic endothelial cells have not yet been detected in adult bone marrow hematopoiesis, which is, presumably, made mainly by HSC renewal [24]. In the murine model, adult HSCs do express the same phenotype as the embryonic HSCs, namely, CD31+ [25] (Table 1), CD45+, cKit+, and vascular endothelial growth factor receptor 2 (VEGFR2) (Flk-1) (Table 1). Nevertheless, hemogenic endothelial progenitors of such mature HSCs expressing VE-cadherin, VEGFR2, and not expressing CD45 were isolated from the embryo, although they could not be isolated from the adult bone marrow [26]. This indicates a difference in the developmental routes of embryonic and adult HSCs. Another difference in the blood-forming system between prenatal and postnatal states resides in the changes of hemoglobin composition from embryonic hemoglobin and HbF to HbA [27]. Recapitulation of the prenatal state has been immunohistochemically demonstrated by observing HbF cells inside tumors, including colorectal tumors [1], embryonal tumors [2], leukemias [28], urinary bladder tumors [3], and others. A quantitative analysis performed in the urinary bladder tumors [3] revealed that in 76% of patients with low-grade tumors, over 50% of blood vessels predominantly or exclusively contained HbF cells.
The involvement of hematopoiesis with angiogenesis as mediated by a common origin of hemogenic endothelial cells has not been hitherto considered as taking part in normal or in tumor in situ, blood forming, although tumor angiogenesis has been investigated for over three decades [8 –11]. Tumor blood forming was found to be different from normal angiogenesis, as it is indispensably induced by growth factors specifically produced by the surrounding tumor cells. These growth factors, angiogenic proteins, include VEGF [11,29], basic fibroblast growth factor, and others [11]. VEGFR2 (in mouse=Flk1), the cell receptor of VEGF, is suggested to play an important role in tumor neoangiogenesis in humans [30]. Similarly, in the mouse model [29], it was shown that endothelial precursor cells from the bone marrow express VEGFR2 that is necessary for their role in tumor angiogenesis. These endothelial precursor cells were recruited for tumor angiogenesis from the bone marrow through VEGF signaling, eventually maturing into endothelial cells, positively stained for VE-cadherin. VEGFR2 [24,26] and VE-cadherin [26] are equally expressed by hemogenic endothelial cells, by which they may resemble those tumor endothelial precursor cells. Nevertheless, to the best of our knowledge, in that work like in all other works that deal with endothelial precursor cells in tumor angiogenesis, no reference was made to erythropoiesis that simultaneously takes place. Our question, therefore, is whether the VEGFR2+, endothelial precursor cells taking part in tumor angiogenesis, can be analogically compared with the hemogenic endothelial cells of prenatal development in their potential for embryonic or fetal erythropoiesis.
Concluding Remarks Regarding Therapeutic Implications
The advantage of HbF [31,32] and embryonic hemoglobin [33] stems from their efficient oxygen supply to the rapidly growing embryo and fetus, owing to their higher oxygen affinity, relative to HbA. Similarly, their production in situ in the tumors should benefit tumor oxygen supply over that of the normal tissues. Embryonic hemoglobin, although hardly evaluated in cancer, should be regarded equally to HbF, as potentially contributing to tumor growth by enhancing its oxygen supply; indeed, embryonic hemoglobin was recently found to be the predominant hemoglobin induced under hypoxic conditions in erythroid cells developing among cancer cells [34]. If we hypothesize that HbF cells are descendants of tumor endothelial precursor cells, as discussed previously, then it seems, quite probable, that antiangiogenic therapeutics in cancer, including anti-VEGF [12] and its receptor VEGFR2 [29], would be effective against HbF cell proliferation. However, there might be other routes of HbF cell regeneration in tumors, including the development of “primitive” HbF erythrocytes, such as in the yolk sac at the onset of embryonic development that might not share a common precursor with endothelial cells [14,19]. We should therefore consider anti-erythropoietic strategies, with an emphasis on HbF cells as contributing to the inhibition of tumor blood forming. In that regard, the idea of targeting erythropoiesis as a therapeutic strategy was very recently raised [34] in view of the failure of anti-angiogenic therapy, per se, to completely normalize tumor blood forming [12].
The involvement of HbF cell regeneration in tumor angiogenesis, although very common, was not, however, observed in all the cases with the same kind of tumor [1 –3]. It was predominantly observed in the early stages of tumor development [3], while in the later stages we found mainly normal HbA in the proliferating blood vessels. It seems, therefore, that there are at least two different kinds of hematoangiogenesis in tumors as related to the hemoglobin species. Each of these processes should be independently investigated for the development of the proper therapeutics.
Further experimental evidence is still needed in order to establish our hypothesis about in situ proliferation of HbF cells in tumors and its therapeutical implications. This investigation shall include more immunohistochemical examinations in various solid tumors regarding their stage, as well as using other methods at the cellular, biochemical, and molecular levels, for which we foresee the beginning [34]. In addition, the involvement of embryonic hemoglobin in tumor should also be examined in the future.
Footnotes
Acknowledgment
The author greatly appreciates the contribution to this work by the team of core pathology laboratory, headed by Prof. J.E. Martin, at the Blizard Institute, Queen Mary University of London.
Author Disclosure Statement
No competing financial interests exist.