
Abstract
A hyaluronic-acid-rich node and duct system (HAR-NDS) was found on the surface of internal organs of mice, and inside their blood and lymph vessels. The nodes (HAR-Ns) were filled with immune cells of the innate system and were especially enriched with mast cells and histiocytes. They also contained hematopoietic progenitor cells (HPCs), such as granulocyte-macrophage, erythroid, multipotential progenitors, and mast cell progenitors (MCPs). MCPs were the most abundant among the HPCs in HAR-Ns. Their frequency was fivefold higher than that of the MCPs in bone marrow. In addition, the system contained pluripotent stem cells (PSCs) capable of producing CD45−Flk1+ hemangioblast-like cells, which subsequently generated various types of HPCs and differentiated blood cells. Although HAR-Ns did not appear to harbor enough number of cells capable of long-term reconstitution or short-term radioprotection of lethally irradiated recipients, bone marrow cells were able to engraft in the HAR-NDS and reconstitute hematopoietic potentials of the system. PSCs and HPCs were consistently found in intravenous, intralymphatic, and intestinal HAR-ND. We infer that PSCs and HPCs reside in the HAR-ND and that this novel system may serve as an alternative means to traffic immature and mature blood cells throughout the body.
Introduction
T
We [6] have described a microscopic node and duct system, which appeared to be the Bonghan or primo vascular system, that was found on the surface of internal organs, and inside blood and lymphatic vessels in rats. The nodes were filled with innate immune cells, and were especially enriched with mast cells, eosinophils, basophils, neutrophils, and histiocytes. Curiously, they also contained chromaffin cells that produced epinephrine and norepinephrine. Secretory granules from mast cells moved through the ducts, and the nodes and ducts could be stained with alcian blue, which indicated that the system was rich in hyaluronic acid [6]. Hence we named it the hyaluronic-acid-rich node and duct system (HAR-NDS), and referred to the nodes and ducts as HAR-Ns and HAR-Ds, respectively, and to the two together as HAR-NDs.
The HAR-NDs appeared to form a network throughout the body, on the surface of organs, inside lymphatics, inside blood vessels, and along the nervous system [7]. We observed that ∼2% of the cells in the nodes were immature cells [6], and hypothesized that the system might contain pluripotent and committed stem cells. In this study, we examined whether hematopoietic stem and progenitor cells (HSPCs) reside in the HAR-NDS of mice.
Materials and Methods
Collection of HAR-NDs
The mice were purchased from a local vendor (Orient) and housed in the SPF facility in the National Cancer Center (NCC), Korea. Some mice were obtained from the animal care of the NIDDK NIH Center of Excellence in Molecular Hematology at the Indiana University School of Medicine (IUSM, Indianapolis, IN). The animal studies were approved by the Institutional Animal Care and Use Committee of the NCC, and the IUSM.
Wild-type, IFNγ−/− and IFNγ+/− mice on a C57Bl/6 background were anesthetized by i.m. injection of Zoletil (2.5 mg/kg) and Rompun (0.5 mg/kg). To collect HAR-NDs on organ surfaces, an incision was made along the abdominal linea alba and HAR-NDs were collected between the anterior wall and the intestine or liver while the abdominal wall was carefully lifted away.
To collect venous components of the HAR-NDS, ∼0.5 mL of 1% alcian blue was injected into one of the common iliac veins, and, with the top and bottom of the lumbar vein clamped by forceps, blood was drained by making an incision along the blood vessel. HAR-NDs were detected because they formed a blue line inside the vein. To collect intralymphatic HAR-NDs, 0.5 mL of 1% alcian blue was injected into the lateral tail base s.c. 1 cm caudal to the rectum, and medial to the tail vein [8,9].
HAR-NDs were observed and collected under a stereomicroscope (Zeiss Stereo Discovery.V20) with a camera (Zeiss AxioCamHRc camera).
Hyaluronic acid assays
HAR-NDs on organ surfaces were collected, weighed, and rapidly frozen with phosphate-buffered saline (PBS) in liquid nitrogen. The frozen HAR-NDs were homogenized with grinders and the supernatants were collected after centrifugation (20 min, 4°C, 2,000 g). The amount of hyaluronic acid in the supernatants was measured with a mouse hyaluronic acid ELISA kit (SunRed, Shanghai Sunred Biological Technology) according to the manufacturer's instructions. Serum, urine, peritoneal fluid, and lymph vessels were also assessed for hyaluronic acid.
