
Abstract
During development of the central nervous system, trophic, together with genetic, cues dictate the balance between cellular proliferation and differentiation. Subsequent to the birth of new neurons, additional intrinsic and extrinsic signals regulate the connectivity of these cells. While a number of regulators of ventral midbrain (VM) neurogenesis and dopaminergic (DA) axon guidance are known, we identify a number of novel roles for the secreted glycoprotein, Wnt7a, in this context. We demonstrate a temporal and spatial expression of Wnt7a in the VM, indicative of roles in neurogenesis, differentiation, and axonal growth and guidance. In primary VM cultures, and validated in Wnt7a-deficient mice, we show that the early expression within the VM is important for regulating VM progenitor proliferation, cell cycle progression, and cell survival, thereby dictating the number of midbrain Nurr1 precursors and DA neurons. During early development of the midbrain DA pathways, Wnt7a promotes axonal elongation and repels DA neurites out of the midbrain. Later, Wnt7a expression in the VM midline suggests a role in preventing axonal crossing while expression in regions flanking the medial forebrain bundle (thalamus and hypothalamus) ensured appropriate trajectory of DA axons en route to their forebrain targets. We show that the effects of Wnt7a in VM development are mediated, at least in part, by the β-catenin/canonical pathways. Together, these findings identify Wnt7a as a new regulator of VM neurogenesis and DA axon growth and guidance.
Introduction
T
During VM development, Sox2-expressing progenitor cells, born within the ventricular zone, migrate ventrally into the intermediate zone where they adopt a post-mitotic Nurr-1+ DA precursor phenotype [1,2]. In mice, from embryonic day (E) 10.5 to E14.5, these Nurr1+ neuroblasts progressively arrive in the marginal zone of the VM where they adopt a mature DA identity [2,3]. Shortly after their birth, DA neurons begin to extend axons dorsally, and subsequently rostrally toward their forebrain targets, forming the nigrostriatal, mesocortical, and mesolimbic pathways [4,5]. At approximately E14.5, DA fibers stall at the ventral border of the ganglionc eminence where they increase in numbers before innervating the ventromedial and subsequently dorsolateral ganglionc eminence [6]. DA synaptogenesis continues into the first few weeks of postnatal life while aberrant sprouting and pruning of established connections continues into adult life [5,7,8]. While a crude summary, development of the VM requires a precisely orchestrated sequence of proceedings regulated by numerous intrinsic and extrinsic factors that ensure the correct birth, organization, and connectivity of the DA neurons. The past 15 years has seen significant efforts to identify and comprehend the role of these regulators; however, they have thus failed to account for or explain all DA developmental events [3,5].
In this regard, we have focused our attention on understanding the role of Wnt proteins in midbrain development. Wnts are a large family of secreted glycoproteins that play a number of important roles in development, including cellular proliferation, fate specification, differentiation, and neural connectivity. They are highly conserved diffusible glycoproteins and signal through seven-transmembrane Frizzled receptors and/or coreceptors, including the atypical tyrosine kinases Ryk and Ror2. Upon receptor binding, Wnts signaling can be transduced through the β-catenin/canonical pathway or noncanonical pathways that include the Ca2+ and planar cell polarity (PCP) pathway [9 –11].
To date, of the 19 Wnts identified in mammals, 13 are expressed within the developing midbrain [12]. Studies have identified important roles for canonical Wnt signaling (Wnt1 and Wnt2) in regulating proliferation of VM progenitors, whereas noncanonical Wnt5a signaling has been demonstrated to be integral in the differentiation of DA progenitors [10,13 –22]. More recently, we and others have additionally identified a role for Wnt5a and Wnt7b (together with the Ryk receptor) in DA axon growth and guidance [6,23,24].
However, the role of Wnt7a, a Wnt previously shown to be expressed in the VM during DA neurogenesis [12], remains to be elucidated. Elsewhere within the central nervous system (CNS), Wnt7a has been demonstrated to activate both the canonical and noncanonical Wnt signaling pathways, and therefore, not surprisingly, has been shown to have multiple and even counteracting effects on neural development. In this study, we examine the temporal and spatial expression of Wnt7a within the developing VM and, using gain and loss of function approaches, examine its role in the proliferation, differentiation, survival, and neural connectivity of DA neurons.
Materials and Methods
Animals
This study conformed to the Australian National Health and Medical Research Council's published code of practice for the use of Animals in Research, and the experiments were approved by the Florey Institute of Neuroscience and Mental Health animal ethics committee. Embryos were isolated from time-mated Sprague–Dawley rats and CD1 mice. Animals were time-mated overnight, and visualization of a vaginal plug the following morning was taken as E0.5. Wnt7A-deficient mouse embryos and their littermate wild-type controls were collected at E14.5 and postnatal day (P) 1 [25].
Quantitative real-time polymerase chain reaction
Quantitative real-time polymerase chain reaction (Q-PCR) was performed to examine the temporal expression of Wnt7a in the developing midbrain DA pathways. The ventral and dorsal midbrain was isolated from mice at E10.5, E11.5, E14.5, P1, and adult. The developing lateral ganglionic eminence (or equivalent adult striatum) was isolated from E14.5, P1, and P70 mice. For each developmental stage (E10.5, E11.5, and E14.5), tissue was pooled from four independent mouse litters. P1 and adult tissue was isolated from 4 to 5 animals.
Following tissue isolation, total RNA was extracted using the RNAeasy Micro kit (Qiagen). RNA was reverse transcribed using Superscript III First-Strand Synthesis supermix for qRT-PCR (Invitrogen), and QPCR was carried out using the SYBR GreenER™ qPCR SuperMix Universal (Invitrogen) on an ABI7700 sequence detection system (Applied Biosystems) using the comparative ΔΔCT method [26]. Oligonucleotide sequences were as follows:
HPRT forward, 5′-CTTTGCTGACCTGCTGGATT-3′
HPRT reverse, 5′-TATGTCCCCCGTTGACTGAT-3′
Wnt7a forward, 5′-TGCGTGCCAGTCGAAACAAG-3′
Wnt7a reverse, 5′-GATATACACCAGGTCAGTGTCCATGG-3′
CyclinD1 forward, 5′-CAC AAC GCA CTT TCT TTC CA-3′
CyclinD1 reverse, 5′-CTT GGG ATC GAT GTT CTG CT-3′
p27 forward, 5′-GGG TCT CAG GCA AAC TCT GA-3′
p27 reverse, 5′-TCT GTT GGC CCT TTT GTT TT-3′
p57 forward, 5′-CGA GAA TCA GGA GCT GAA GG-3′
p57 reverse, 5′-GTT CTC CTG CGC AGT TCT CT-3′
In situ hybridization and immunohistochemistry in vivo
Embryos were isolated in ice-cold phosphate-buffered saline (PBS), fixed overnight in 4% paraformaldehyde, followed by overnight immersion in 30% sucrose in PBS. E11.5, E14.5, and P1 embryos were cryosectioned on a coronal or sagittal plane at a thickness of 16 μm. In situ hybridization (ISH) was performed as previously described [1], using a DIG-labeled single-stranded RNA probe for Wnt7a (a kind gift from Prof. A. McMahon, Harvard University). The specificity of the probe was validated in Wnt7a-deficient tissue. Following ISH, the tissue was again fixed using 4% paraformaldehyde before immunohistochemistry for tyrosine hydroxylase (TH; the rate-limiting enzyme in DA synthesis and marker of DA neurons and neurites).
