
Abstract
MicroRNAs (miRNAs) are potential therapeutic targets in a variety of pathological conditions in the brain; however, their clinical application is hampered by lack of efficient delivery modes. Mesenchymal stromal stem cells (MSCs) migrate to sites of injury and inflammation and exert therapeutic effects in various neurological disorders. Here, we examined the ability of MSCs to deliver exogenous miRNA mimics and pre-miRNAs to human neural progenitor cells (NPCs) and astrocytes and characterized the functional impact of this delivery. We found that MSCs efficiently delivered fluorescent-labeled miR-124 and miR-145 mimics to cocultured NPCs and astrocytes. We further demonstrated the delivery of the miRNAs using novel reporter plasmids that contain a sequence complementary to miR-124 or miR-145 downstream of luciferase or mCherry. Binding of the specific miRNAs to these sequences results in decreased luciferase activity or mCherry fluorescence and therefore enable analysis of miRNA delivery in living cells. The delivered exogenous miR-124 significantly decreased the expression of the target gene Sox9 by targeting its 3′-UTR, and increased the neuronal differentiation of the NPCs. In addition, the delivered miR-124 increased the expression of the glutamate transporters, EAAT1 in NPCs and EAAT2 in both NPCs and astrocytes. Similar results were obtained with MSCs transfected with pre-miR-124. The miRNA delivery was mediated by MSC-derived exosomes and was cell contact independent. These results suggest that MSCs can functionally deliver exogenous miRNAs to neural cells and provide an efficient route of therapeutic miRNA delivery to the brain in pathological conditions with clinical implications for regenerative medicine.
Introduction
M
Adult human mesenchymal stromal stem cells (MSCs) obtained from autologous bone marrow or adipose tissues [16,17], can be easily expanded in vitro and genetically modified for therapeutic strategies [18]. Recent studies reported that MSCs exhibit homing abilities, which enable them to migrate to sites of brain injury, inflammation, and tumors [19]. In addition, MSCs have been shown to improve clinical outcomes in a variety of preclinical animal models and in preliminary clinical trials of different pathological conditions in the CNS [20,21]. Their effects have been attributed to the secretion of trophic factors, suppression of inflammatory responses, and effect on endogenous neurogenesis and neural differentiation [19,22 –24]. In addition, MSCs have been shown to act as vehicles of various therapeutic agents such as cytokines [20], oncolytic viruses, and peptides [25] for the treatment of both cancer and neurological disorders. We recently demonstrated that MSCs derived from different sources can also deliver miRNA mimics to tumor cells in vitro and in vivo and that the delivered miRNAs downregulate specific target genes and impact cellular functions of the recipient cells [26].
In this study we examined the ability of MSCs derived from bone marrow (BM), to deliver exogenous miRNAs to human neural progenitor cells (NPCs) and astrocytes. We found that the BM-derived MSCs delivered exogenous miR-124 and miR-145 to NPCs and astrocytes via contact independent and exosome-dependent processes. Moreover, the delivered miRNAs altered gene expression in the recipient neural cells and impacted their function.
Materials and Methods
Materials
Anti β3-tubulin and anti Sox9 antibodies were obtained from Chemicon. Rabbit anti-EAAT1 and EAAT2 antibodies were obtained from Santa Cruz Biotechnology and goat anti-actin antibody was obtained from Sigma.
Human NPCs and astrocytes
Human NPCs were obtained from Lonza, Inc. and neurospheres were maintained in NP maintenance medium (Lonza, Inc.). For the differentiation studies, the NPCs were disaggregated into small neurospheres and were plated on laminin-coated plates in NP basal medium with specific supplements according to the manufacturer's recommendations.
Human astrocytes were obtained from ScienCell Research Laboratories and were maintained in astrocytic medium according to the manufacturer's recommendations.
Mesenchymal stem cell cultures
BM-derived MSCs (BM-MSCs) were obtained from ScienCell Research Laboratories and were characterized and maintained as previously described [26]. Briefly, the cells were propagated and maintained in Dulbecco's modified Eagle's medium (DMEM) supplemented with fetal bovine serum (10%), gentamicin (50 μg/mL), nonessential amino acid (5 mM), and glutamine (5 mM) at 37°C in a 5% CO2 humidified atmosphere. Using FACS analysis the cells were found to be positive for CD73, CD90, and CD105 but negative for the hematopoietic markers CD14, CD34, and CD45.
Transfection of miRNA mimics
RNA duplexes corresponding to hsa-miR-124a and hsa-miR-145 labeled with Cy3 were synthesized by Sigma. Transfection of the MSCs with the miRNA duplexes was carried out with siIMPORTER (Millipore) according to the manufacturer's instructions and as previously described [27]. Briefly, 100 nM of diluted miRNA per well was combined with siIMPORTER reagent in DMEM serum-free medium. The transfection complex was directly added to the cells and replaced with a fresh medium after 24 h. This method resulted in the transfection of more than 90% of the MSCs as assessed by analysis of the fluorescent miRNAs. In some of the experiments, MSCs were transfected with the miRNA mimics by electroporation using the Nucleofector device, protocol number U29 (Amaxa Biosystems) to eliminate the presence of residual transfection reagents. Transfection efficiency using nucleofection was about 70% to 90%.
Lentiviral transduction of MSCs
Lentivirus-based vectors expressing pre-miR-124-GFP and control pre-miR-GFP were purchased from System Biosciences. Lentivirus production and titration were carried out according to the manufacturer's protocols. Transduction of MSCs was performed at an MOI of 20.
Cocultures of MSCs and neural cells
The green fluorescence CMFDA CellTracker reagent (Molecular Probes, Invitrogen) was used to label the NPCs and the astrocytes according to the manufacturer's protocol and as previously described [26]. Briefly, cells were incubated with a solution of 5 μM dye for 30 min at 37°C and were then thoroughly washed. The MSCs were transfected with fluorescently labeled miR-124 or miR-145 mimics 24 h before the experiment and were thoroughly washed following the transfection to remove residual miRNA and transfection reagent solutions. For the pre-miRNA experiments, the MSCs were transduced with lentivirus vectors expressing control pre-miR or pre-miR-124. Green CellTracker-labeled NPCs or astrocytes were added at a ratio of 1:2 to miRNA-transfected MSCs plated on chamber eight-well slides (ibidi GmbH) and were then analyzed by fluorescence microscopy.