Clonogenic assays
Bone marrow mononuclear cells (BM-MNCs) were collected by flushing the medullary cavities of tibias and femurs with PBS (pH 7.4) using a 25G needle. A single-cell suspension of HAR-NDs was obtained by dissociating the collected HAR-NDs with a cell strainer tube. The HAR-ND cells were plated at 1×105 cells/mL, and BM-MNCs were plated at 0.25×105 cells/mL in 1% methylcellulose culture medium in the presence of 0.1 mM hemin and 30% FBS (Hyclone). Growth factors used were as follows: 1 U/mL of recombinant human erythropoietin (STEMCELL Technologies), 50 ng/mL murine recombinant stem cell factor (rSCF; R&D Systems), 10 ng/mL murine rGM-CSF (STEMCELL Technologies), 10 ng/mL murine rIL-3 (STEMCELL Technologies), or, in place of GM-CSF and IL-3, 5% vol/vol pokeweed mitogen-stimulated mouse spleen-cell-conditioned medium [10]. We also used MethoCult GFM 3534 and GFM 3434 from STEMCELL Technologies to grow colony-forming unit–granulocyte macrophage (CFU-GM) and colony-forming unit–granulocyte erythroid macrophage megakaryocyte (CFU-GEMM) colonies, respectively. Colonies formed from HAR-NDs and BM were scored between days 7 and 14 under an inverted microscope (Olympus CKX31). Colony assays in methylcellulose were used to detect hematopoietic progenitors isolated from the HAR-NDS and BM. To examine their morphology, single colonies were resuspended in PBS and fixed in 10% neutral-buffered formalin (NBF, pH 7.4). The component cells were then harvested by centrifugation, resuspended in PBS, and mounted on slides. They were then stained with Wright–Giemsa or toluidine blue, and observed under a light microscope (Leica DMD108).
Mast cell progenitor (MCP) colonies derived from HAR-NDs were harvested and cultured in RPMI1640 medium supplemented with 10% FBS, 2 mM L-glutamine (Gibco), 0.1 mM NEAA (Gibco), 50 μM 2-ME, and 100 U/mL penicillin/streptomycin containing 10 ng/mL murine rIL-3 (STEMCELL Technologies) and 10 ng/mL murine rSCF (STEMCELL Technologies) [11,12]. The cells were cultured for 2–4 weeks at 37°C in a humidified chamber under 5% CO2 and harvested for flow cytometric analysis. Expression of FcɛRI was determined after preincubating the cells with 1 μg/mL α-DNP IgE (Sigma) for 24 h at 37°C to enhance FcɛRI expression [13]. Colony assays were performed in Korea and the United States.
Antibodies and flow cytometric analysis
All antibodies were purchased from BD Pharmingen, and the following antibody markers were used: Sca-1 (E13-161.7) as a phycoerythrin (PE) conjugate; c-kit (2B8), IgE (R35-72), Gr-1 (RB3-8C5), CD11b (M1/70), CD8 (53–6.7), and Flk-1 (AVAS12) as fluorescein isothiocyanate (FITC) conjugates; CD45 (30-F11), CD4 (H129.19), B220 (RA3-6B2), and CD135 (Flt3; A2F10) as phycoerythrin-Cy5 conjugates; Alexa Fluor 647-conjugated CD34 (RAM34) and CD150 (TC15-12F 12.2); and biotinylated lineage cocktail. Biotinylated primary antibodies were developed with either streptavidin-FITC or streptavidin-PE. PE-Cy5-conjugated CD135 (Flt3; A2F10) was purchased from eBioscience. Flow cytometry was performed on an FACSCalibur or LSR II flow cytometer (Becton Dickinson).
Differentiation of potential pluripotent stem cells into hemangioblast-like cells and blood cells
HAR-NDS cells collected from organ surfaces were plated at 1×105 cells over OP9 cells and cultured for 6 days in α-MEM containing 20% FBS, and antibiotics at 37°C in a humidified chamber under 5% CO2. OP9 cells were used to generate B-lineage and myeloid cells, and OP9-DL1 cells were used to support T-lineage cells. The following cytokines were added to the culture medium: recombinant mouse SCF (50 ng/mL; PeproTech), recombinant mouse Flt3L (5 ng/mL; ProSpec), and recombinant mouse IL-7 (5 ng/mL; ProSpec). Cobblestone-area-forming cells (CAFCs) were produced on OP9 monolayers with or without mSCF, and the resulting hemangioblast-like cells were analyzed by FACS after staining with CD45 (PE-Cy5) and Flk-1 (FITC) antibodies. Cytokine-induced myeloid and B-lineage cells were detected with Gr-1/CD11b (FITC) in combination with CD45 (PE-Cy5) and B220 (PE-Cy5) antibodies, respectively, while T-lineage cells were identified with CD4 (PECy5) and CD8 (FITC) antibodies.