Immunohistochemistry was performed on 4% paraformaldehyde-fixed cultures and slides as previously described [23,27]. The following primary antibodies were used: mouse anti-TH (1:800; Chemicon), sheep anti-TH (1:500; PelFreez), mouse anti-βIII-tubulin (1:1,500; Promega), rabbit anti-Nurr1 (1:200; Santa Cruz), goat anti-Sox2 (1:200; R&D Systems), rabbit anti-Ki67 (1:1,000; Neomarkers), and cleaved caspase-3 (Asp175) (1:100; Cell Signaling Technology). Appropriate fluorophore-conjugated (DyLight 488, 549, 694) secondary antibodies or biotinylated secondary antibody together with the Vector laboratories ABC immunoperoxidase kit were used for visualization. TUNEL staining was performed according to the manufacturer's guidelines (DeadEnd™ Fluorometric TUNEL System; Promega). All cultures were counterstained with Hoechst 33258 for 10 min (1:1,000; Invitrogen).
Purification of HA-tagged Wnt7a
Rat B1A fibroblasts stably overexpressing hemagglutinin-tagged Wnt7A (a kind gift from Professor Ernest Arenas, Karolinska Institute) were generated as previously described [17]. B1A-control or B1A-Wnt7a cells were expanded in Dulbecco's modified Eagle's medium (DMEM), 10% fetal bovine serum (FBS),
Immunoblotting and activity of Wnt7a
The presence of Wnt7a within the conditioned media (CM) was verified by immunoblotting for Wnt7a. The activity of Wnt7a CM was tested on the DA cell line, SN4741. The SN4741 cells were expanded in the presence of DMEM, 10% FBS,
VM primary cultures
The VM of E13.5 and E16.5 rat embryos (approximately equivalent to mouse E11.5 and E14.5, respectively) was microdissected in chilled L15 media (Invitrogen). The isolated VM was enzymatically dissociated in Hank's balanced salt solution containing 0.075% trypsin and 0.1% DNase for 12–15 min at 37°C. Cells were subsequently resuspended in serum-free N2 media and seeded at a density of 180,000 cells/cm2 at 37°C, 5% CO2 for 72 h. Five percent B1A or Wnt7A condition media (±Dkk-1, 500 ng/mL) was added to the wells before seeding the VM cells. Cells were fixed with 4% paraformaldehyde for 20 min before immunocytochemistry.
Cells immunoreactive for TH, βIII-tubulin, Nurr1, Ki67, TUNEL, and stained with Hoechst were counted from 10 fields of view in three parallel wells per experiment. Counts were obtained from 3 to 4 independent cultures performed on separate days. Neurite morphology of DA neurons [tyrosine hydroxylase-immunoreactive (TH+)] and non-DA neurons (TH−/TUJ+) within the cultures were analyzed as previously described [6]. Assessments included the number of neurites per neuron, the number of neurite branches, length of dominant neurite (axon), and total length of all neurites per neuron.
Analysis of Wnt7a embryos
The total number of Sox2, Nurr1, and TH-ir cells within the VM was quantified from E14.5 and P1 mice. All Wnt7a−/− embryos were compared with wild-type Wnt7a+/+ littermates with 4–6 embryos analyzed per genotype at each time point. Changes within the midbrain DA pathways in Wnt7a+/+ and Wnt7a−/− were observed by chromogenic staining for the TH+ neurons and quantified using Stereoinvestigator software (MicrobrightField) on a Leica DML microscope. At E14.5 and P1, the volume of the medial forebrain bundle (MFB) was estimated by delineating the area of the TH+ fibre bundle in the first section rostral to the midbrain TH+ neurons until the final section before the arrival of TH+ fibres in the striatum. The MFB was delineated in approximately seven sections from each brain, with the area and total volume estimated using Stereoinvestigator software. The density of TH+ fibres in the MFB was also assessed at E14.5 and P1 as described previously [6]. The density of TH+ fibres in the MFB at P1 was assessed at two independent levels: (i) 320 μm and (ii) 800 μm rostral to the midbrain DA neurons. In E14.5 embryos, the density of TH-ir fibres was only assessed at 320 μm rostral to midbrain DA neurons. At P1, the density of TH-ir varicosities in the striatum was also assessed using Stereoinvestigator software. TH+ varicosities in the ventral 350 μm of the striatum were counted from 16 μm serial sections, 1:10 series, with four sections sampled from each striatum [6]. For TH+ fibre counts and TH+ varicosity numbers, the coefficients of error and coefficients of variance were calculated as estimates of precision, and values of <0.1 were accepted.
Statistical analysis
Student's t-test or one-way analysis of variance with the Tukey post hoc test was used to identify statistically significant changes. Data represent mean±standard error of the mean unless stated otherwise.
Results
Wnt7a is spatially and temporally expressed within the developing VM and nigrostriatal pathway
To assess the potential roles of Wnt7a in midbrain development, we examined its temporal and spatial expression within the developing VM, along the DA pathways, and within the target striatum using ISH and quantitative PCR.
In mice at E11.5, a period during DA neurogenesis and corresponding to the onset of DA axonal growth, Wnt7a was expressed in the ventricular zone (Fig. 1B, C, E, F) and the intermediate zone (Fig. 1C, F) of the VM, but not in the mantle zone (Fig. 1C′, C′′, F′, F′′) where DA neurons reside. Later in development, E14.5, corresponding to the end of DA neurogenesis and the timing of axonal arrival at the ventral border of the striatum, Wnt7a expression was maintained within the VM overlapping with DA neurons (Fig. 1H, H′, H′′) and showed stronger expression surrounding the midline (Fig. 1H). Sagittal sections revealed that Wnt7a was also expressed within the hypothalamus and thalamus, adjacent to the TH+ MFB (Fig. 1K, K′′, arrowheads), as well as throughout the developing medial and lateral ganglionic eminences (LGE)/striatum (Fig. 1I, K). By E16.5, Wnt7a expression within the striatum opposed TH expression, with DA fibers present in the ventral striatum (Fig. 1L), whereas Wnt7a expression was observed in the dorsal striatum and surrounding the lateral ventricle (Fig. 1L′).