Preparation of exosomes
MSCs were transfected with control miR or miR-124 mimic and maintained in OPTIMEM supplemented with 10% exosomes-free FBS. Following 3 days of transfection, exosomes were isolated from the supernatants of the MSC cultures using the exosome precipitation solution, ExoQuick, (System Biosciences) according to the manufacturer's instructions and as previously described [26]. The protein content of the enriched exosomal fractions was determined using the Micro BCA assay kit. The average exosome concentration from 10 mL of culture supernatant obtained from 5×107 cells (n=6) was 37.8 μg. For the exosome experiment, 2 μg of exosomes was administered to cultured NPCs.
Fluorescence microscopy
Cells were analyzed by fluorescence microscopy (Olympus, Cellsens Dimension) or by a LSM510 Meta confocal microscope equipped with ultraviolet, argon, and helium/neon lasers (Nikon).
Real-time quantitative PCR analysis
Total RNA was isolated using QIAzol reagent (Qiagen) according to the manufacturer's protocol. About 0.5 μg of RNA was employed to synthesize cDNA by Thermoscript (Invitrogen) with oligo dT primers. Sox9 mRNA was measured by the SYBR green method using the following primers: Sox9–forward CCAGCAAGAACAAGCCGCACG; reverse CTCCGCCTCCTCCACGAAG, Nestin–forward GGCTGCGGGCTACTGAAAA; reverse AGGGACATCTTGAGGTGCG, β-III tubulin–forward CTTTGGTCAGAGTGGGGC; reverse CAGGCAGTCGCAGTTTTC, and S12–forward TGCTGGAGGTGTAATGGACG; reverse CAAGCACACAAAGATGGGCT. S12 mRNA was used as an internal control. Expression of miR-124 was determined by TaqMan stem-loop RT-PCR method. The primers and probes of miR-124 and the endogenous control RNU6B were purchased from System Biosciences. Quantitative miRNA or mRNA expression data were acquired and analyzed using the ABI Prism 7000 Sequence Detection System (Applied Biosystems). Data were further analyzed by Comparative CT (ΔΔCT) method, and results are expressed in arbitrary units.
Luciferase reporter assays
NPCs were co-transfected with the pEZX-MT01-Sox9-3′-UTR construct (GeneCopoeia) and the pRL-TK Renilla luciferase control vector (Promega). Transfection was performed by electroporation using the Nucleofector device program A027 and the mouse NPC Nucleofector kit (Amaxa Biosystems). The cells were also transfected with miR-124 or a control miRNA using siIMPORTER. After 48 h incubation, firefly and Renilla luciferase activities were measured using the dual-luciferase assay kit (Promega), according to the manufacturer's instructions.
miR-124 reporter system
For analyzing miR-124 delivery, we generated a miR-124 luciferase and mCherry reporter plasmids. A unique miR-124 binding site (ATCAAGGCCGCTGTGAACACG), which is a fully complementary sequence of mature miR-124, was cloned downstream of luciferase reporter gene of the pMiR-Luc reporter vector from Signosis, Inc. For the mCherry reporter, the luciferase gene of pMiR-Luc reporter vector was replaced with pmCherry-N1 obtained from Clonetech. Binding of miR-124 to the specific binding site decreases the luciferase activity or the mCherry fluorescence, which can be used for the monitoring of changes in miR-124 levels in living cells. The miR-145 luciferase reporter was purchased from Signosis.
Western blot analysis
Cell lysates (30 μg protein) were resolved by SDS-PAGE and transferred to nitrocellulose membranes as previously described [28]. Following incubation with the primary and secondary antibodies the immunoreactive bands were visualized by the ECL Western blotting detection kit (Amersham).
Statistical analysis
The results are presented as the mean value±SE. Data were analyzed using analysis of variance and a Student's t-test.
Results
MSCs deliver a miR-124 mimic to neural cells
MSCs have been reported to secrete multiple factors including cytokines and extracellular matrix molecules that promote neurogenesis [29,30]. In addition, MSCs can be also genetically modified to deliver specific growth factors and cytokines [18]. Here, we examined the ability of BM-MSCs to deliver exogenous miRNA mimics to human NPCs and astrocytes. We focused on the delivery of miR-124 since this miRNA has been reported to play a role in neurogenesis and BM-MSCs express very low levels of this miRNA. We first examined the expression of miR-124 in human NPCs and found that the NPCs expressed low levels of this miRNA, which increased following differentiation of the cells (Fig. 1A). Similarly, we found that transfection of the human NPCs with miR-124 increased their neuronal differentiation as indicated by their morphological appearance (Fig. 1B), the increased β3-tubulin and the decreased nestin expression (Fig. 1C).