CAFCs were plated in 1% methylcellulose-based GEMM medium to induce hematopoietic progenitor cells (HPCs). Colonies were harvested, pooled, and replated at 1×105 cells per mL into fresh GEMM medium every 10 days. This replating strategy was repeated several times to confirm the differentiation potential of these CAFCs. Further, differentiation of the HSPCs into various blood cells was detected under a light microscope after Wright–Giemsa or toluidine blue staining.
Hematopoietic engraftment assay
For the competitive repopulating HSC assay, lethally irradiated (1100 cGy) 8-week-old C57Bl/6 F1 mice (CD45.1+/CD45.2+) were reconstituted with a mixture of 5×105 HAR-NDS cells from B6 (CD45.2+) mice and an equal number of B6.BoyJ (CD45.1+) bone marrow cells (BMCs). Engraftment of donor cells was assayed at 1 and 3 months post-transplantation. For the noncompetitive engraftment assay, 5×105 HAR-NDS cells (Thy1.2+) were injected i.v. into lethally irradiated (1100 cGy) congenic mice (Thy1.1+). Survival of the recipients, and engraftment, was examined. To determine engraftment potential of HAR-ND-derived CAFCs, EGFP+CAFCs were prepared by culturing HAR-N cells from the intestinal surface HAR-ND of EGFP mice (C57BL/6-Tg CAG-EGFP 10sb/J; Jackson Laboratory) on OP9 cells in the presence of 50 ng/mL mrSCF (Prospec) for 7 days. EGFP+CAFCs (1×105) were mixed with EGFP− BMCs (4×105) prepared from age-matched C57BL/6 mice. The mixture was injected into the tail vein of lethally irradiated (1100 cGy) normal (EGFP−) C57BL/6 mice. Two months after the injection, intestinal surface HAR-NDs were collected and searched for EGFP+ cells under fluorescence inverted microscope (Observer Z1; Zeiss). Engrafting studies were done in Korea and the United States.
Trafficking of BM cells to the HAR-NDS
Lethally irradiated (1100 cGy) B6 mice were injected i.v. with EGFP+ syngeneic BM-MNCs at 2×106 cells per mouse. Engraftment of EGFP+ BM-MNCs in the HAR-NDS was examined under a fluorescence inverted microscope (Observer Z1; Zeiss).
Electron microscopy of HAR-NDs
Intestinal HAR-NDs were collected and fixed for 2 h at 4°C in Karnovsky's fixative (2% paraformaldehyde and 2% glutaraldehyde in 0.05 M sodium cacodylate buffer [pH 7.2], at 4°C). For transmission electron microscopy, HAR-NDs were postfixed for 2 h at 4°C in 1% osmium tetroxide (EMS), dehydrated in an ethanol series, embedded in SURR resin (ERL, DER, NSA, and DMAE mixture EMS), and polymerized at 70°C overnight. Ultrathin (0.5–1.0 μm) sections were cut with an ultramicrotome (RMC MTX) employing a diamond knife (Diatome), and stained with uranyl acetate (EMS) for 20 min followed by lead citrate for 10 min. The sections were examined with a transmission electron microscope (JEOL JEM1010) operated at an accelerating voltage of 80 kV. For scanning electron microscopy, HAR-NDs were washed three times for 10 min each with 0.05 M sodium cacodylate buffer (pH 7.2, 4°C) after fixation in Karnovsky's fixative. Next, they were postfixed in 1% osmium tetroxide in 0.05 M sodium cacodylate buffer (pH 7.2) and briefly washed twice with distilled water at room temperature. After dehydration with an ethanol series at room temperature, each for 10 min, the HAR-NDs were stiffened by two incubations for 10 min each in 100% isoamyl acetate at room temperature and critical point dried with liquid carbon dioxide. A field-emission scanning electron microscope (Carl Zeiss SUPRA 55VP) was used to examine the HAR-NDs mounted on metal stubs and coated with gold using a sputter coater (BAL-TEC/SCD 005).
Results
Isolation of components of the HAR-NDS
The HAR-NDS can be demonstrated without staining in the form of nodes and interconnecting thread-like ducts that generate a spider's web on the surface of organs (Fig. 1A-a). HAR-NDs can also be seen inside veins (Fig. 1A-b) and lymphatics (Fig. 1A-c, d) by alcian blue staining. We often found that the HAR-Ds branched off from a nearby node (asterisks in Fig. 1A-a, b). Scanning electron microscopy (SEM) revealed that the intestinal HAR-Ns appeared as oval-shaped sacs with ducts at both prolate ends (Fig. 1B-a). The insides of the HAR-Ns were filled with resident cells (Fig. 1B-b). SEM of the HAR-Ds showed that they were composed of three subducts (ductules) in this example (Fig. 1B-c, d). Transmission electron microscopy (TEM) revealed that the HAR-N had three sinuses (arrows), which served as paths for the three ductules, and was filled with cells (Fig. 1C-a). Some of them were identified morphologically as mast cells (Fig. 1C-b), polymorphonuclear leukocytes (Fig. 1C-c), monocytes (Fig. 1C-d), eosinophils (Fig. 1C-e), and small cells with very large nuclei and little cytoplasm (Fig. 1C-f, g).