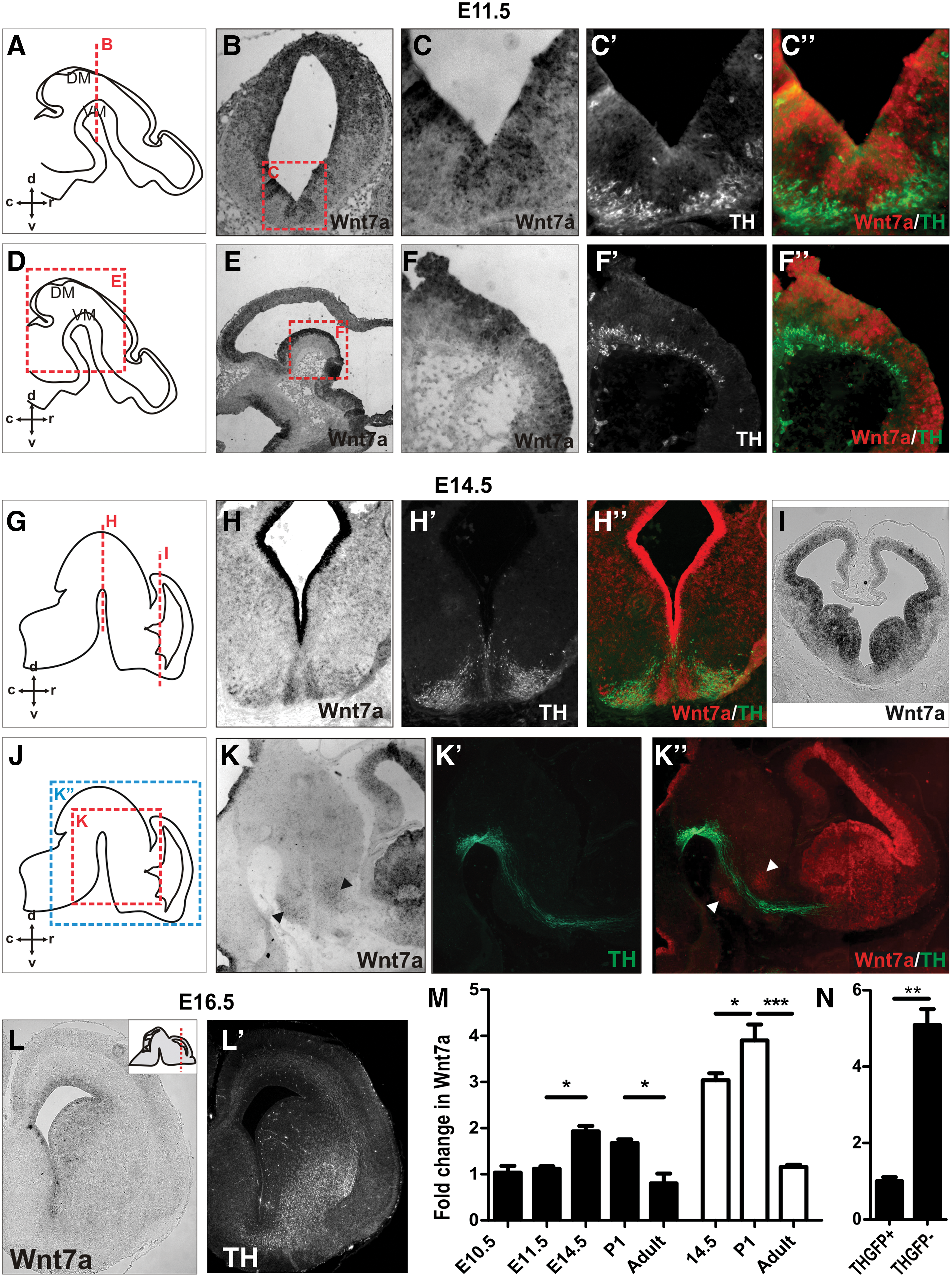
Wnt7a shows a temporal and spatial expression suggestive of a role in ventral midbrain (VM) neurogenesis and development of dopamine (DA) pathways.
Quantitative PCR was performed to quantify and verify the temporal expression of Wnt7a in the VM and LGE/striatum of mice. VM tissue was isolated at E10.5 (the onset of DA neurogenesis), E11.5 and E14.5 (periods of DA neurogenesis and axonal morphogenesis), P1 (late DA development/synaptogenesis), and adult. Within the VM, Wnt7a was significantly upregulated at E14.5 and subsequently downregulated postnatally (from P1 to adult; Fig. 1M). LGE tissue was isolated at E14.5 (corresponding to the time of arrival of DA axons), P1 (DA synaptogenesis), and adult. Wnt7a showed a two-fold increase in expression in the LGE at E14.5 compared with VM, a level that was further elevated in P1 tissue before downregulation in the adult brain (Fig. 1M).
Wnt7a mRNA expression was more closely examined in VM cells isolated by fluorescence-activated cell sorting from the TH-GFP reporter mouse [29]. Q-PCR revealed a fivefold increase in Wnt7a mRNA expression in the GFP− cell fraction (TH− non-DA cells within the VM) compared to the GFP+ fraction (TH+ DA neurons; Fig. 1N).
Wnt7a CM activated the canonical/β-catenin pathway within DA neurons
Next, the effect of Wnt7a on VM cells was examined. Due to the lack of a reliable, functional commercially available Wnt7a recombinant protein, we partially purified Wnt7a CM from stably transfected fibroblast cell lines engineered to secrete Wnt7a. CM was also partially purified from mock-transfected fibroblast cells. The CM was concentrated (∼250-fold) and partially purified by size exclusion-based filtration. The presence of Wnt7a within the CM was verified by immunoblotting using an anti-Wnt7a antibody (Fig. 2A). To confirm that the Wnt7a-CM was biologically active, we examined its ability to activate Wnt signaling in a DA cell line (SN4741). Wnt7-CM, but not mock-CM, induced disheveled-2 phorphorylation (P-Dvl2), which was visible as a motility shift of the protein (Fig. 2B) [30]. Within the CNS, the role of Wnt7a has largely been attributed to canonical Wnt signaling, hence, we examined the ability of Wnt7a-CM to stabilize β-catenin (a key mediator of this pathway) within a DA cell line. Wnt7a-CM treatment of SN4741 cells resulted in an increase in active (dephosphorylated) β-catenin (ABC), an effect that could be abrogated by the addition of the canonical pathway inhibitor, Dkk (Fig. 2B).