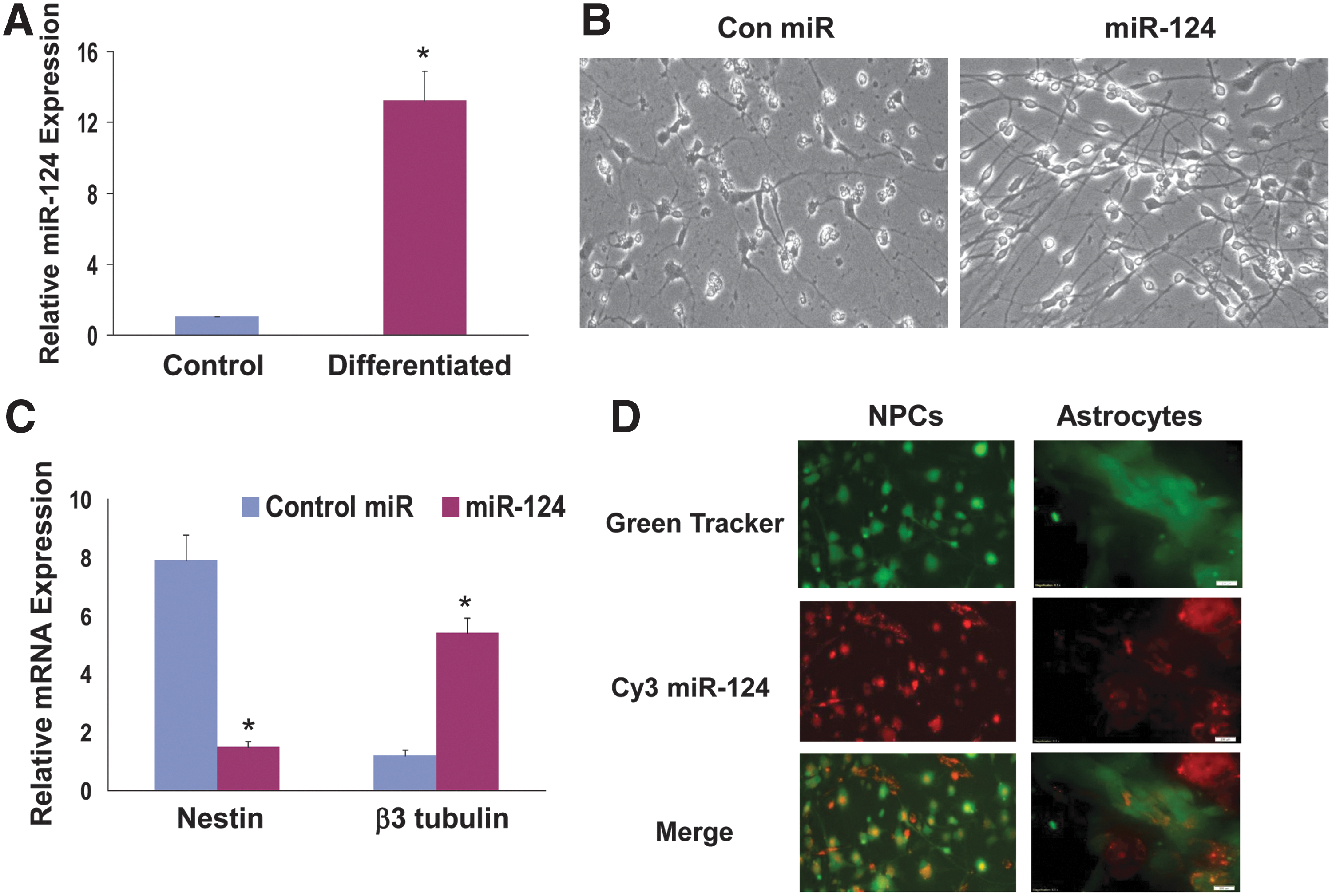
MSCs deliver exogenous miRNAs to cocultured neural cells. The expression of miR-124 was measured in control and differentiated NPCs using RT-PCR.
Transfection of the astrocytes with miR-124 did not alter their morphology or the expression of GFAP (data not shown).
To examine the ability of MSCs to deliver miR-124 mimic to NPCs and astrocytes we employed MSCs transfected with a miR-124 mimic labeled with Cy3 as previously described [26] and human NPCs or astrocytes labeled with Green Cell Tracker. Once in the cells, this cell tracker reagent is transformed into a nonpermeable fluorescent dye that is not transferred between cells. Following 2 days of coculture, the cells were viewed under a fluorescence microscope. As presented in Fig. 1D, Cy3-miR-124 was strongly expressed in the MSCs (red alone) and in most of the NPCs and astrocytes that were labeled with the Green Cell Tracker (Fig. 1D). Merged images demonstrated that the transfected MSCs efficiently delivered the miR-124 mimic into the adjacent cocultured cells (Fig. 1D). The miR-124 mimic delivery was observed up to 7 days post coculturing (data not shown).
Generation of reporter plasmids for the analysis of miR-124 delivery
To further monitor the delivery of the miR-124 mimic, we generated luciferase and mCherry-based miR-124 reporter plasmids by inserting a unique miR-124 target site downstream of either luciferase or mCherry. Thus, increases in the expression of miR-124 in the cells result in the downregulation of either luciferase activity or mCherry immunofluorescence. We first demonstrated the ability of the luciferase and mCherry-based miR-124 reporter plasmids to monitor miR-124 levels in the cells. NPCs were transfected with the miR-124 luciferase reporter and 24 h later the cells were transfected with a miR-124 mimic. Transfection of the NPCs with the miR-124 mimic increased the expression of miR-124 level as determined by real-time PCR (Fig. 2A). The miR-124-transfected cells exhibited a 72% decrease in the luciferase activity of the miR-124 reporter plasmid (Fig. 2B) and similarly a large decrease in the fluorescence of the mCherry reporter as compared to cells transfected with a control miRNA (Fig. 2C). We next examined the ability of these reporters to detect delivery of miR-124 mimic by MSCs. For these experiments, NPCs were transfected with either miR-124 reporter tagged to luciferase or to mCherry and after 24 h NPCs were cocultured with MSCs expressing Cy3-miR-124. After additional 24 h, the cocultures were examined for luciferase activity or for immunofluorescence. Figure 2D demonstrates that coculture of NPCs expressing the miR-124 luciferase reporter with MSCs transfected with Cy3-miR-124 exhibited a decrease of about 61% in luciferase activity compared with NPCs cocultured with MSCs transfected with a control miRNA. Similar results were obtained with NPCs transfected with miR-124 reporter tagged to mCherry (data not shown), although these data were more difficult to quantitate. Analysis of the delivery of miR-124 by MSCs to cocultured astrocytes gave similar results and luciferase activity of the miR-124 reporter decreased by 49.7%±5.9% (data not shown).