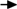
Hyaluronic-acid-rich node and duct (HAR-ND) on the surface of the intestine and inside veins and lymphatics of the mouse. ) stained with alcian blue can be seen inside a lumbar vein ( )
)  , magnification,×50
, magnification,×50
We measured the concentrations (ng/L) of hyaluronic acid in HAR-NDs, lymphatics (lymph vessel), serum, urine, and peritoneal fluid (PF). HAR-NDs had the highest concentration of hyaluronic acid of the samples tested (Fig. 1D), consistent with their preferential staining with alcian blue.
HAR-NDS contains HPCs
We collected the HAR-NDs from organ surfaces, and from the interiors of veins and lymphatics. Next, we isolated cells from these three different sites, and used the cells in hematopoietic progenitor assays. CFU-GM, burst-forming unit–erythroid (BFU-E), and CFU-GEMM colonies were obtained (Fig. 2A-a–c). We identified all three progenitor types formed from HAR-NDs on intestinal surfaces and inside vein but only CFU-GEMM and CFU-GM from HAR-NDs inside lymphatics (Supplementary Table 1; Supplementary Data are available online at
Hematopoietic progenitors are found in HAR-NDs.
Interestingly, the colonies grown under CFU-GM conditions were mainly MCP colonies (Fig. 2A-d), identified by staining with toluidine blue and Wright–Giemsa (data not shown). Wright–Giemsa staining revealed precursors to basophils/megakaryocytes/eosinophils/erythroblasts (Fig. 2B) in CFU-GEMM colonies (Fig. 2A-a); neutrophils/macrophages (Fig. 2B) in CFU-GM colonies (Fig. 2A-b); erythroblasts/erythrocytes (Fig. 2B) in BFU-E colonies (Fig. 2A-c); and mast cells (Fig. 2B) in MCP colonies (Fig. 2A-d).
There were far fewer CFU-GEMM, CFU-GM, and BFU-E colonies from HAR-NDs than from BM (Table 1). Although the frequencies of CFU-GM and BFU-E colonies in spleens were higher than those in the HAR-NDS, the frequencies of CFU-GEMM colonies in spleen and HAR-NDS were similar (Table 1). On a per-cell basis, however, the number of MCP from HAR-NDS was approximately fivefold higher than from BM and 100-fold higher than from spleen (Table 1 and Fig. 2C).
HAR-ND was obtained from the surface of intestine.
BM (2.5×104) and spleen cells (1×105), and HAR-ND cells from the surface of the intestine (1×105) were cultured for 7–14 days in 1% methylcellulose-based medium in the presence of 0.1 mM hemin, 30% FBS (Hyclone), and other growth factors. Numbers of CFU-GEMM, CFU-GM, and BFU-E colonies were scored on day 7, and MCPs on day 14.
Numbers are mean±SD of three independent experiments.
CFU-GEMM, colony-forming unit–granulocyte erythroid macrophage megakaryocyte; CFU-GM, colony-forming unit–granulocyte macrophage; BFU-E, burst-forming unit–erythroid colonies; HAR-ND, hyaluronic-acid-rich node and duct; MCPs, mast cell progenitors.
To confirm that cells from the MCP colonies developed into mast cells, we took the cells directly from MCP colonies. We then used a portion of colony-derived cells for phenotype analysis before culture and put the rest into culture in liquid medium supplemented with IL-3 and rSCF for 14 days. The cells from the MCP colonies and 14-day cultures displayed the lin−sca-1+c-kit+FcɛRI+ phenotype, confirming that they were mast cells (Fig. 2D; left and right panels, and Supplementary Fig. S1).
Hemangioblast-like cells can be induced from HAR-ND cells
We isolated HAR-NDs on the surface of the intestine (shown in Fig. 1A-a) for further analysis of the immature cells. Phenotypic analysis revealed that ∼2% of the HAR-ND cells were lineage− sca-1+, c-kit+, and CD34−, suggesting that the HAR-NDs contain a small number of hematopoietic stem cells (HSCs). We then hypothesized that HAR-NDS might contain adult pluripotent stem cells (PSCs) that constantly gave rise to progenitor cells. We examined whether hemangioblasts, precursors of immature blood cells, could be induced from HAR-NDs by culturing the entire cellular content of HAR-NDs with OP9 cells. Hemangioblasts, which are CD45− but express Flk-1, can differentiate into all types of blood cells [14 –16].