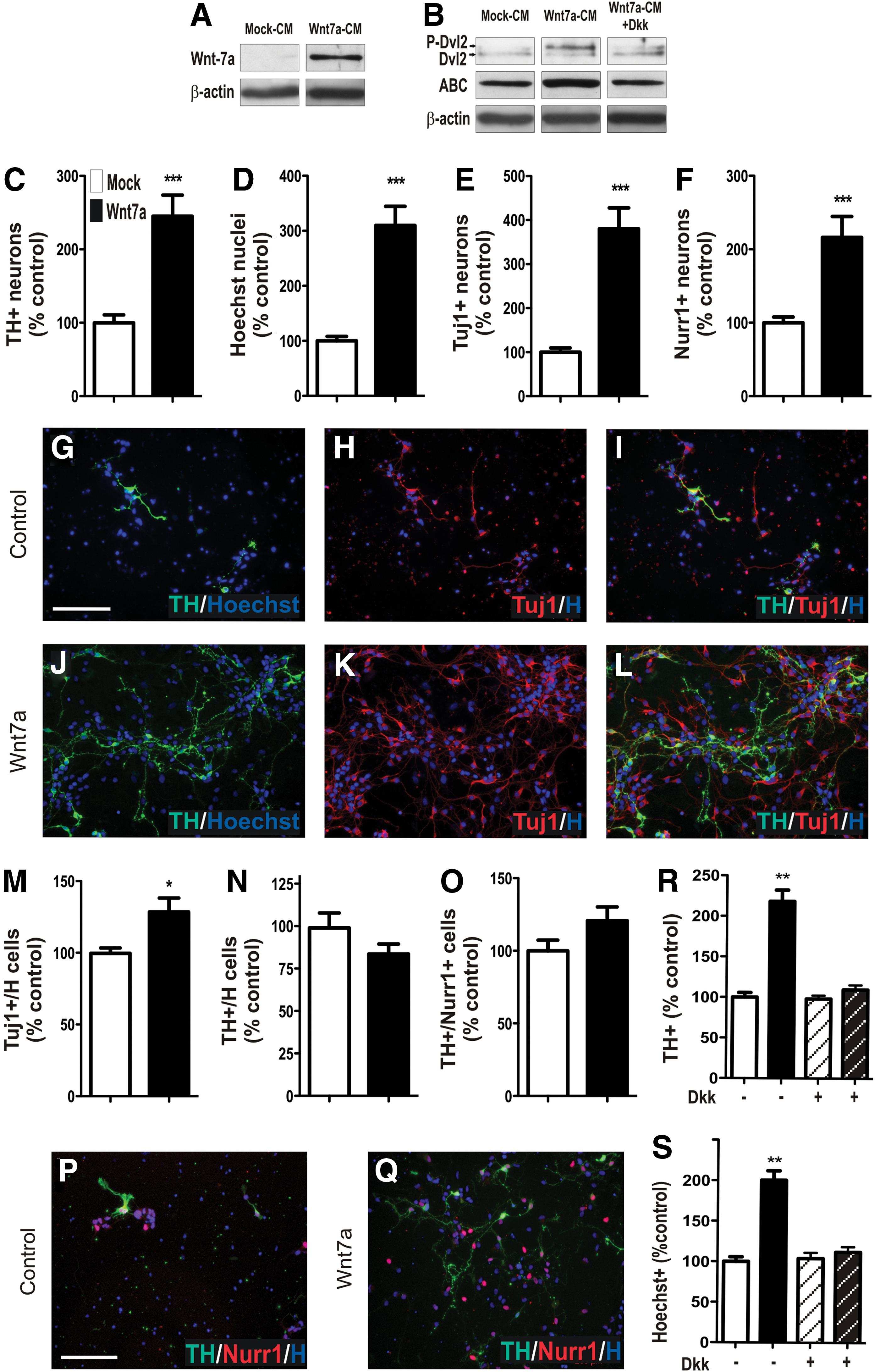
Active Wnt7a influences DA neurogenesis by regulating VM neuronal number.
Wnt7a increases the neuronal pool and consequently DA neurons in culture
To examine the function of Wnt7a in VM DA development, rat E13.5 VM primary cultures were treated with Wnt7a-CM (or mock-CM). Wnt7a resulted in robust (138%) increase in number of TH+ cells compared with mock-CM (Fig. 2C, G, J). To ascertain whether the increase in TH+ DA neurons was a consequence of VM progenitor proliferation, differentiation, and/or cell death, total cell (Hoechst), neuronal (TUJ), and DA progenitor (Nurr1) counts were performed. Wnt7a-CM resulted in a 217% increase in the total number of cells in culture (Fig. 2D, G–L), a 280% increase in the number of Tuj+ neurons (Fig. 2E, H, K), and 116% increase in Nurr1+ DA precursors (Fig. 2F, P, Q). We show that Wnt7a promoted neuronal differentiation, with a significant (24%) increase in TUJ/H cells (Fig. 2M), however, had no effect on the proportion of TH/H cells (Fig. 2N) or TH/Nurr1 (Fig. 2O), indicating that the increase in DA progenitors and DA neurons was a consequent of an increase in total neurons in culture.
Elsewhere within the developing CNS, the effects of Wnt7a have largely been attributed to signaling through the β-catenin/canonical Wnt pathway. Hence, to confirm the effects of Wnt7a on VM cultures and to characterize the pathway mediating these responses, we employed an antagonist of the canonical pathway, Dkk1, to prevent Wnt7a's interaction with the Fz/LRP receptor complex [31]. Dkk1 in the presence of Mock-CM had no effect on TH (Fig. 2R) or Hoechst (Fig. 2S) cell counts, indicating that the compound at these doses was not toxic to the cells. In the presence of Wnt7a-CM, Dkk-1 was capable of ablating the previously observed Wnt7a-induced increases in total cell and TH cell numbers (Fig. 2R, S). These findings demonstrate the specificity of Wnt7a on VM cells and identify canonical Wnt signaling in mediating these effects.
Wnt7a influences survival and regulates cell cycle progression within the VM
We examined survival, proliferation, and cell cycle progression in VM primary cultures in an effort to identify the mechanism through which Wnt7a increased neuronal numbers. We observed a significant (74%) decrease in the number of TUNEL+ apoptotic cells in cultures treated with Wnt7a, suggesting a neuroprotective role for the protein (Fig. 3A–C). To determine whether cellular proliferation also contributed, we assessed the number of Ki67+ cells as well as genes involved in cell cycle progression. We observed a notable (49%, P=0.054) increase in Ki67+ cells. Surprisingly, a significant increase in p57, moderate increase in p27, and decrease in cyclinD1 (Fig. 3G–I), downstream Wnt/β-catenin target genes, were also observed. Collectively, these findings suggest that the increased rate of proliferation, accompanied by early cell cycle exit, may explain the increased TUJ+ neuronal differentiation observed within these cultures.