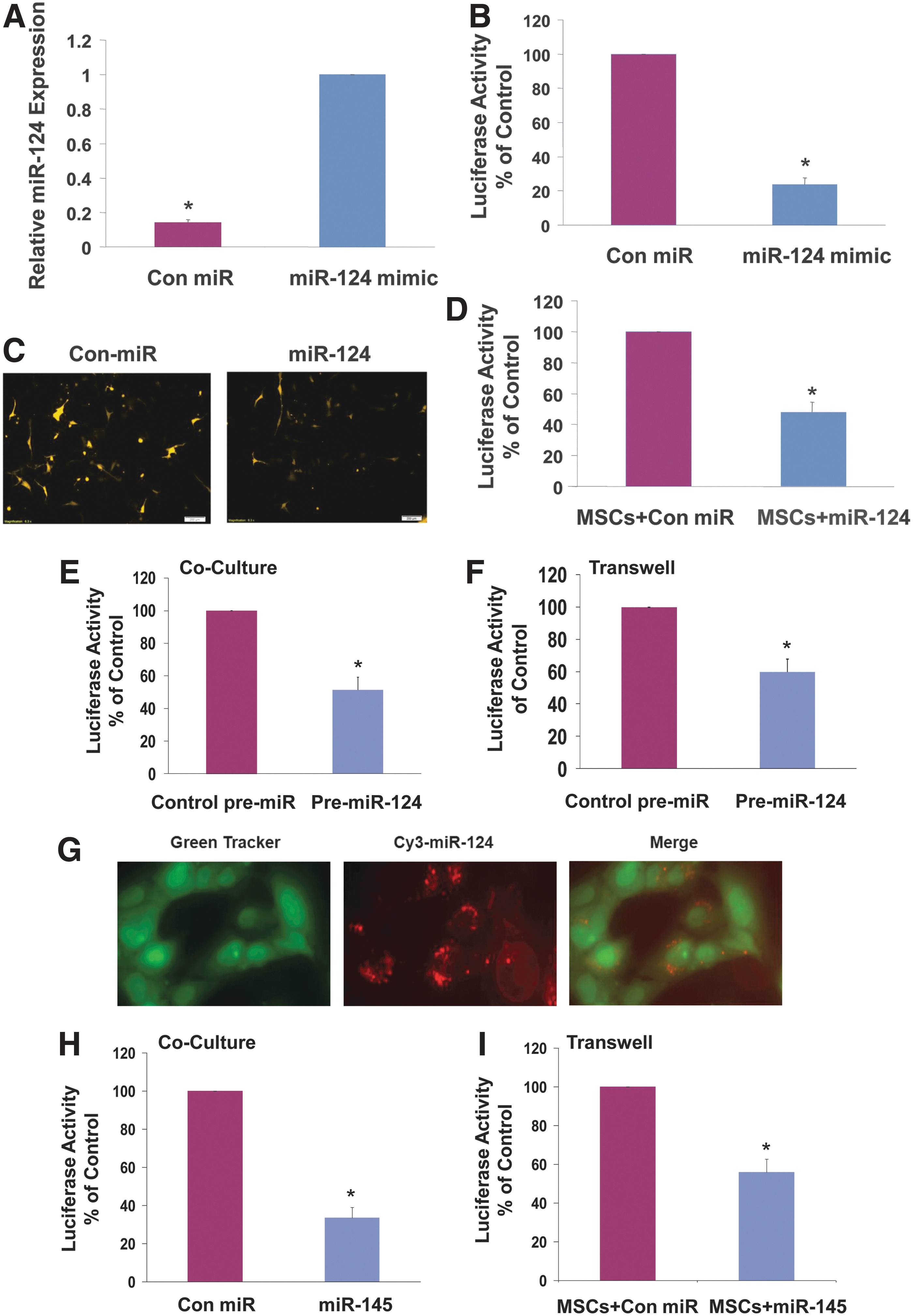
Generation of miRNA reporter systems for the analysis of miRNA delivery. NPCs were transfected with a miR-124 mimic and the expression of miR-124 was measured using RT-PCR
We further analyzed the delivery of mature miR-124 using MSCs transduced with a lentivirus vector expressing pre-miR-124. BM-MSCs were transduced with a lentivirus vector expressing pre-miR-124 for 3 days. Cocultures of the transduced MSCs and astrocytes transfected with the miR-124 luciferase reporter resulted in a 54.6%±6.8% decrease in luciferase activity, demonstrating the ability of the transduced MSCs to deliver mature miR-124 to astrocytes (Fig. 2E). Similar results were also obtained when cells were assayed in transwell chambers with 0.4 μm pore-size membranes that do not allow the infiltration of the cells (Fig. 2F). In summary, these results demonstrate the ability of the reporter systems to efficiently measure miR-124 delivery in living cells in vitro and probably in vivo as well.
To demonstrate that the ability of MSCs to deliver exogenous miRNAs to neural cells was not limited to miR-124, we employed MSCs transfected with Cy3-miR-145 and examined the ability of the MSCs to deliver the fluorescent miRNA mimic to cocultured astrocytes. As presented in Fig. 2G, the MSCs delivered the Cy3-miR-145 to the cocultured astrocytes, similar to the results obtained for Cy3-miR-124 (Fig. 1D). Using a luciferase miR-145 reporter, we demonstrated that transfection of human astrocytes with miR-145 significantly decreased the luciferase activity of the miR-145 reporter in these cells (Fig. 2H). Similarly, coculture of MSCs transfected with miR-145 and astrocytes transfected with the luciferase miR-145 reporter in a transwell with 0.4 μm filter also resulted in a decrease of the luciferase activity of the astrocytes (Fig. 2I), indicating delivery of miR-145 by the MSCs.