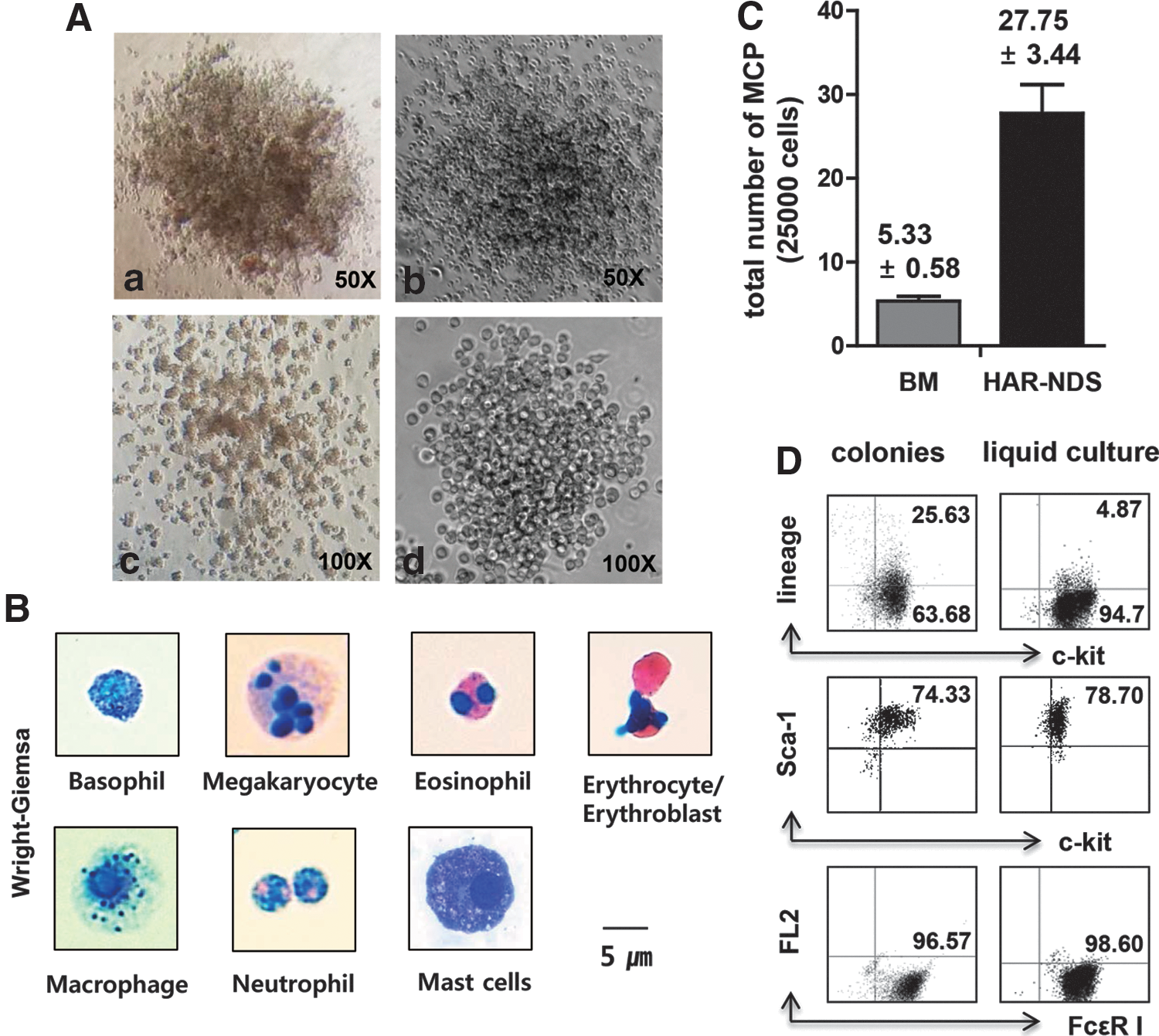
When we cocultured HAR-ND cells on OP9 cells for 6 days, CAFCs were produced (Fig. 3A). Four to five cobblestone areas were formed per 1×105 HAR-ND cells. When the entire CAFCs were harvested on day 6 and analyzed by flow cytometry, ∼2.3% of the cells were CD45−Flk-1+, and 12.4% were CD45+Flk-1− (Fig. 3B-a, b), suggesting that hemangioblast-like cells (CD45−Flk-1+ cells) had developed.
Identification of hematopoietic stem cells in HAR-NDs.
Wright–Giemsa staining of CAFCs showed small immature cells (∼5 μm in diameter) with high nucleus/cytoplasm ratio (Supplementary Fig. S2A). To examine further whether CAFCs would contain MCPs, the CAFCs were stimulated with α-DNP IgE and analyzed for their expression of FcɛRI. The CAFCs did not express FcɛRI, indicating that CAFCs were not mast cells or MCPs (Supplementary Fig. S2B).