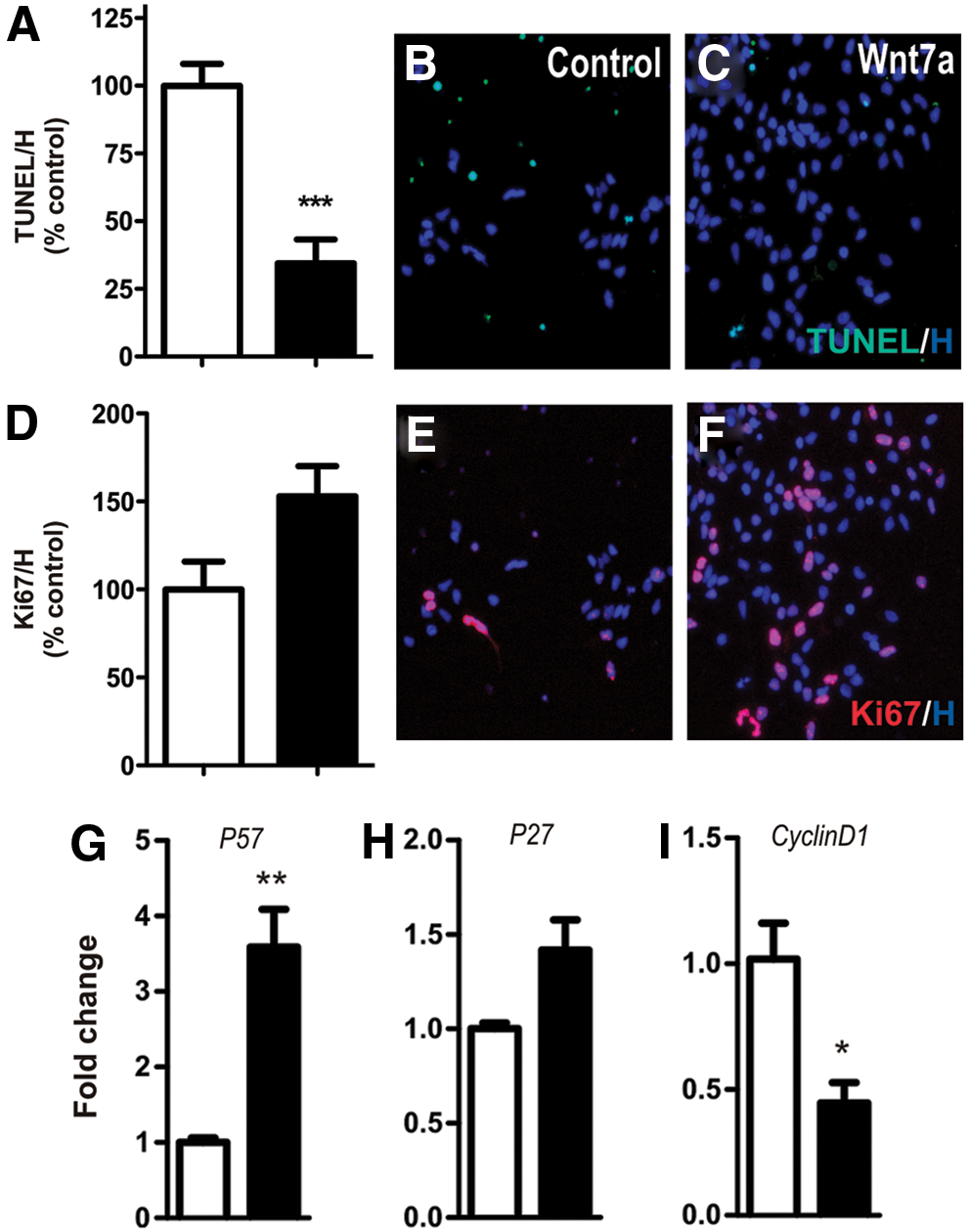
Wnt7a regulates VM progenitor proliferation and survival.
While Wnt7a demonstrated roles in the survival and cell cycle progression within the VM, examination of cultures at later stages of development (E16.5) revealed that these effects were specific to early (E13.5) development. At E16.5, Wnt7a had no significant effect on the total number of Hoechst, Tuj, Nurr1, or TH-immunoreactive cells (Supplementary Fig. S1; Supplementary Data are available online at
Decreased neuronal progenitors, DA precursors, and DA neurons in Wnt7a-deficient mice
To confirm the observed roles of Wnt7a in VM development, we examined Wnt7a-deficient mice at E14.5 and P1. Compared with wild-type littermate controls, at E14.5, Wnt7a-null mice displayed a significant decrease in Sox2+ progenitors within the ventricular zone (25.0%, Fig. 4A, E, I), Nurr1+ precursors in the intermediate zone (25.2%, Fig. 4B, F, J), and consequently, TH+ neurons within the marginal zone (19.3%, Fig. 4C, G, K). No change was observed in Caspase-3+ cells within the VM (data not shown). In support of in vitro findings, by later stages of development (postnatal day 1), TH+ cell numbers (Fig. 4D) as well as Sox2 and Nurr1 (data not shown) within the VM were not significantly different from wild-type littermates, suggesting that compensatory mechanisms were able to correct for earlier observed defects.
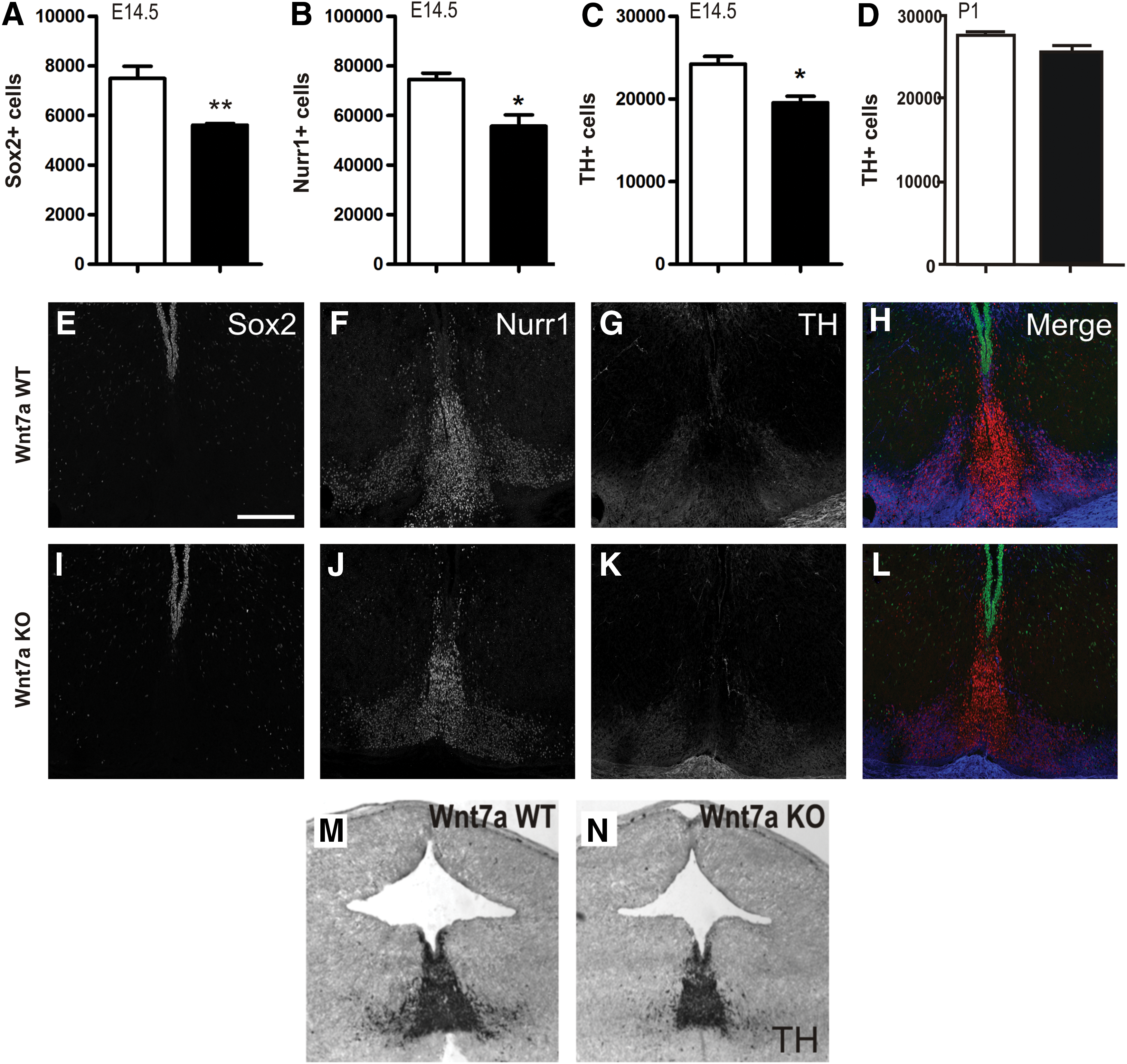
A reduction in proliferating progenitors in Wnt7a-deficient mice results in reduced postmitotic DA precursors and neurons in the VM.