MSC-delivered miR-124 mimic downregulates Sox9 gene expression in NPCs
To examine whether the delivered miR-124 mimic was functional in the NPCs, we analyzed its ability to decrease the expression of one of miR-124 known target gene, Sox9. We first examined the effect of miR-124 mimic on the expression on Sox9 in the NPCs and found that transfection of these cells with the miR-124 mimic decreased Sox9 mRNA (Fig. 3A) and protein (Fig. 3B) levels. Using a luciferase reporter assay with a construct containing the 3′-UTR of Sox9 cloned downstream of firefly luciferase, we found that the miR-124 mimic significantly decreased the luciferase activity of this construct in the NPCs (Fig. 3C). We then expressed the Sox9 3′-UTR-luciferase plasmid in the NPCs cells and examined the luciferase activity in cells when cocultured with MSCs transfected with a control miR or with the miR-124 mimic. As presented in Fig. 3D, coculture of the Cy3-miR-124-transfected MSCs with the NPCs decreased the luciferase activity of the Sox9 reporter, whereas no significant decrease was observed in NPCs cultured with miR-control-transfected cells. We further employed this system to examine the ability of MSCs overexpressing pre-miR-124 to deliver miR-124 to NPCs. The expression of miR-124 in these cells was measured using real-time PCR (Fig. 3E) and the MSCs were then cocultured with NPCs transfected with the Sox9-luciferase reporter. Following additional 24 h, the luciferase activity of the cocultures was determined. As demonstrated in Fig. 3F, the luciferase activity of the NPCs cocultured with MSCs transduced with pre-miR-124 decreased to 48.7% of the luciferase activity of NPCs cocultured with MSCs transduced with a control pre-miR.
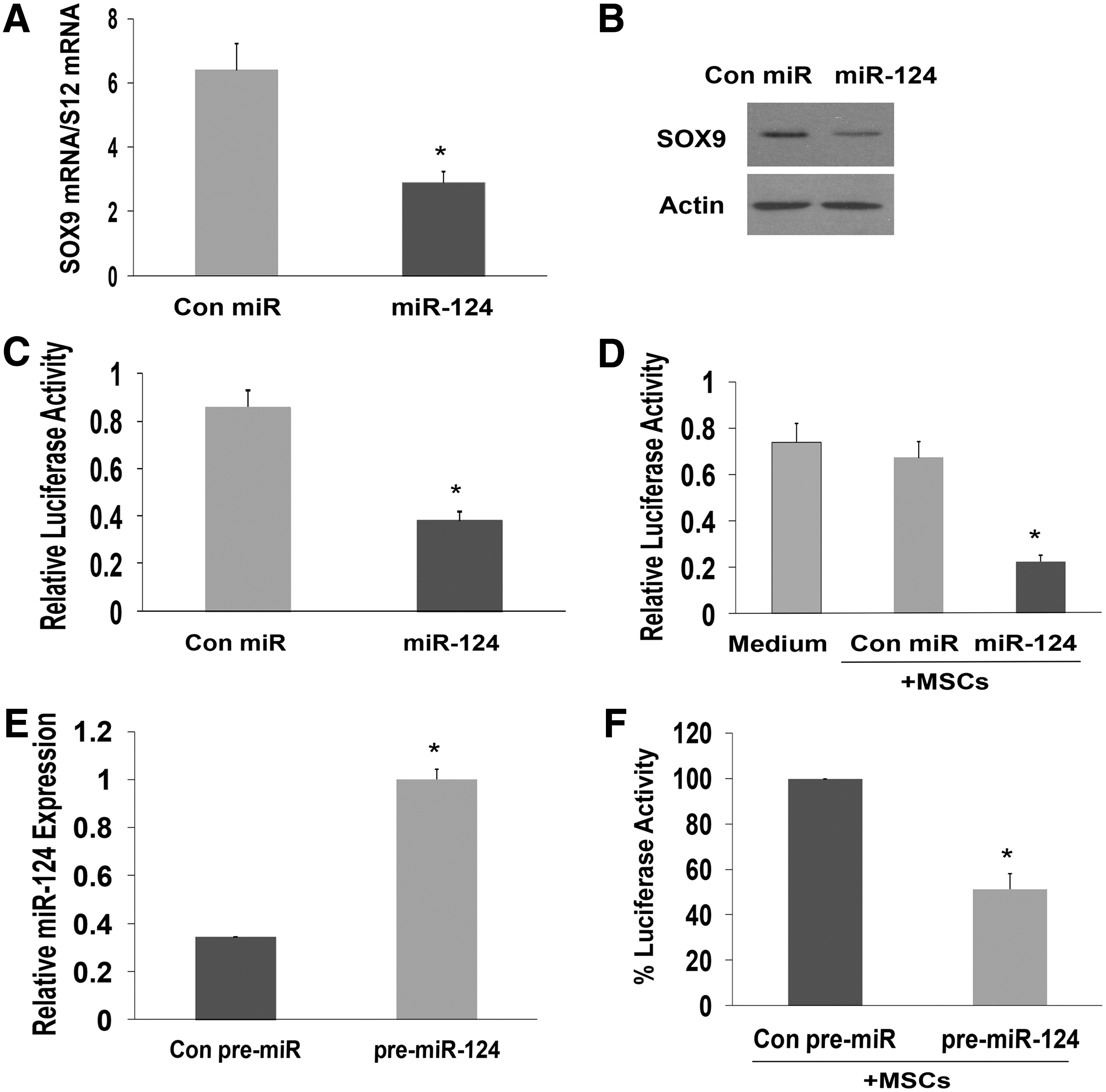
MSC-delivered miR-124 mimic downregulates the expression of Sox9 in NPCs. NPCs were transfected with miR-124 mimic and the expression of Sox9 was examined using qRT-PCR
Role of exosomes in the delivery of exogenous miR-124 by MSCs
To delineate the mechanisms by which MSCs deliver miRNA mimic to the NPCs, we examined the role of exosomes in this process. Exosomes were extracted from MSCs transfected with a control miR or with miR-124 mimic and characterized as previously described (Fig. 4A) [26]. The expression of miR-124 in the extracted exosomes was determined using RT-PCR (Fig. 4B). Treatment of miR-124 reporter-transfected NPCs with exosomes isolated from miR-124-transfected MSCs decreased the luciferase activity of the reporter plasmid by 58.8% demonstrating that this reporter can be also employed for the monitoring of miRNA delivery by exosomes (Fig. 4C). The luciferase activity of the Sox9 reporter plasmid in the NPCs was also significantly decreased by the miR-124-loaded exosomes (Fig. 4D).