We further analyzed the phenotype of the CAFCs formed in NDS/OP9 coculture. We found that the majority of the cells became CD45+Flk1− when the coculture was maintained for 10 days (Fig. 3C-a). Addition of rSCF greatly increased CD45+Flk− cell numbers. On day 10 more than 90% of the cells were lin−CD45+ (Fig. 3C-b). The majority of the cells (∼70%) were sca-1+c-kit+, and again CD34−CD135−. The major cell population of the CAFCs was lin−sca-1+c-kit+CD34−CD135− (Fig. 3C-c, d), thus displaying the phenotypic markers of primitive HSCs [17 –19]. Further analysis of these cells with SLAM markers [20 –22] (Fig. 3C-e) revealed that the major population (∼82.5%) was CD48+CD150− and thus lin−sca-1+c-kit+CD34−CD135−CD150−CD48+ (CD150−CD48+LSK), while a minority were CD150−CD48−LSK (14.8%) or CD150+CD48−LSK (0.5%) (Fig. 3D).
When cells from HAR-NDs (1×105 cells) were cocultured on OP9 with rSCF and IL-3 for 9 days, myeloid lineage cells appeared (Fig. 3E-a), and when they were cocultured on OP9 in the presence of rSCF, IL-7, and Flt3L for 15 days, B220+ B lymphocytes were detected (Fig. 3E-b). If instead they were cocultured on OP9-DL1 with rSCF, IL-7, and Flt3L for 9 days, then CD4+, CD4+CD8+, and CD8+ T lymphocytes were produced (Fig. 3E-c) [23,24]. These results indicate that HAR-ND cells have the potential to produce mature blood cell types under the right culture conditions.
When the CAFCs were cultured in GEMM methylcellulose medium for another 10 days, colonies consisting of uniformly small cells (colony-forming cells, CFCs) (∼5 μm in diameter) were formed (Fig. 4A), and the cells were lin−sca-1+c-kit+CD34−CD135−, suggesting that they still maintained the differentiation status of HSPCs [19,22] (Fig. 4B). These cells were replated every 10 days and their numbers increased over a 50-day culture period (Fig. 4C). Wright–Giemsa and toluidine blue staining after the fifth passage identified RBCs, immature megakaryocytes, MCPs, and monocytes (Fig. 4D). These results show that HAR-NDs contain immature pluripotent cells that can be differentiated into hemangioblast-like cells capable of generating phenotypically HSCs and functional HPCs, and mature blood cells.

Properties of colony-forming cells.
Differential regulation of MCPs
Because MCPs were more frequent in HAR-NDs than in bone marrow or spleen, we examined whether the production of MCPs was regulated differently in HAR-NDs than in bone marrow or spleen. We employed various gene-deletion mice that might reveal differences in MCP production between bone marrow and spleen on the one hand and HAR-NDs on the other hand.
IFN-γ has been shown to regulate mast cell development and function in vitro and in vivo [25 –28], and there was a drastic reduction of MCP production in the HAR-NDs of IFNγ−/− mice, suggesting that IFN-γ is closely associated with the production of HAR-ND MCPs (Fig. 5A). In contrast, the production of MCPs in the spleen was not dependent on IFN-γ (Fig. 5B-a, b). MCP production in HAR-NDs was also greatly reduced in c-kitW-sh/W-sh, compared with W-sash heterozygotes (Fig. 5C), indicating that the c-kit locus is important for HAR-ND mast cell development, as it is for the development of bone marrow and spleen mast cells.

Differential regulation of MCPs by IFNγ.
The hematopoietic engraftment potential of HAR-NDS cells
Competitive repopulation HSC assays using HAR-NDS (CD45.2+) donor cells and CD45.1+ competitor cells at a 1:1 ratio showed that HAR-NDS cells (CD45.2+) did not engraft in lethally irradiated F1 (CD45.1+/CD45.2+) mice whereas the competitor BM-MNCs (CD45.1+) did at 1, 2, and 4 months post-transplant. In contrast, at a 1:1 ratio of donor BM-MNC CD45.2+ cells and CD45.1+ competitor BM-MNCs, the percent chimerism of the donor BM-MNCs ranged from 40% to 55% over months 1, 2, and 4 (data not shown). This range of donor BM-MNCs in such a competitive transplant assay has been seen in numerous other studies done in our laboratory. Hence, the fact that HAR-NDS cells did not engraft in this competitive repopulating HSC assay suggests non- or low-engrafting capability of HAR-NDS donor cells. Moreover, when we transplanted EGFP+ HAR-NDS cells into lethally irradiated syngeneic mice at various doses up to 5×105 cells/mouse in a noncompetitive assay, all the mice died within 10–14 days of transplantation. These results indicate that HAR-NDS cells either lack radioprotective and HSC engrafting potential or the numbers of engrafting HAR-NDS cells are very low.