Wnt7a temporally regulated DA axon morphogenesis
Wnt7a expression within the VM was observed to extend beyond the period of DA neurogenesis and showed both temporal and spatial expression suggestive of a role in the development of the midbrain DA pathways (Fig. 1). We therefore examined the effect of Wnt7a-CM (or Mock-CM) on the growth and guidance of DA neurons (TH+) as well as other non-DA neurons (TH−/TUJ+) within VM primary cultures. Wnt7a treatment of E13.5 VM cultures had no effect on the number of neurites per DA neuron (Fig. 5A); however, neuritic branching was significantly increased (87%, Fig. 5B). More notable, Wnt7a enhanced total neurite length (112%, Fig. 5C, E, F) and promoted dominant neurite (axonal) elongation (108%, Fig. 5D–F), compared with control-treated cultures. Examination of non-DA neurons in culture (ie, TH−/TUJ+) showed no changes in neurite morphology, indicating the effects of Wnt7a were selective for DA neurons.
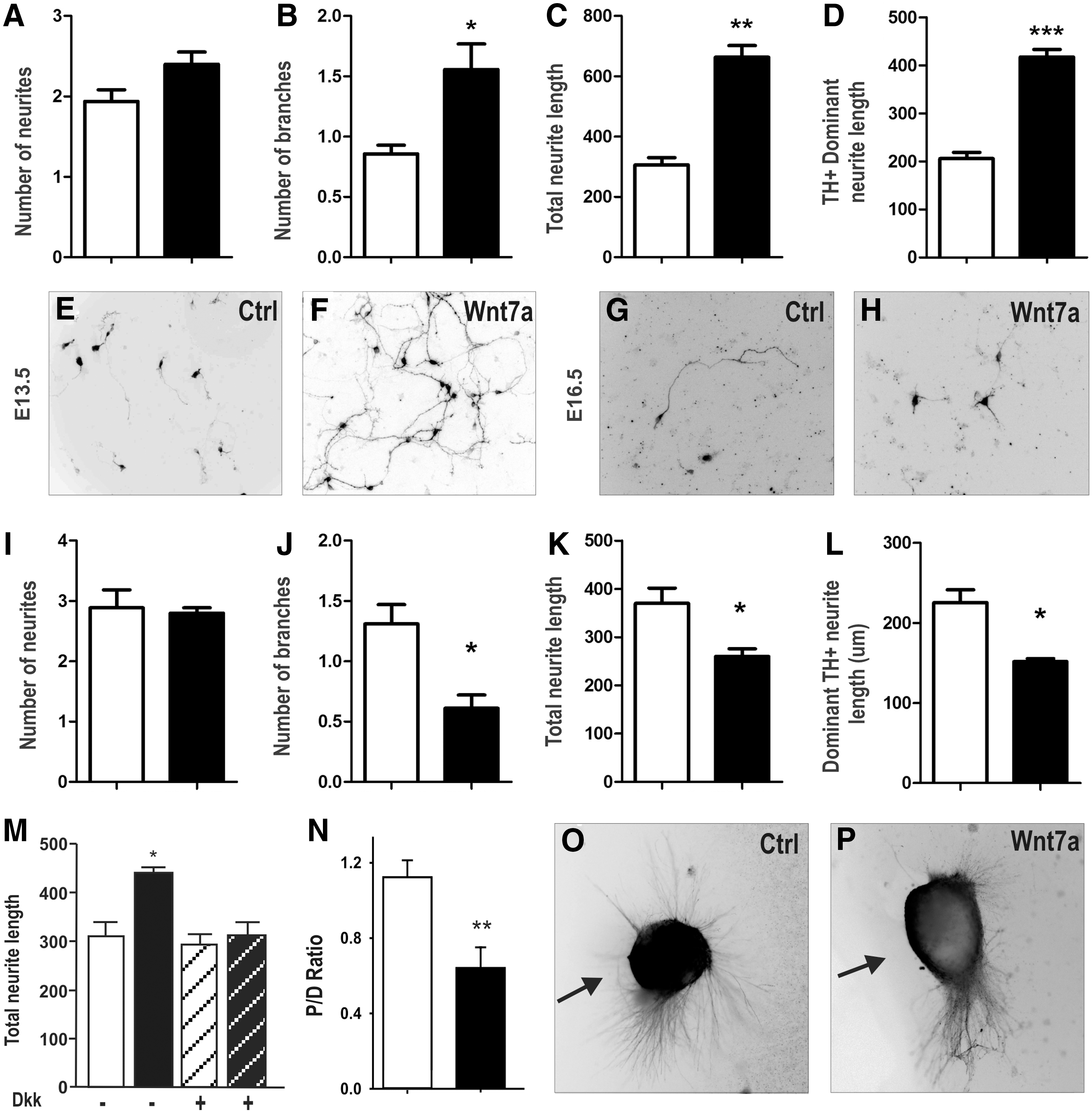
Wnt7a regulates DA axonal growth and guidance in vitro, effects mediated by the β-catenin/canonical pathway.
We recently identified dual roles for Wnt5a in axonal growth, promoting neurite extension in young DA neurons while inhibiting growth in cultures from older embryos [6]. We therefore examined the effects of Wnt7a on older (E16.5) VM primary cultures. Similar to Wnt5a, both total neurite length and axonal length were significantly reduced compared with control-treated cultures (Fig. 5G, H, K, L). Furthermore, Wnt7a treatment also reversed the effect on neurite branching, and continued to have no impact on neurite number (Fig. 5I, J). Collectively, these results indicate that Wnt7a differentially regulates DA neurite growth and morphology during both early and later stages of development. The specificity of Wnt7a on DA neurites was confirmed by cotreatment with Dkk1. Dkk abolished the effects of Wnt7a on total TH neurite length (Fig. 5M), TH dominant neurite length, and neurite branching (data not shown) in E13.5 VM cultures, suggesting that the action of Wnt7a in DA neurite morphogenesis was mediated by canonical Wnt signaling and provides the first evidence of canonical Wnt signaling in DA axon morphogenesis.