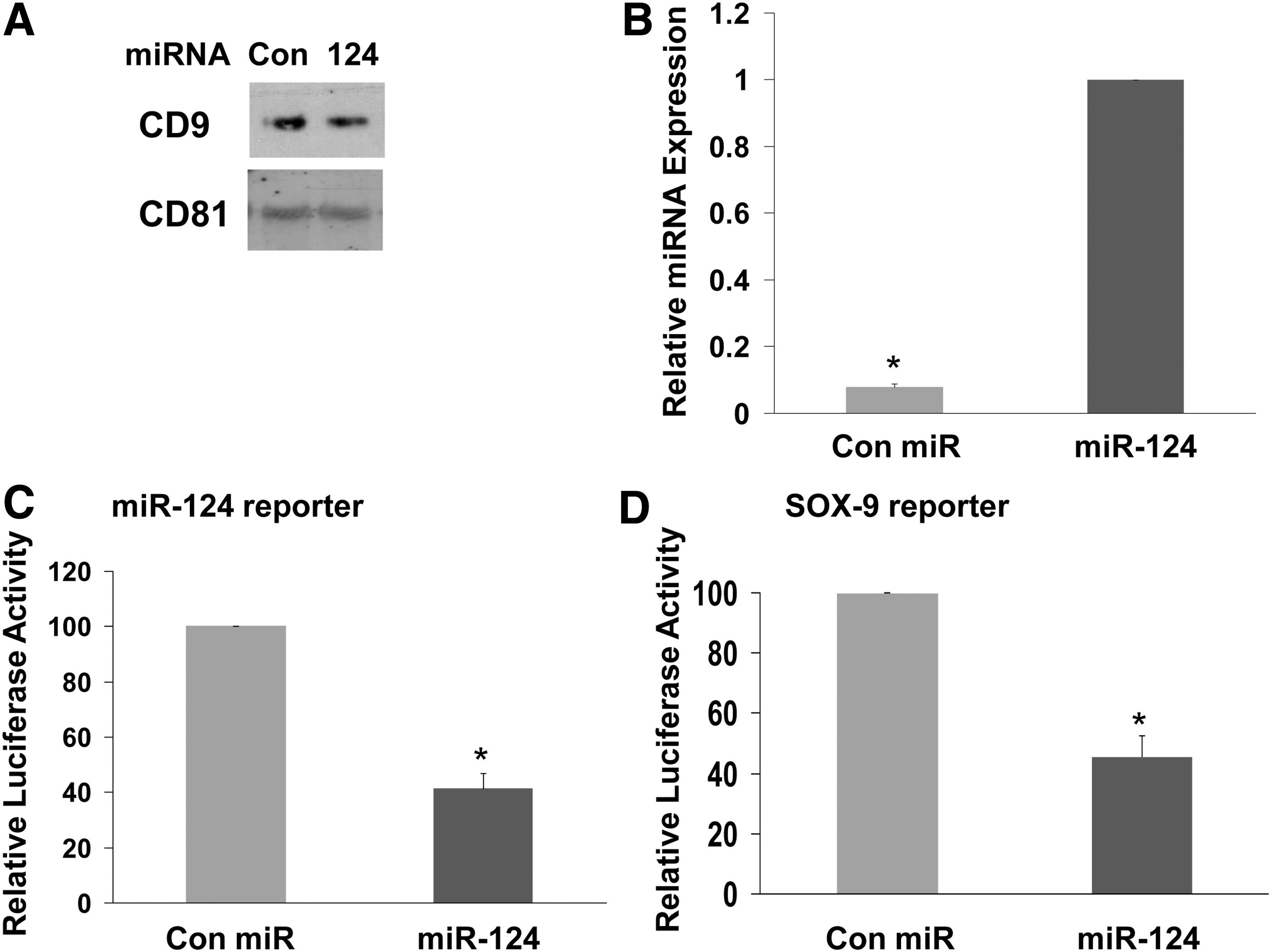
Role of exosomes in the miR-124 mimic delivery by MSCs. Exosomes were isolated from BM-MSCs transfected with either control miR or miR-124 mimic as described in the Materials and Methods section and were analyzed for the expression of CD9 and CD81 by western blot analysis
Delivery of miR-124 mimic by MSCs increases the neuronal differentiation of NPCs and the expression of glutamate transporters in NPCs and astrocytes
miR-124 has been reported to induce differentiation of NPCs in the subventricular zone [29,30]. In addition, our results demonstrated a similar effect in cultured human NPCs (Fig. 1B). We therefore examined if the delivery of miR-124 mimic by the MSCs can also induce the differentiation of the NPCs. We found that coculturing of MSCs transfected with miR-124 mimic and NPCs significantly increased the expression of β3 tubulin over that observed in NPCs cocultured with MSCs expressing a control miR (Fig. 5A) and as demonstrated for transfection of NPCs with miR-124 (Fig. 1C). We also found that the MSC-delivered miR-124 also did not affect the expression of GFAP in cocultured astrocytes (Fig. 5B). Using transwell cocultures (Fig. 5C) or isolated exosomes (Fig. 5D) of MSCs transfected with miR-124 mimic or a control miR with NPCs we found increased levels of β3 tubulin and decreased nestin expression.