Then we determined whether or not HAR-NDS-derived cells could have engrafting potential. We transferred HAR-NDS-derived CAFCs with BMCs according to the scheme shown in Fig. 6A. Survival rate was a little higher in the HAR-NDS plus BM group compared with BM alone, although statistical significance could not be determined because of the small sample sizes (Fig. 6B). These mice survived more than 2 months. At 2 months, we were able to detect GFP+ cells in the intestinal HAR-ND group as shown in Fig. 6C. The data may suggest that HAR-NDS-derived CAFCs have potential to engraft and perhaps the number of cells that have engrafting potential is small.

In vivo transplantation of HAR-ND CAFCs.
BMCs engraft in the HAR-NDS
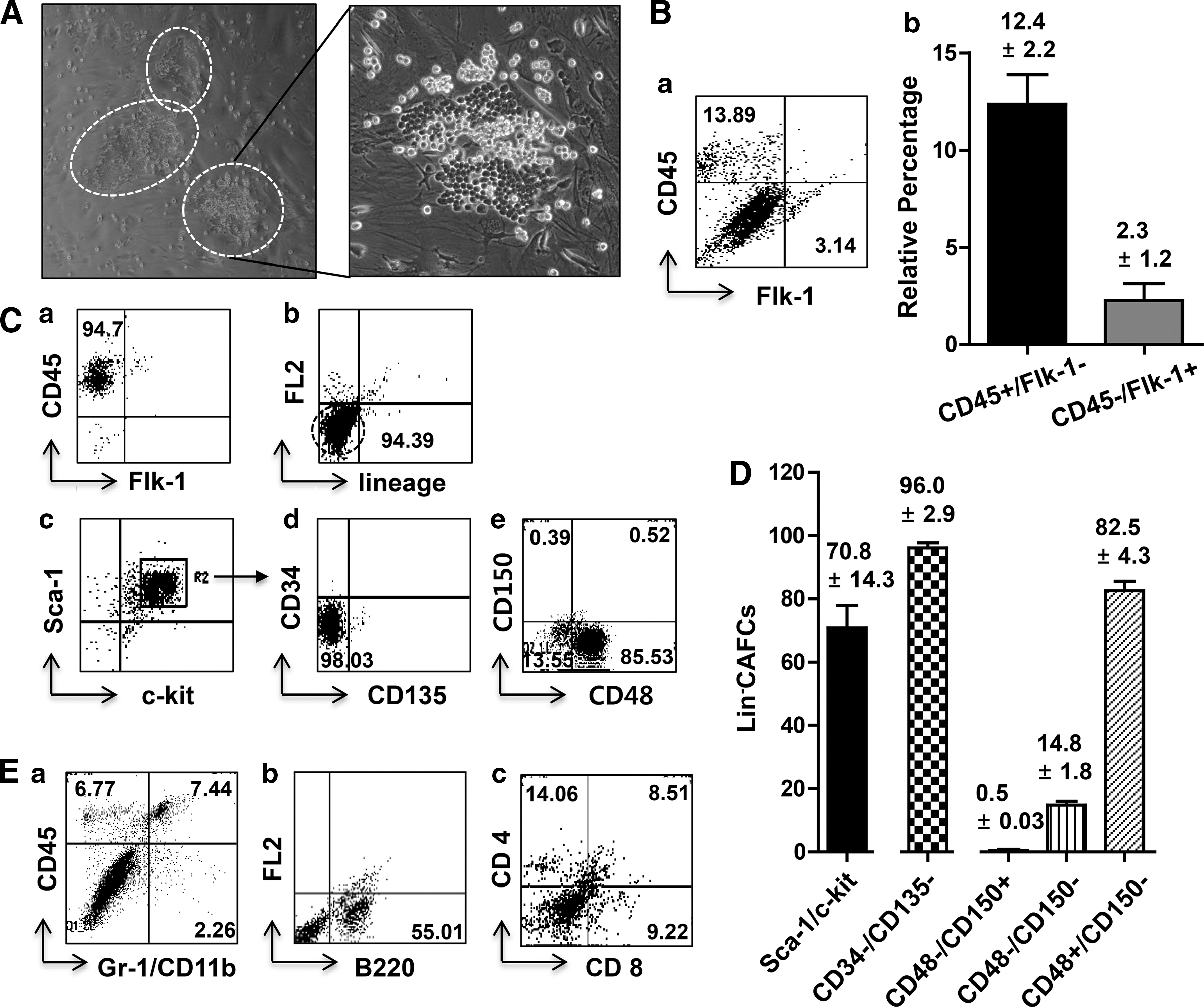
We examined whether BM-MNCs trafficked to and engrafted in the HAR-NDS. When we injected 2×106 EGFP+ BM-MNCs into syngeneic lethally irradiated B6 mice, EGFP+ cells were detected from day 10 after transplantation (Fig. 7A), and the HAR-NDS was fully reconstituted by day 21 (Fig. 7B). When cells from the reconstituted HAR-NDS were cultured over OP9, CAFCs were produced, indicating that the hemangioblast-like cell cycled between the bone marrow and the HAR-NDS (Fig. 7C).