Wnt7a provides chemorepulsive cues for developing DA neurites
Next, we examined whether Wnt7a provided chemotaxic cues for developing DA axons. We isolated VM explants from E13.5 and E16.5 rat embryos and positioned them in close proximity to cell aggregates overexpressing Wnt7a- (or mock-) transfected cell aggregates. After 96 h in culture, we assess the number of TH+ neurites emanating from the proximal verses distal side of the explants, relative to the Wnt7a or Mock-cell aggregate, and expressed the data as a proximal to distal ratio [6]. At both early (Fig. 5N–P) and late (data not shown) stages of development, VM explants cocultured with mock-transfected aggregates showed TH neurites emanating from all aspects of the explant, showing a proximal to distal ratio close to 1 (1.13±0.07, n=23). By contrast, VM explants cultured adjacent to Wnt7a overexpressing aggregates showed the majority of neurite emanating from the distal side of the explant (P:D=0.62±0.10, n=20), indicative of a chemorepulsive effect. Furthermore, staining for total neurites (TUJ+) radiating from the explant revealed that the chemorepulsive effects of Wnt7a were specific to DA neurites, with TUJ neurites maintaining an even distribution surrounding the explant whileTH neurites were repelled (data not shown).
Ablation of Wnt7a causes defasiculation of the MFB
Finally, we examined the phenotypic consequence of Wnt7a ablation on the development of the midbrain DA pathways in Wnt7a-deficient mice. Mice were examined at P1, a time when the DA pathway is established. Stereological quantification of the TH+ fiber bundle volume revealed that the Wnt7a−/− mice possessed a significantly enlarged MFB (0.258 μm3±0.017×107 μm3) compared with wild-type (0.191±0.011 μm3) littermates (Fig. 6A, E, F), indicative of defasiculation of the axons within the bundle. Further quantification revealed no significant difference in the number of TH+ fibers within the MFB (Fig. 6B) or the density of TH+ terminal in the ventral striatum (Fig. 6C), suggesting that while evident roles for Wnt7a were observed in DA axon growth and guidance in vitro, these functions could be compensated for in vivo.
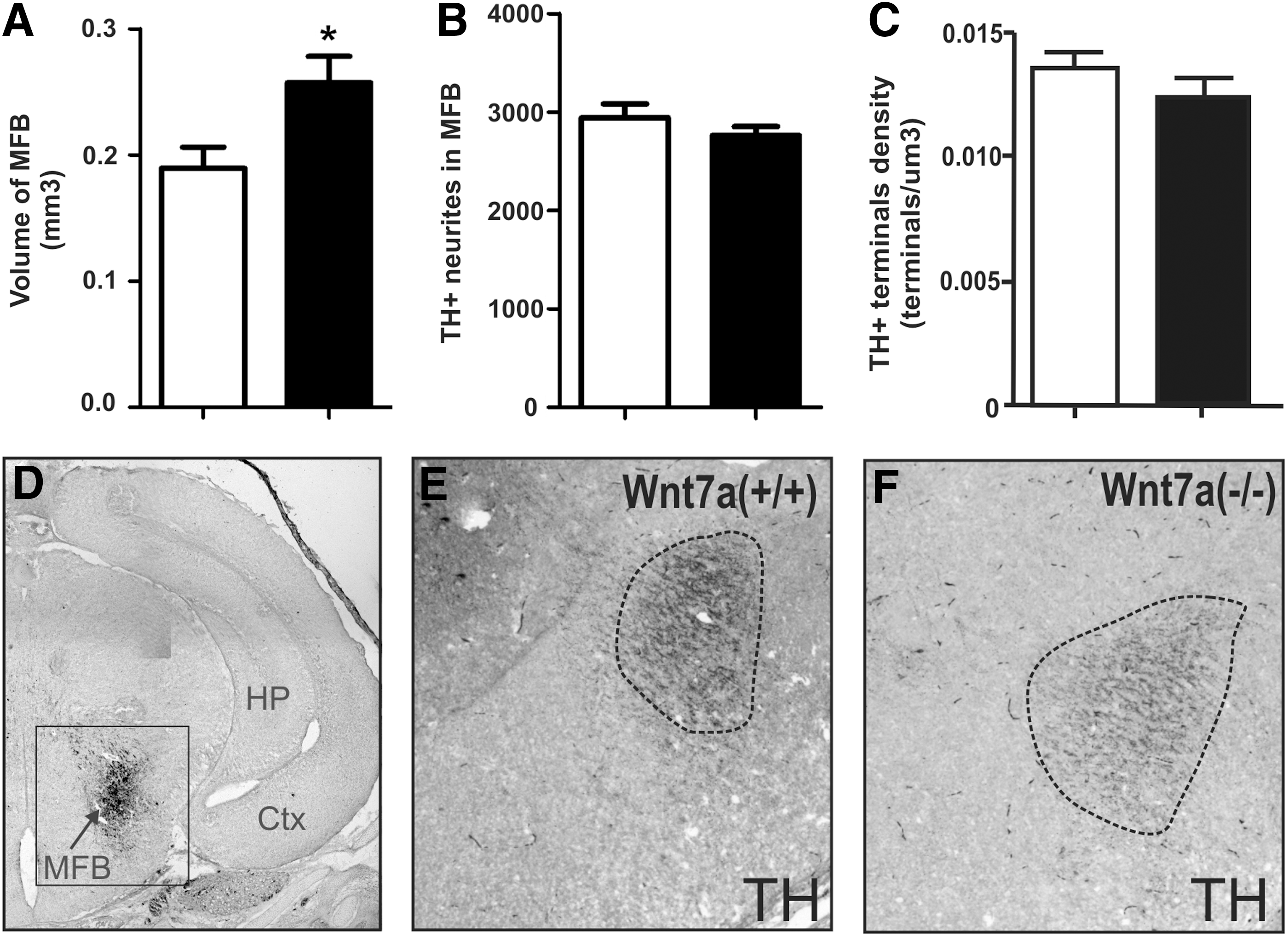
Wnt7a-deficient mice show defasiculation of the medial forebrain bundle, yet maintain appropriate DA axonal trajectory and striatal innervation.
Discussion
During development, a precise interplay between cellular proliferation and differentiation dictates the size of each nuclei within the central nervous system. This fate decision is determined by a complex interplay of both intrinsic and extrinsic factors. Upon exiting cell cycle, establishing a final resting place and adopting a terminally differentiated state, neurons respond to an additional wave of signals that dictate their neuronal connectivity. While a number of these cues in early and late development of VM neurons have been identified, they fail to account for all events. In the present study, we identify a number of novel roles for Wnt7a in VM development and the connectivity of DA axons.
Wnt7a influences cell cycle and neuronal number within the VM
We speculate that the early expression of Wnt7a within the ventricular and intermediate zones of the VM may play important roles in the regulation of progenitor proliferation and determining cell cycle exit, respectively. In support, our E11.5 VM cultures treated with Wnt7a resulted in a significant increase in total cells and marked increase in Ki67+ proliferating progenitors. Additionally, we observed an increase in total TUJ+ neurons (and consequential Nurr1+ precursors and TH+ DA neurons). Further examination suggested that the increased neuronal numbers in culture were the consequence of early cell cycle exit of VM progenitors, as supported by the elevated levels of cell cycle exit genes p27 and p57, in conjunction with a decrease in the cell cycle progression gene, cyclinD1. While Wnt7a increased the number of Nurr1+ DA precursors and mature TH+ DA neurons in culture, these effects were attributed to the influences of the glycoprotein on progenitor proliferation and as well as acquisition of an early neuronal fate, rather than DA differentiation (TH+/H and TH+/Nurr1+ remaining unchanged). Complementary to these findings, Wnt7a-deficient mice displayed a decrease in Sox2+ neural progenitors, resulting in a reduction in DA precursor and mature DA neurons in vivo. Interestingly, elsewhere in the central nervous system, Wnt7a has been shown to regulate neural progenitor proliferation and dictates cell cycle progression, thereby delaying neuronal maturation in cortical, hippocampal, olfactory bulb, and subventricular zone development [32 –34]. By contrast, others have reported Wnt7a's role in promoting neuronal differentiation [35], suggesting that critical timing and various complex functions mediate the balanced roles of Wnt7a in proliferation to differentiation progression. Here, our findings suggest an ongoing interplay between these two mechanisms, reflective of the asynchronous nature of neuronal development. We propose that Wnt7a influences the proliferation of progenitors within the ventricular zone of the VM as well as subsequently affecting their cell cycle exit within the intermediate zone, thereby dictating nuclei size and the rate of DA differentiation (Fig. 7).