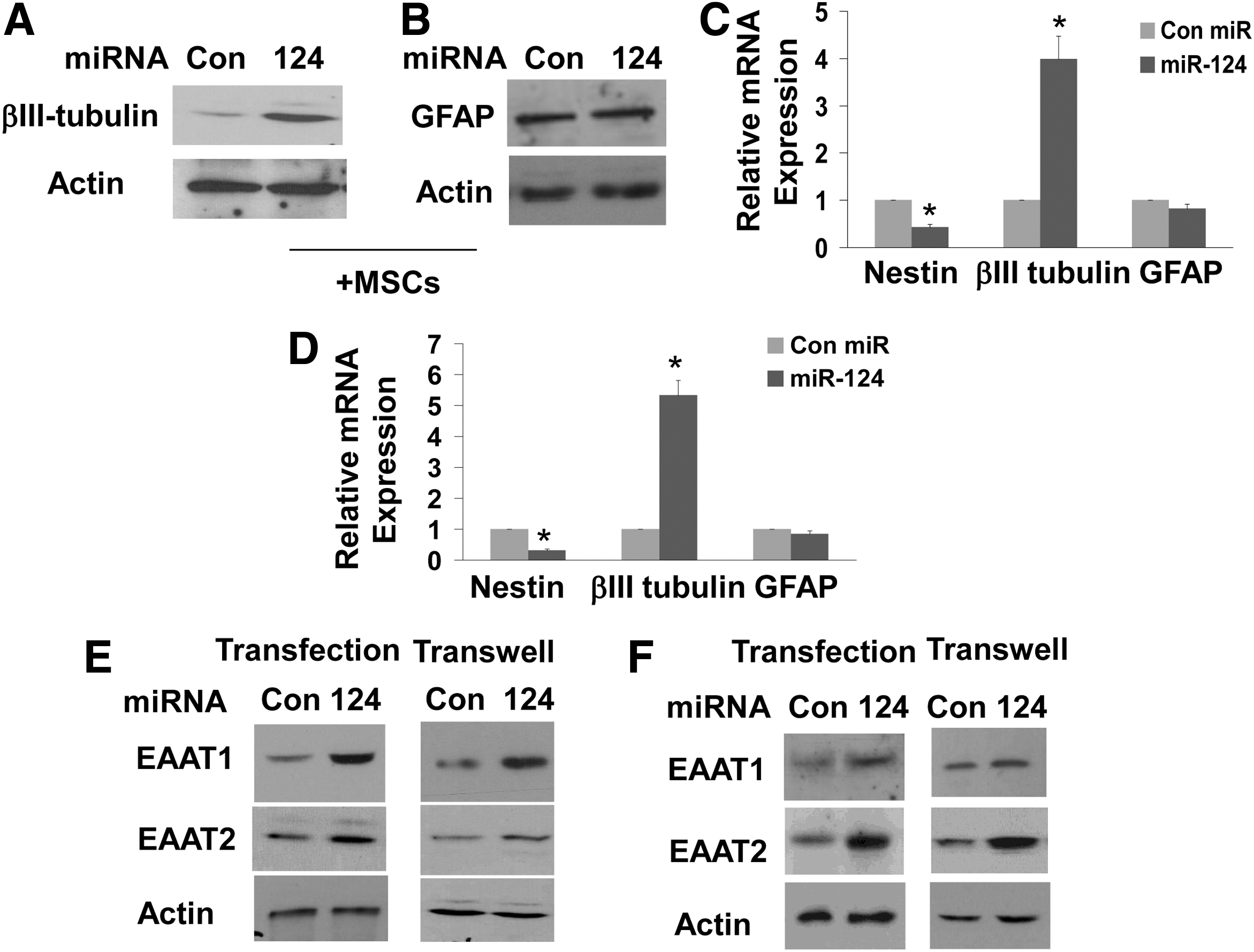
MSC-delivered miR-124 induces the differentiation of NPCs and the expression of glutamate transporters. NPCs
miR-124 has been recently reported to increase the expression of the glutamate transporter EAAT2 in mouse astrocytes [31]. Since changes in the expression of this transporter play a critical role in a variety of neurological disorders, we examined whether the delivery of miR-124 by MSCs can also increase the expression of the glutamate transporters EAAT1 and EAAT2 in human NPCs and astrocytes. We found that NPCs express both glutamate transporters EAAT1 and EAAT2 (Fig. 5E). Transfection of the NPCs with miR-124 mimic induced some increase in EAAT2 and a larger increase in EAAT1 expression (Fig. 5E). The increased expression of the glutamate transporters preceded the neural differentiation of the NPCs (data not shown). Cocultures of MSCs transfected with miR-124 with NPCs in a transwell system with 0.4 μm filter also induced a significant increase in the expression of EAAT1 and EAAT2, whereas coculture of NPCs with MSCs transfected with a control miR did not exert a significant effect (Fig. 5E). Astrocytes also expressed both EAAT1 and EAAT2 (Fig. 5F). In contrast to NPCs, both the transfection of human astrocytes with miR-124 mimic and coculture with MSCs transfected with miR-124 in a transwell system with 0.4 μm filter induced a large increase in EAAT2 and a marginal increase in EAAT1 expression in the human astrocytes (Fig. 5F).
Discussion
In this study we demonstrated the ability of adult BM-MSCs to deliver miRNA-124 to cocultured NPCs as a potential approach for the targeted delivery of miRNA-based therapy to neural cells in the CNS.
miRNAs have recently emerged as important potential therapeutic targets in neurodegenerative disorders and other pathological conditions of the CNS [15]. Indeed, changes in miRNA expression have been reported in various neurological disorders [5,11,13 –15,32 –34], and the administration of specific miRNA mimics or pre-miRNAs have demonstrated therapeutic benefits in pathological conditions in the CNS [4,35]. However, despite their promising therapeutic potential, a major drawback of the clinical application of miRNA mimics is their limited targeted delivery to neural cells.
MSCs have been reported in recent years to ameliorate various pathological conditions in the CNS including stroke, brain injury, and neuroinflammatory and neurodegenerative disorders such as multiple sclerosis, Parkinson's disease, and ALS [21,22,30]. The therapeutic effects of MSCs in both preclinical animal models and in preliminary clinical studies are attributed to their ability to migrate to areas of injury and inflammation [19], to secrete neurotrophic factors and to decrease inflammatory responses [24]. In addition, the tropism of MSCs to damaged areas has been utilized for their use as delivery vehicles for gene therapy and of various growth factors [36]. Recent studies [37], including ours [26], demonstrated that MSCs can also deliver siRNAs and miRNAs to tumor cells, both in vitro and in vivo.