Trafficking of bone marrow cells to the HAR-NDS. Detection in the HAR-NDS of BM-derived GFP+ cells on days 10
Discussion
We have described a new anatomical location, the HAR-NDS, occupied by cells with many of the known properties of hematopoietic progenitors. An anatomical system similar to the HAR-NDS was described, mainly in rabbits and humans, by Kim who named it the Bonghan vascular system [1 –3], and Soh et al., mainly studying rabbits and rats, renamed it the primo vascular system [4,5]. We have previously described an HAR-NDS in rats whose nodes were enriched with mast cells, eosinophils, basophils, neutrophils, and histiocytes, and through whose ducts secretory granules from mast cells appeared to circulate [6].
In the present work we wished to determine whether the HAR-NDS serves as a transient reservoir of immune cells, and whether this system is capable of producing blood cells independently of the BM and blood system. We therefore investigated whether HAR-ND contained hematopoietic progenitors and pluripotent cells that could generate HSCs. We have presented evidence that the HAR-NDS contains PSCs capable of generating hemangioblast-like cells that are able to direct nonmarrow hematopoiesis in the system (Fig. 3B).
HAR-ND on the intestine was enriched with MCPs. We identified mast cell precursors by metachromatic staining with toluidine blue, transmission electron microscopy, and by the expression of the membrane receptors for IgE. MCP colonies were first detected at 7 days of culture, and the maximum number of colonies was found on day 14, while GEMM or GM colonies were first detected on day 3 and the colony numbers reached their peak by day 7. This is in agreement with the previous report [11]. We stained all colonies on a plate on day 14 and found that they all displayed mast cell morphology. The c-kit gene was expressed in HSPCs but it was downregulated during further differentiation into various lineages of cells. Mast cells, however, expressed high c-kit levels and maintained during the development into mature mast cells [29]. When we stained the cells from MCP colonies with c-kit antibody, all of the cells were positive for c-kit (Supplementary Fig. S1A). Although MCPs were differentiated into mast cells, c-kit expression was maintained during the long period of in vitro culture (Supplementary Fig. S1B).
Bone-marrow-derived CAFCs formed colonies during the first 7 days of culture and started to die, and no cells survived in the second passage. In contrast, CAFCs from HAR-NDS stayed undifferentiated until the fourth passage and differentiated cells began to appear on the fifth passage. These cells started to die after the fifth passage (Fig. 4D). We, therefore, believe that CAFCs from HAR-NDS are different from the hematopoietic cells from bone marrow.
As shown in Fig. 6, HAR-ND-derived CAFCs contained immature cells of engrafting potential. HAR-ND cells alone, however, failed to produce long-term reconstituting or short-term radioprotective ability. Interestingly, BM cells did engraft in the HAR-NDS and appeared to reconstitute a certain hematopoietic potential on the part of HAR-ND cells (Fig. 7C). These results suggest that we may not have transplanted enough HAR-NDS-derived stem-like cells to reconstitute the lethally irradiated bone marrow.
The W-sash mutation (c-kitW-sh/W-sh) is known to disrupt normal mast cell development and survival [30]. c-kit signaling is required for the normal production of HAR-NDS-derived MCPs like that of bone-marrow- or spleen-derived MCPs. IFN-γ signaling, however, was critical only for the production of HAR-NDS-derived MCPs, not for that of spleen-derived MCPs. The reason for this differential effect requires investigation.
We believe that the HAR-NDS constitutes an important biological system because it appears to be densely distributed on the surface of all internal organs, inside the organs, under the skin, inside all vessels, and along the nervous system [7]. This system could be a route for delivering pluripotent cells, progenitor cells, and immune cells in a precise and controlled fashion to target areas.
Footnotes
Acknowledgments
This work was supported by grants from the Korean Research Foundation (KRF-2005-084-E00001), the National Cancer Center, Korea (NCC-1310430-2), and the Advanced Institute of Convergence Technology (AICT-1341540-1).
The authors wish to thank Drs. Nadia Carlesso, Sonia Rodriguez-Rodriguez, and Man Lee at the Indiana University School of Medicine for providing the OP9-DL1 cell line.
Author Disclosure Statement
The authors have no conflicts of interest to disclose.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.