Schematic representation illustrating the role of Wnt7a in VM neurogenesis. During VM development, the presence of Wnt7a within the ventricular zone (VZ, green) increases the proliferation of Sox2+ progenitors. The increased progenitor pool results in increased cells that enter the intermediate zone (IZ, blue) where Wnt7a influences cell cycle exit, resulting in an increase in postmitotic neuroblasts. The increased precursor pool subsequently results in an increase in mature neurons, including DA neurons, within the marginal zone (MZ, purple). Within this zone, Wnt7a plays a neuroprotective role and influences the initiation of DA neurite outgrowth and repulsion toward forebrain targets. Color images available online at
We demonstrate that Wnt7a additionally promoted cell survival within the VM (Fig. 3), thereby contributing to the increase in total cells, and consequently DA neurons in culture. Treatment of the midbrain DA cell line confirmed the ability of Wnt7a to protect midbrain DA neurons and demonstrated (by way of Dkk co-administration) that the mode of action was via the activation of the canonical Wnt pathway. In support, the loss of Wnt/β-catenin-TCF signaling in a number of neuronal populations has been demonstrated to increase the vulnerability of neurons to apoptotic death. More recent studies have even suggested that defects within this pathway could contribute to these pathogeneses of a number of neurodegenerative disorders, including autism, schizophrenia, Alzheimer's disease and Parkinson's disease (PD) [36,37]. Of notable relevance, the work of Marchetti has defined a novel Wnt1/Frizzled-1/β-catenin astrocyte-DA neuroprotective loop within the VM and links to neuroprotection in in vitro and in vivo models of PD [38,39]. While in vitro we demonstrate a complementary neuroprotective role for Wnt7a within the developing VM, no change in caspase3-mediated cell death was observed in Wnt7a-deficient embryos, thereby suggesting possible redundancy between Wnt1 and Wnt7a in this context.
Wnt7a has differential roles in DA axonal growth and pathfinding in development
We, and others, recently identified Wnt5a (via the activation of the Wnt/PCP pathway) as an integral protein in the development of the midbrain DA pathways [6,24]. Here, we demonstrate a temporal and spatial expression of Wnt7a within the developing brain also suggestive of roles in DA axon morphogenesis. Our findings of early Wnt7a expression within the VM, combined with its roles in axonal elongation and chemorepulsion at E13.5 (Fig. 5), were suggestive of a role in initiation of neurite extension and repulsion out of the VM toward the forebrain targets, similar to the effects previously observed for Wnt5a [6,24]. Later, we maintained Wnt7a expression within the ventricular zone of the VM, in addition to strong midline expression in the intermediate and marginal zones (Fig. 1H). In light of the maintained chemotaxic role of Wnt7a at this age, we speculate that Wnt7a's expression in the VM at this time may be to prevent midline crossing of DA axons. This idea is further supported by the observation that Nkx2.1 mutant mice display aberrant midline crossing of MFB fibers and a disorganization of the caudal hypothalamus suggesting that medial brain structures (and the molecules they produce) are involved in maintaining the (ipsi)-lateral position of the MFB [5,40].
Subsequent to the initial dorsal and subsequent rostral orientation of TH+ neurites, axons adopt a ventrolateral position in the midbrain and extend axons toward the forebrain forming the median forebrain bundles. It has long been speculated that molecular cues emanating from regions flanking the bundles, such as the thalamus and hypothalamus, control this ventrolateral trajectory [4,5]. In the present study, we detected Wnt7a mRNA within the thalamus and hypothalamus (Fig. 1K). Previous studies have shown that while thalamic explants have no long-range chemotropic effects on midbrain DA axons, DA axons fail to enter thalamic explants suggesting the presence of a contact-dependent inhibitor(s) of mdDA axon outgrowth in the thalamus [41]. While Wnt7a is a secreted protein, its levels within these nuclei suggest a local autocrine chemorepulsive cue to prevent DA axon entry. In support, despite the ability to maintain striatal targeting in Wnt7a-deficient embryos (likely the consequence of compensation by other Wnt signaling mediators), we showed defasiculation of the DA MFB in these embryos. Collectively, our findings suggest that the broadening of the bundle in the Wnt7a−/− embryo is a consequence of loss of repulsive cues from the neighbouring nuclei adjacent to the bundle. Finally, we show an inverse expression of Wnt7a expression and TH (marking DA axons and terminals) within the striatum. Here, Wnt7a may additionally prevent premature entry of DA axons into the dorsal before ventral striatum but may additionally play a function in establishment of synapse formation, a role already attributed to Wnt7a elsewhere in the CNS [42 –44]. While we, and others, have previously demonstrated the role of the noncanonical PCP/Wnt pathway in regulating DA axon growth and guidance, here we demonstrate for the first time the role of β-catenin/canonical Wnt signaling in mediating axon growth effects.
While here we identify multiple roles for Wnt7a in dictating the size of the VM neuronal population and influencing the connectivity of the midbrain DA pathways, functions attributed in part to the canonical Wnt pathway, further studies are required to determine which Wnt-related receptors mediate these effects and whether the noncanonical pathways are also activated in these functions. An outstanding question also remaining is the specificity of each Wnt's role in VM development and more specifically VM DA neurons. To what extent is there redundancy between Wnt7a and other established Wnts in VM proliferation (ie, Wnt1 and 2), differentiation (Wnt5a), and axon guidance (Wnt5a/7b)? Future studies employing double and/or conditional knockouts are likely to shed more light on this redundancy as well as the signaling pathways and receptors involved. Together these findings shed more light on the intricacies involved in the establishment of midbrain DA neural networks and may in the future aid in the development of novel therapies for DA-related disorders.
Footnotes
Acknowledgments
This work was supported by grants from the Australian National Health and Medical Research Council (NHMRC) and Victorian-California Stem Cell Alliance grant. B.D.B. was supported by an Australian Postgraduate Award. C.R.B. is an NHMRC Peter Doherty Fellowship. C.L.P. was supported by an NHMRC Career Development Award and subsequent Viertel senior medical research fellowship.
Author Disclosure Statement
The authors declare no commercial associations that might create conflict of interest in connection with the submitted manuscript.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.