We found that the BM-MSCs efficiently delivered miR-124 and miR-145 to cocultured NPCs as evident by the localization of the Cy3-labeled miRNA mimics in these cells. In addition, we demonstrated the delivery of exogenous miRNAs using novel reporter plasmids that allow the tracking of changes in miRNA levels in living cells in vitro and probably in vivo. miR-124 is a well-characterized neuronal-associated miRNA, which plays an important role in neurogenesis and differentiation of adult NPCs [38]. In addition, miR-124 has been shown to decrease the proliferation of SVZ-derived NPCs after stroke and induce their differentiation [39].
MiR-124 exerts some of its effect by targeting Sox9 [40], SCP-1 [38], Jagged-1 [41], or PTBP-1 [42]. We demonstrated that the delivered miR-124 mimic decreased the expression of Sox9, as did the transfected miR-124 mimic. Sox9 has been recently reported to be essential for the formation of multipotent NSCs and for their maintenance [43]. Thus, the downregulation of Sox9 expression by miR-124may mediate some of the miR-124 effects on the differentiation of the NPCs.
MSCs have been shown to increase neurogenesis in the subventricular zone in control and disease-model animals [29]. Moreover, in addition to enhanced neurogenesis, these cells can also induce specific differentiation of the locally residing NPCs [30]. Therefore, the delivery of miR-124 or other relevant miRNAs can further enhance the neuronal differentiation ability of the NPCs toward a specific neural phenotype.
We also found that the delivered miR-124 increased the expression of the glutamate transporters EAAT1 and EAAT2 that are expressed in the human NPCs and astrocytes, albeit to a different degree. Glutamate is the main excitatory neurotransmitter in the CNS and increased levels of this neurotransmitter have been associated with a variety of pathological conditions [44]. The glutamate transporters, EAAT1 and EAAT2, which are mainly expressed in glial cells, have been implicated in the regulation of glutamate levels in the synaptic cleft [45]. EAAT2 is a broadly expressed transporter in the brain [46], while EAAT1 is mainly expressed in the cerebellum [47]. An aberrant expression or function of these transporters has been implicated in several neurological disorders such as Alzheimer's disease [48], ALS [49], and Rett Syndrome [50]. miR-124 secreted from neuronal-derived exosomes was recently reported to mediate the increased expression of astrocytic GLT1/EAAT2 protein in an indirect yet undefined mechanism and to restore the pathological downregulation of this transporter in SOD1G93A mice [31]. Our results in human astrocytes are similar to the reported regulation of GLT1/EAAT2 by miR-124 in mouse astrocytes but in contrast to the very minor effect of miR-124 on the induction of the astrocytic EAAT1, we did find an increased expression of this transporter in NPCs. The role of the glutamate transporters in neural stem cells is not well understood, however, recent studies demonstrated basal transporter expression in these cells and further upregulation in response to injury, which decreases NSC proliferation [44].
The expression of the glutamate transporters in astrocytes has been reported to be regulated by insulin [51] and Ying-Yang1 [52] for EAAT1 and CREB and NF-kB [53], for EAAT2. The mechanisms by which miR-124 increases the expression of EAAT1 and EAAT2 in NPCs and astrocytes are currently not clear and under study. The increased expression of the two glutamate transporters by miR-124 may have important implications in pathological conditions where higher levels of glutamate are accumulated or when the expression of glutamate transporters is downregulated such as in ALS.
In addition to its important role in neuronal differentiation and glutamate transporter expression, miR-124 has been also recently associated with the inhibition of microglia and the inflammatory response in experimental autoimmune encephalomyelitis [54]. Since inflammatory responses are also associated with and contribute to the pathogenesis of stroke, brain injury, and various neurodegenerative disorders [24], the delivery of miR-124 mimic by MSCs can also enhance their anti-inflammatory effects.
Endogenous miRNAs have been shown to be transferred by MSCs via specific microvesicles and exosomes [55] and to play a role in intercellular communication and in pathological processes such as bone marrow metastases of breast cancer [56,57]. We recently found that MSCs delivered miRNA mimics to glioma cells and GSCs and that this delivery impacted the migration and self-renewal of these cells [26]. The delivery of the specific miRNA mimics by the MSCs was mediated by gap-junction-dependent and independent mechanisms and via the secretion of exosomes. We found in this study that the delivery of miR-124 mimic by the BM-MSCs to both NPCs and astrocytes was mediated by contact-independent mechanism and exosomes secretion. Our current and recent results clearly demonstrate that in addition to the physiological delivery of endogenous miRNAs, MSCs can be also employed to deliver exogenous miRNAs [26]. Although recent reports [58,59] and our recent [26] and current results demonstrate the ability of MSC-derived exosomes to deliver miRNA mimics, the use of MSCs for miRNA delivery may be superior to that of isolated exosomes, since these cells can migrate to injury, inflammation and tumor sites and therefore can provide a targeted miRNA delivery, in addition to their other therapeutic impacts.
In summary, MSCs have been implicated as potential vehicles for the delivery of growth factors and cytokines in various pathological instances in the CNS [21]. Our results indicate that they can also deliver exogenous miRNAs to NPCs and astrocytes, impact cell differentiation, and induce glutamate transporter expression similar to endogenous and transfected miRNAs. Therefore, MSCs may be employed for the delivery of miRNA-based therapy in various pathological conditions in the CNS such as inflammation, brain injury, and neurological disorders.
Footnotes
Acknowledgments
This work was supported by the William and Karen Davidson Fund, Hermelin Brain Tumor Center and by the Lori and Alan Zekelman Fund.
Author Disclosure Statement
The authors declare no conflict of interest related to this article.